普通烟草CCCH 类锌指蛋白家族的全基因组鉴定和表达分析
2022-07-06唐春闺邓兆龙金维环向世鹏谢鹏飞郭红祥刘卫群
唐春闺,邓兆龙,刘 琼,金维环,向世鹏,谢鹏飞,郭红祥,刘卫群
(1. 湖南省烟草公司长沙市公司,湖南 长沙 410011;2. 河南农业大学生命科学学院,河南 郑州 450002)
植物中,CCCH 类锌指蛋白不仅参与植物的生长发育过程[1‐2],同时也参与植物对生物和非生物胁迫的响应[3]。植物CCCH 类锌指蛋白通常包含1~6个保守CCCH 基序[4],CCCH 基序最初被定义为CX6~14-C-X4~5-C-X3~4-H(X 代表任意氨基酸)[5],但后续在拟南芥和水稻中发现了不同于原始分类的CCCH 基序[6],因此,CCCH 基序被定义为C-X4~15-CX4~6-C-X3~4-H[6]。不同植物包含有相同的CCCH 基序[7],并且C-X7-8-C-X5-C-X3-H 为最常见的基序[8]。TZF(Tandem CCCH zinc‐finger)蛋白通常包含有串联重复的CCCH 基序[2],到目前为止,CCCH 蛋白功能研究主要集中在TZF 类蛋白上,非TZF 类CCCH类锌指蛋白研究较少[2‐3]。
CCCH 类锌指蛋白在脱落酸(ABA)、赤霉素(GA)等激素介导的信号通路中发挥作用[9‐10]。拟南芥中,SOMNUS 蛋白通过调节内源ABA 和GA 含量从而调控ABA 和GA 的代谢[11];AtTZF1、AtTZF4、AtTZF5 和AtTZF6 通过ABA 和GA 信号途径,调节拟南芥对高盐、冷和干旱胁迫的耐受性[12‐13]。OsC3H47 通过降低对ABA 的敏感性从而增加水稻对干旱胁迫的耐受性[14]。棉花GhTZF1、水稻OsTZF1 和OsTZF2 、拟南芥AtTZF2 和AtTZF3 都通过茉莉酸(JA)信号途径调节植物叶片衰老[9,15‐17]。除了激素信号途径之外,CCCH 锌指蛋白还参与其他多种生物和非生物胁迫[3,18]。尽管在拟南芥、水稻等物种中发现CCCH锌指蛋白家族基因参与多种胁迫过程,但烟草CCCH 类锌指蛋白基因家族成员的鉴定、环境信号感知以及激素信号响应研究鲜有报道。鉴于此,运用生物信息学方法,以拟南芥CCCH类锌指蛋白家族成员蛋白质基序为基础,在烟草基因组数据库中进行烟草CCCH 类锌指蛋白家族鉴定,并从系统进化、染色体分布、蛋白质理化性质、Motif 预测等方面分析烟草CCCH 类锌指蛋白家族成员的特性;同时,分析家族成员基因NtC3H-39启动子顺式作用元件、基因表达模式以及对干旱、激素的响应,为进一步探究CCCH 类锌指蛋白基因在烟草应对非生物胁迫中发挥的功能奠定理论基础。
1 材料和方法
1.1 CCCH类锌指蛋白基因家族鉴定及系统进化树构建
从茄科数据库(https://solgenomics.net/)下载普通烟草(Nicotiana tabacum)基因组数据(2017 年发布)并建立本地数据库。利用HMMER 3.0软件构建隐马可夫模型,通过拟南芥CCCH 类锌指蛋白基因家族pfam(PF00642)进行本地比对,经过在线网站CD-seach(https://www. ncbi. nlm. nih. gov/Structure/bwrpsb/bwrpsb.cgi)筛选得到烟草中含CCCH 的序列。利用MEGA 7.0 软件,采用最大似然法(Maximum likelihood,ML)构建系统进化树(泊松模型,Bootstrap 检验重复1 000 次),对烟草CCCH 类锌指蛋白基因家族进行亚族分类。
1.2 CCCH类锌指蛋白基因在染色体上的位置、亚细胞定位以及启动子分析
利用TBtools[19]提取基因组注释文件中的基因定位信息,并利用其染色体定位可视化功能,展示烟草CCCH 类锌指蛋白基因序列在染色体上的位置。在烟草数据库中找出Nitab4.5_0004188g0030.1和Nitab4.5_0005411g0020.1(NtC3H-39)基因启动子上游2 000 bp 序列,利用PlantCARE 网站分析其顺式作用元件,并利用TBtools工具进行可视化处理。
1.3 材料的种植、处理和取样
将野生型烟草K326 种子播种在育苗盘中,置于光照培养箱(26~28 ℃,相对湿度70%,光照16 h/黑暗8 h)中培养,至烟苗四叶期时移入花盆(直径15 cm,高22 cm)中。在不同生长时期,分别取根、叶组织,液氮速冻后放于-80 ℃冰箱中保存,用于后续分析。
选取长势一致的野生型K326 烟株进行如下处理:对照组(喷施清水)、生长素组(喷洒20 mL 0.5 mmol/L 生长素溶液)、茉莉酸甲酯(MeJA)组(喷洒20 mL 4.0 mmol/L MeJA 溶液)、乙烯利组(喷洒20 mL 4.0 mmol/L 乙烯利溶液)、脱落酸组(喷洒20 mL 4.0 mmol/L ABA 溶液),24 h 后取顶端第2片叶,液氮速冻后保存于-80 ℃冰箱,用于测定NtC3H-39基因表达量。
干旱胁迫:选取长势一致的6 株烟草,3 株正常浇水,另外3株停止供水,采取自然干燥法进行干燥处理。7 d 后分别取样,液氮速冻后保存于-80 ℃冰箱,用于测定NtC3H-39基因表达量。
1.4 烟草NtC3H-39基因表达分析
采用TRIzol 法提取烟草总RNA,反转录成cDNA,以烟草β-Actin(GenBank:AF126810)作为内参基因,使用实时荧光定量PCR 仪(BioRad)检测NtC3H-39基因表达情况,每个样品设置3 个重复,采用2–ΔΔCt法进行基因表达水平分析。
2 结果与分析
2.1 烟草CCCH类锌指蛋白家族的鉴定
通过本地数据库比对,从烟草数据库中共获得213 条包含CCCH 基序的蛋白质序列。通过进一步结构域筛选,保留E 值<0.01 且包含CCCH 基序的全长蛋白质序列,最终得到86 个蛋白质。分析发现,86 个CCCH 类锌指蛋白中,氨基酸残基长度不一,多数集中在300~800 个(表1)。目前已鉴定的最短的烟草CCCH 类锌指蛋白基因序列号为Nitab4.5_0002783g0030.1,包含148 个氨基酸残基;最长的烟草CCCH 类锌指蛋白基因序列号为Nitab4.5_0000753g0060.1,包含1 615 个氨基酸残基。大多数CCCH类锌指蛋白定位在细胞核中,少部分定位在叶绿体、细胞骨架、细胞质膜、内质网和线粒体上。通过计算CCCH类锌指蛋白的分子质量和等电点发现,烟草CCCH 类锌指蛋白分子质量变化较大,分布在16 662.65~176 551.48 u,等电点在4.51~9.43(表1)。

表1 烟草中CCCH类锌指蛋白基因家族成员Tab.1 The CCCH type zinc-finger proteins in Nicotiana tabacum

续表1 烟草中CCCH类锌指蛋白基因家族成员Tab.1(Continued) The CCCH type zinc-finger proteins in Nicotiana tabacum
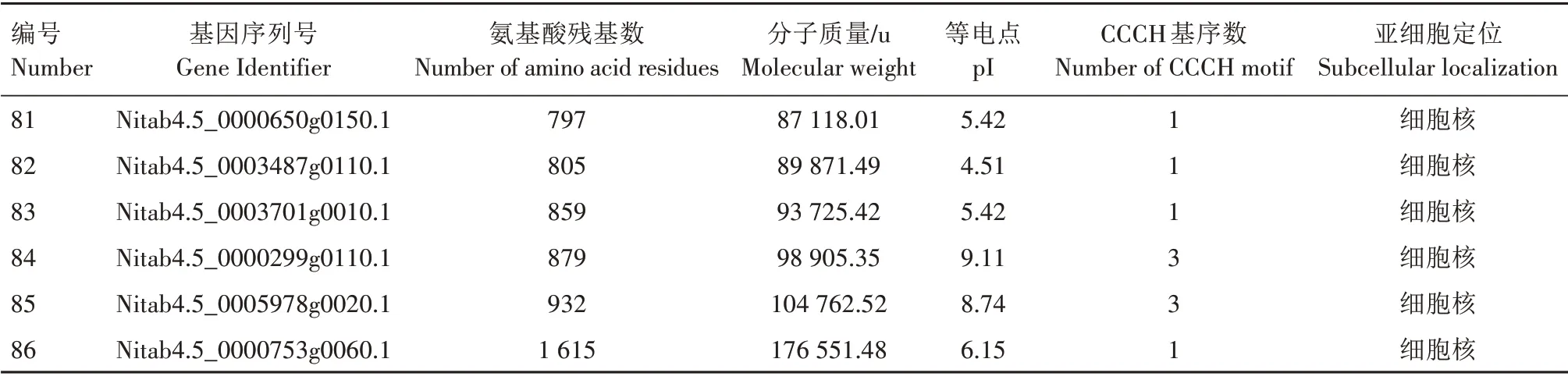
续表1 烟草中CCCH类锌指蛋白基因家族成员Tab.1(Continued) The CCCH type zinc-finger proteins in Nicotiana tabacum
2.2 烟草CCCH类锌指蛋白分类分析
为了研究CCCH 类锌指蛋白家族特性,采用最大似然法将比对出来的86个蛋白质进行聚类分析。参照拟南芥分类方法,将86 个CCCH 类锌指蛋白家族中的51 个蛋白质聚类到9 个亚家族中(Bootstrap值>50%),而位于进化树其他分支的35 个锌指蛋白Bootstrap 值<50%,认为没有聚到一个亚家族中。在这些亚家族中,Ⅰ类亚家族最大,包含11个蛋白质;其次为Ⅹ和Ⅺ亚家族,包含9个蛋白质;最小的家族为Ⅲ亚家族,包含1 个蛋白质(图1)。进一步分析发现,在Ⅰ类亚家族蛋白质中,大部分包含5 个CCCH 基序和6个CCCH 基序,而在Ⅸ、Ⅹ、Ⅺ类亚家族中的蛋白质包含2 个CCCH 基序,在Ⅵ、Ⅶ类亚家族中的蛋白质包含3 个CCCH 基序,Ⅲ类亚家族只有1个蛋白质,其包含1个CCCH基序。

图1 烟草与拟南芥CCCH类锌指蛋白的系统进化树分析Fig.1 Phylogenetic tree of CCCH type zinc-finger proteins in Nicotiana tabacum and Arabidopsis thaliana
2.3 烟草CCCH类锌指蛋白结构分析
通过对烟草CCCH类锌指蛋白家族成员包含的基序数目进行分析,发现烟草中该类蛋白质包含的基序为1~6 个不等。按照基序拷贝数目不同,可将烟草CCCH类锌指蛋白分为6类,其中含有2个基序数目的蛋白质最多,共包括29 个蛋白质,占蛋白质总数的33.7%;其次为包含1 个基序的蛋白质,共25个,占蛋白质总数的29.1%;包含6 个基序的蛋白质共2个(图2)。

图2 烟草CCCH类基序Motif锌指蛋白的统计特征以及其部分基序的序列标识Fig.2 Characterization of CCCH type zinc-finger proteins and sequence logos for common CCCH zinc finger motifs
根据半胱氨酸和组氨酸之间间隔的氨基酸数目不同,可以对基序进一步分类。烟草CCCH 类锌指蛋白共包含11 类基序(图2)。与拟南芥和水稻CCCH 类锌指蛋白一样,烟草CCCH 类锌指蛋白家族结构域高度保守。其中,含有CCCH 基序最多的为C-X8-C-X5-C-X3-H,占比57.71%,其次为C-X7-C-X5-C-X3-H,占比20.57%;基序含量第三的为CX5-C-X4-C-X3-H,占比为8.00%,而C-X8-C-X4-CX3-H、C-X5-C-X10-C-X3-H、C-X9-C-X5-C-X3-H、C-X6-C-X4-C-X3-H 以及C-X10-C-X5-C-X3-H 各有2个,占比均为1.14%。
使用在线工具MEME 与TBtools 对烟草86 个CCCH 类锌指蛋白家族成员蛋白质序列进行可视化分析(图3),结果表明,烟草CCCH 锌指蛋白共包含10 个保守的基序,将其命名为Motif1—Motif10。不同Ccch类锌指蛋白家族成员包含的基序数目不同,Ⅰ类亚家族主要包含Motif9 和Motif5,Ⅵ包含Motif1、Motif4 和Motif8,Ⅹ类亚家族包含Motif1、Motif2、Motif3 和Motif7,说明不同亚家族基序为高度保守结构。结构域分析发现,CCCH 类锌指蛋白还携带其他几种已知的功能性结构域,包括KH、RRM 和zf-CCCH 等(图3)。值得一提的是,这些结构在拟南芥、玉米和水稻中也有发现[6,20]。可见,烟草中CCCH 类锌指蛋白功能可能与拟南芥、玉米和水稻中CCCH类锌指蛋白功能相似。

图3 烟草CCCH类锌指蛋白的结构Fig.3 Schematic structures of tobacco CCCH type zinc-finger proteins
2.4 烟草CCCH类锌指蛋白基因的染色体定位分析
利用Tbtools 染色体定位可视化功能,展示序列在染色体上的位置。在这86 个基因中,只有37个基因定位在17 条染色体上(图4),在烟草的24条染色体中,最长的染色体为17 号染色体,长度为220 Mb,上面定位1 个CCCH 类锌指蛋白基因,第2长的染色体为22 号染色体,上面定位4 个CCCH 类锌指蛋白基因。19 号染色体上定位的CCCH 类锌指蛋白基因最多,达到6 个,最短的染色体为21 号染色体,上面定位到了3 个CCCH 类锌指蛋白基因,其中1号、3号、10号、11号、16号、18号以及20号染色体没有定位到CCCH类锌指蛋白基因。
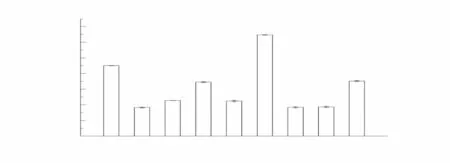
图4 烟草CCCH类锌指蛋白基因在染色体上的位置分析Fig.4 Chromosomal location of CCCH type zinc-finger proteins in Nicotiana tabacum
2.5 烟草NtC3H-39蛋白的序列分析
前期转录组测序发现,烟草基因NtC3H-39(Nitab4.5_0004188g0030.1)在机械损伤后表达量上升(未发表)。为了进一步研究该基因编码蛋白质的功能,以NtC3H-39 蛋白序列构建不同物种的进化树,同时对该蛋白质的结构、理化特性、亚细胞定位以及启动子序列进行分析。
通过分析发现,NtC3H-39 和拟南芥AtC3H39、水稻OsC3H56、玉米ZmC3H56 同源性很高,属于一个亚家族(图5)。该亚家族具备C-X8-C-X5-C-X3-H 基序结构,并且有3 个重复基序(图5)。对NtC3H-39 序列进行比对分析发现,该蛋白质包含331个氨基酸。ExPaSy网站预测其编码蛋白质的分子质量为37.7 ku,等电点为8.02;亚细胞定位(CELLO v.2.5)预测表明,该蛋白质定位在细胞核中。

图5 烟草NtC3H-39蛋白的系统进化关系及序列分析Fig.5 Phylogenetic relationships and sequence analysis of NtC3H-39
通过对NtC3H-39启动子进行分析发现,烟草NtC3H-39启动子包含有茉莉酸响应元件、生长素响应元件、防御和胁迫响应元件以及水杨酸响应元件(图6)。NtC3H-39-1(Nitab4.5_0004188g0030.1)与NtC3H-39-2(Nitab4.5_0005411g0020.1)蛋白质序列相似度高达94.86%,并且二者的启动子包含的响应元件也非常类似,表明二者在烟草中的生物学功能可能相似。

图6 烟草NtC3H-39基因启动子区域中的顺式作用元件Fig.6 Cis-acting elements in the promoter regions of NtC3H-39
2.6 烟草NtC3H-39基因对胁迫的响应
为了进一步分析烟草CCCH 亚家族NtC3H-39基因的功能,检测了该基因在不同生长时期和不同处理下的表达情况。结果如图7 所示,该基因在烟草不同生长时期都有表达,旺长期根和叶中表达量低于幼苗期和成熟期,成熟期根和叶中的表达量最高,表明NtC3H-39基因参与烟草的发育过程。
干旱、生长素、茉莉酸甲酯能显著抑制NtC3H-39基因在叶中表达,而脱落酸和乙烯利则显著诱导该基因在叶中表达;干旱、生长素、茉莉酸甲酯以及乙烯利能显著诱导该基因在根中表达,而脱落酸抑制该基因在根中表达(图7)。表明NtC3H-39基因参与植物对干旱和激素的信号转导,同时,该基因在不同部位对干旱、激素信号响应不一致。


图7 NtC3H-39在不同生长时期以及干旱、生长素、MeJA、脱落酸、乙烯利胁迫下的表达Fig.7 Expression of NtC3H-39 in different growth periods and abiotic stress of drought,auxin,MeJA,ABA and ethrel
3 结论与讨论
CCCH 类锌指蛋白是参与植物生长发育以及响应生物和非生物胁迫的重要调节因子[2‐3]。目前,在多种植物中已经完成了对CCCH类锌指蛋白基因家族成员的鉴定,如拟南芥中68 个[6]、水稻中67 个[6]、玉米中68 个[20]、甘蓝型油菜中103 个[7]。本研究运用生物信息学的方法,在烟草中共鉴定到86 个CCCH 锌指蛋白家族成员,表明烟草该家族基因多样化高于拟南芥、水稻、玉米,低于甘蓝型油菜。
86 个烟草CCCH 类锌指蛋白氨基酸残基数目为148~1 615 个,分子质量分布在16 662.65~176 551.48 u。大多数CCCH 类锌指蛋白定位在细胞核中,少部分定位在叶绿体、细胞骨架、细胞质膜、内质网和线粒体上,大多数CCCH类锌指蛋白的等电点在4.51~9.43。对拟南芥和烟草CCCH 类锌指蛋白家族成员构建系统进化树,结果显示,51 个烟草CCCH 类锌指蛋白家族成员被聚类到9 个亚家族中,同一亚族蛋白质保守基序较为相似,表明蛋白质功能在亚族成员之间高度保守。与拟南芥相比,烟草中包含特有的亚家族成员,如Ⅹ亚家族,表明进化中烟草CCCH 类锌指蛋白基因出现了分化。在染色体定位分析中,只有37 个CCCH 类锌指蛋白基因被定位到染色体上,目前烟草基因组注释不全,可能是造成其他CCCH 类锌指蛋白基因无法定位的原因。与水稻、玉米以及拟南芥相比,烟草CCCH 类锌指蛋白基序的种类和数量上大致相同[6,20],表明保守的基序起着至关重要的功能。但烟草也有特有的基序,如C-X6-C-X4-C-X3-H 等,表明不同物种之间的基序也有差异性。
拟南芥中,CCCH 类锌指蛋白参与植物的发育[2]、激素响应[10‐12]和胁迫反应[13‐15]。NtC3H-39 为烟草CCCH 类锌指蛋白家族成员,该蛋白质可能参与对干旱、ABA、MeJA 等胁迫响应[12‐14,16]。实时定量PCR 结果显示,在烟草不同发育阶段和不同部位,NtC3H-39基因表达量发生变化,表明该基因参与烟草发育过程;同时,通过干旱和外源激素胁迫处理后,发现NtC3H-39表达量发生显著变化,表明NtC3H-39参与对干旱、ABA、生长素、MeJA 以及乙烯利的响应,功能上与拟南芥CCCH 类锌指蛋白相似。
本研究利用生物信息学方法对烟草CCCH类锌指蛋白基因家族成员进行鉴定分析,并对家族成员NtC3H-39 蛋白进行序列和表达分析,为后续研究烟草CCCH类锌指蛋白基因功能提供参考。
