黄土高原半干旱区2020年生长季草地土壤呼吸特征及其影响因素
2022-07-01张琳焓闭建荣张雪腾李正鹏赵长明马晓俊
张琳焓,闭建荣,张雪腾,李正鹏,赵长明,马晓俊
(1. 兰州大学大气科学学院,半干旱气候变化教育部重点实验室,甘肃 兰州 730000;2. 西部生态安全省部共建协同创新中心,甘肃 兰州 730000;3. 甘肃省半干旱气候与环境野外科学观测研究站,甘肃 兰州 730000;4. 兰州大学生命科学学院,甘肃 兰州 730000;5. 兰州大学科学观测台站管理中心,甘肃 兰州 730000)
引 言
全球土壤中有机碳储量为1300~2000 PgC,分别是陆地植物碳库(560 PgC)的3 倍和大气碳库(750 PgC)的2 倍,其微小变化将会对大气中二氧化碳浓度产生重大影响,从而加剧或减缓全球气候变化[1-2]。世界干旱半干旱区面积为5.55×107km2,约占全球陆地总面积的41%[3],是全球陆地表面增温最显著的区域,主导着全球碳循环的年际变化[4]。然而,这里降水稀少,蒸发量大,植被覆盖稀疏,生态系统十分脆弱,其土壤呼吸速率的变化对温度和水分的响应更加敏感[5],对全球碳平衡过程影响较大。研究表明,虽然热带雨林是全球陆地生态系统中最主要的碳汇,但是半干旱区由于具有较大的降水变率与气温年际差异,其对全球陆地生态系统碳汇年际变异的贡献约为39%,明显高于热带雨林(19%)和农田(27%)[6]。
目前我国针对土壤呼吸及其影响的研究主要集中在内蒙古羊草草原[7-8]、新疆荒漠植被[9]和青藏高原高寒草甸[10],而对黄土高原半干旱区草地的关注较少。受自然条件制约与人类活动的影响,黄土高原已成为中国水土流失最严重的地区,其生态恢复是国家生态文明建设的主要任务之一[11-12]。在气候条件、植被类型、土壤性质和微生物活性等因素的综合影响下,该地区土壤呼吸常常表现出很大的时空分布差异,极大限制了对碳交换过程的全面认识。李旭东等[13]研究表明黄土高原农田系统土壤呼吸速率的日变化和季节变化均呈明显的“单峰型”曲线,且土壤温度是主控因素,而土壤水分与呼吸作用季节变化的相关性较差;郭丁等[14]使用概率反演分析法对比分析了兰州大学半干旱气候与环境观测站(Semi-Arid Climate and Environment Obser⁃vatory of Lanzhou University,SACOL)草地和农田生态系统,发现黄土高原半干旱地区草地的固碳潜力是农田的1.2 倍;黄土丘陵区围封草地土壤呼吸的日变化曲线接近于正弦函数模型,土壤温度、水分与土壤呼吸的日变化之间呈极显著的指数关系[15];陕西长武黄土旱塬草地土壤温度对土壤呼吸季节变化的作用最大,对其变异的解释程度为79%[16]。上述工作主要探究了土壤温度或土壤水分等单变量因子对呼吸速率的影响,不足以全面刻画环境因子与土壤呼吸的相互作用。此外,受观测仪器和探测技术的制约,以往工作对呼吸速率的研究主要集中在生长季的白天时段,缺乏长期、连续的昼夜变化资料,极大影响了对黄土高原碳循环机制的全面了解。因此,本文分析了黄土高原半干旱区草地系统的土壤呼吸速率、土壤温度及含水量的连续观测数据,试图揭示生长季土壤呼吸的日动态和月变化规律,研究水热因子对土壤呼吸的综合影响,建立合理优化的非线性多元回归方程,以期为准确评估该地区的碳循环过程与影响机制提供基础支撑。
1 研究区概况
试验设置在SACOL 站,位于兰州大学榆中校区萃英山顶(104.137°E,35.946°N,海拔1965.8 m)。气候类型属大陆性半干旱气候,年平均气温6.7 ℃、年蒸发量1340 mm、年日照时数约2600 h、年降水量382 mm(主要集中在7—9 月)。实验场地为天然草地,受人类活动干扰较少,可代表方圆几百公里半干旱草地的平均状况。土壤类型为栗钙土,0~10 cm土壤层的有机碳、全氮、pH值和土壤容重分别为8.96 g·kg-1、0.75 g·kg-1、8.45和1.14 g·cm-3,10~20 cm土壤层的对应值分别为7.42 g·kg-1、0.69 g·kg-1、8.49 和1.14 g·cm-3。主要植被有铁杆蒿(tripolium vulgare)、冷蒿(artemisia frigida)、赖草(leymus secali⁃nus)和本氏针茅(stipa bungeana)等[17]。
2 试验设置
在平坦草地内选择一块20 m×20 m的实验样地,样地内植被分布总体较均匀,但在局地尺度上,有多种类型的植被和裸地分布,为使观测结果更具代表性,采用LI-8100A 型多通道土壤碳通量仪测定土壤呼吸速率时,选取8个不同植被覆盖密度的样点分别放置长期测量气室,相邻气室间隔4 m,按逆时针方向依次标记为1~8号。每个气室内的植被分布状况如图1 所示,其中1、2、7 号植株覆盖相对稀疏,其余气室的植被覆盖度为65%~80%。测量时每个气室都放置于一个直径20 cm、横截面积371.8 cm2、高度10 cm 的PVC 呼吸测定圈上,测定圈嵌入土壤深度为6 cm,为减小安装对测量的干扰,测定圈在测量前2 d嵌入土壤中。

图1 8个气室内的植被分布状况(2020年8月27日拍摄)Fig.1 Vegetation distributions in eight air chambers(taken on August 27,2020)
3 资料与方法
观测时段为2020 年5 月20 日至10 月20 日(生长季),每30 min依次完成8个气室土壤呼吸速率的测量,即每天进行48次连续测定。每个气室还配备一个土壤温度(8100-203 型)和水分(GS1 型)传感器,同步测量5 cm 土壤温度及含水量。降水量和太阳辐射资料分别由SACOL站的雨量筒(TE525MM-L型)和短波辐射表(CM21 型)测量,时间分辨率均为30 min。5 月20 日至7 月20 日3、4、7 号气室的温度传感器被送回厂家进行校准标定,该时段的土壤温度资料缺失;5 月26 日至7 月4 日,由于雨量筒仪器故障,该时段的降水资料缺失。
LI-8100A 型土壤碳通量仪的测量原理是通过测量气室内CO2的增加速率,推算气室外CO2扩散到自由大气中的通量,具体公式①Instruction manual for the LI-8100A Soil Gas Flux System&the LI-8150 Multiplexer.LI-COR Biosciences,Lincoln,Nebraska,USA.如下:
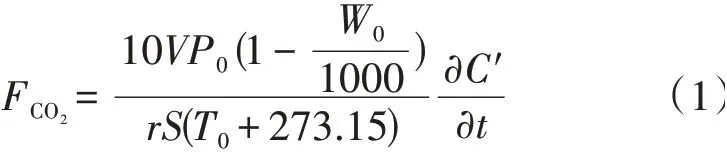
式中:FCO2(µmol·m-2·s-1)为土壤呼吸速率;V(cm3)为气室体积;P0(kPa)为初始压强;W0(mmol·mol-1)为初始水蒸气摩尔分数;r为气体常数,取值8.314 J·mol-1·K-1;S(cm2)是所测土壤环内面积;T(0℃)是初始温度指经水分校正后CO2摩尔分数的初始变化速率①Instruction manual for the LI-8100A Soil Gas Flux System&the LI-8150 Multiplexer.LI-COR Biosciences,Lincoln,Nebraska,USA.。
原始数据按如下步骤处理:首先根据仪器公司提供的File Viewer 软件导出8 个气室的土壤呼吸速率、土壤温度及含水量数据,通过对不同气室测量值的相互比较与验证,有效剔除因仪器故障或外界干扰等产生的异常野点数据,然后利用SPSS 22.0和IDL8.3软件进行相关分析、回归拟合与绘图。
分别用线性[18]和指数[19]模型拟合土壤温度或含水量单一变量对呼吸速率的影响,其中指数模型公式如下:
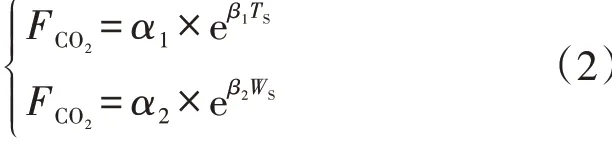
式中:TS(℃)为土壤温度;WS(m3·m-3)为土壤含水量;α1(µmol·m-2·s-1)为温度为0 ℃时的土壤呼吸速率;β1为温度敏感性系数;α2(µmol·m-2·s-1)为土壤含水量为0 m3·m-3时的土壤呼吸速率;β2为水分反应系数。
通常使用温度敏感性指数Q10研究土壤温度与土壤呼吸速率的关系,Q10是指温度每升高10 ℃时土壤呼吸增加的倍数。作为一个重要的模型参数,Q10已被广泛应用于各种生态碳循环模型的模拟中[20],具体公式如下:

考虑到水热因子的共同影响,分别采用线性和非线性模型(交叉、幂次、复合)[21]研究土壤温度与含水量的协同作用对呼吸速率的影响,并根据决定系数R2筛选最优拟合模型。
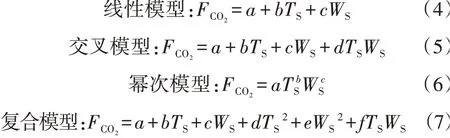
式中:a、b、c、d、e、f均为拟合参数。
4 结果与分析
4.1 土壤呼吸速率及环境因子的时间变化
4.1.1 土壤呼吸速率及环境因子的逐30 min变化
图2为2020 年生长季土壤呼吸速率、土壤含水量、土壤温度和降水量的逐30 min 变化。可以看出,整个观测期内不同气室的土壤呼吸速率、土壤含水量和温度均呈现较为一致的动态变化,变化范围分别为0.20~7.15µmol·m-2·s-1,0.0~0.39 m3·m-3和10~40 ℃。8 个气室的呼吸速率都是从5 月植被返青期开始逐渐增强,7—8 月达到峰值(6.4~7.15µmol·m-2·s-1),9—10月随着植被逐渐变枯黄而减小(2.0~4.0µmol·m-2·s-1)。2020年SACOL站降水主要集中在7—8月,占生长季总降水量(212.4 mm)的74.2%。受降水过程影响,土壤含水量表现出明显的脉冲波动变化,其最大值与降水量具有很好的对应关系,而土壤温度的动态变化主要受太阳辐射和天气状况控制。8 个不同气室的土壤呼吸速率差异较小(标准差为0.10~0.46 µmol·m-2·s-1)且变化趋势较为一致。为减小空间异质性对实验结果的影响,下文使用8个气室土壤呼吸速率、土壤温度及含水量的逐小时算术平均值代表该地区土壤物理参量的典型变化。
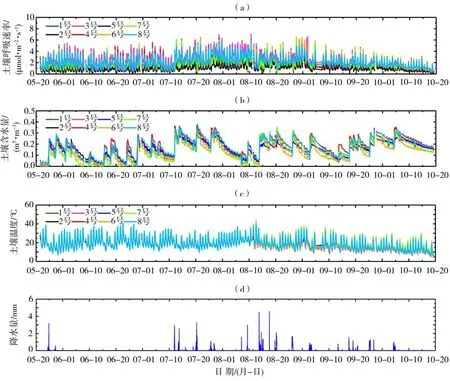
图2 2020年生长季8个气室土壤呼吸速率(a)、土壤含水量(b)、土壤温度(c)及降水量(d)逐30 min变化Fig.2 The 30-min variation of soil respiration rate(a),soil water content(b),soil temperature(c)and precipitation(d)for eight air chambers during the growing season of 2020
4.1.2 土壤呼吸速率及环境因子的日变化
图3为2020 年8 月26 日(晴天)、28 日(多云)和29 日(阴天)3 种典型天气条件下土壤呼吸速率的日变化个例。可以看出,不同天气状况下土壤呼吸速率的日变化差异明显,晴天和多云天均呈“单峰型”曲线,即从日出前开始逐渐增强,正午达最高值,午后至20:00(北京时,下同)为快速减弱阶段,而夜间下降趋于平缓;晴天土壤呼吸速率的日平 均 值 为2.90 µmol·m-2·s-1,变 化 范 围 为1.73~4.92µmol·m-2·s-1,最大和最小值分别出现在13:00和06:00;多云天呼吸速率的日均值(2.60µmol·m-2·s-1)和变化范围(1.86~3.91µmol·m-2·s-1)均低于晴天,最高值出现时间比晴天提前1 h;而阴天呼吸速率一直处于较低水平(1.44~1.89µmol·m-2·s-1),日变化规律不明显。
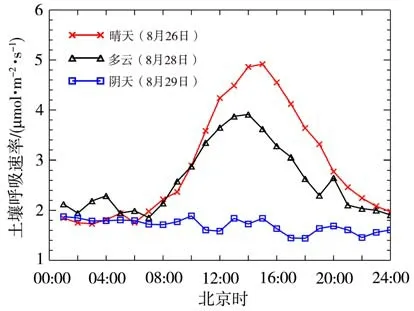
图3 3种典型天气条件下土壤呼吸速率的日变化Fig.3 Diurnal variations of soil respiration rate under three typical weather conditions
图4为生长季不同月份土壤呼吸速率、土壤温度和含水量的平均日变化。可以看出,5—10月土壤呼吸速率的平均日变化都呈明显的“单峰型”曲线,表现为正午较高而早晚时段偏低,与土壤温度的变化特征一致性较好,呼吸速率最低值出现在05:00 或06:00,变化范围为0.71~1.70µmol·m-2·s-1,而最高值出现在12:00 或13:00,对应变化范围为2.20~4.40µmol·m-2·s-1。这主要由于夜间时段土壤温度较低,植被根系活动和土壤微生物活性也相对微弱,而白天土壤温度会随着太阳辐射的增强逐渐升高,适宜的温度促进了土壤微生物的分解和植被的光合作用,使得土壤呼吸在正午达到最大值,即在适宜温度范围内,土壤呼吸随着温度的升高而增强;午后随着土壤温度继续升高至最大值,微生物酶分解的活性受到抑制,导致呼吸作用逐渐减弱,出现“时滞”现象,即土壤温度最大值的出现时间比土壤呼吸滞后1~2 h。各月土壤呼吸速率的总体趋势为8 月>7 月>5 月>9 月>6 月>10 月,日均值分别为2.52、2.51、1.81、1.80、1.62、1.17 µmol·m-2·s-1,均接近于10:00 或19:00 的观测值。而平均土壤含水量的波动变化较小,在00:00—12:00 基本保持恒定,从13:00开始呈逐渐下降趋势。
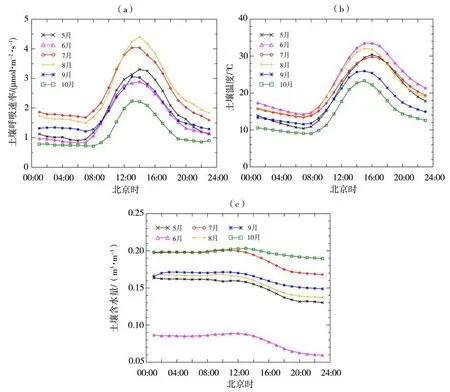
图4 2020年生长季不同月份土壤呼吸速率(a)、土壤温度(b)和含水量(c)的平均日变化Fig.4 Average diurnal changes of soil respiration rate(a),soil temperature(b)and soil water content(c)for different months during the growing season of 2020
4.1.3 土壤呼吸速率及环境因子的月变化
本文将08:00—18:00 规定为白天时段,20:00至次日06:00 定为夜间时段,分别计算生长季逐月白天、夜间及全天土壤呼吸速率、土壤温度及含水量的平均值(图5)。可以看出,白天和夜间土壤呼吸速率的变化趋势较为一致,均在5—6月处于较小值,从6 月开始逐渐增大,8 月达到峰值(白天3.31±0.98 µmol·m-2·s-1、夜间1.80±0.39 µmol·m-2·s-1),之后开始逐渐减小,10 月降到最低值(白天1.55±0.55 µmol·m-2·s-1、夜间0.81±0.12 µmol·m-2·s-1)。这是由于随着气温回暖、降水增多,植被生长良好,适宜的水热条件促进了植被的光合作用、根系呼吸和微生物对土壤有机质的分解,使得呼吸作用明显增强,在植被生长最旺盛的8月达到峰值;而到了生长季末期,呼吸速率随着土壤温度的降低和降水过程的减少而逐渐减弱。另外,整个生长季土壤呼吸速率全天、白天和夜间的平均值分别为1.80±1.01、2.35±1.02、1.39±0.52µmol·m-2·s-1,如果只用白天时段观测的平均值代表全天平均值,将对土壤呼吸造成约30.6%的高估。早期研究[22]指出,夜间草地系统的土壤呼吸作用较弱,对全天的贡献远小于白天,在观测和模拟时常常被忽略。本文计算发现2020 年生长季不同月份夜间的土壤呼吸约占全天总呼吸的27.2%~32.4%,因此,为准确评估半干旱区草地生态系统的碳源汇收支,应该在模型中考虑夜间土壤呼吸的贡献。
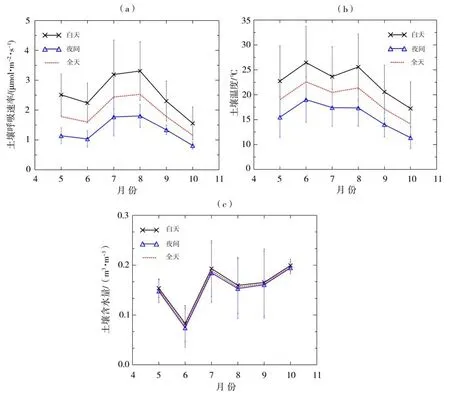
图5 2020年生长季逐月土壤呼吸速率(a)、土壤温度(b)和含水量(c)的白天、夜间及全天平均值(误差棒为对应的标准偏差)Fig.5 The mean values of soil respiration rate(a),soil temperature(b)and soil water content(c)in daytime,nighttime and all-day in each month during the growing season of 2020(Error bars represent the standard deviations)
4.2 环境因子对土壤呼吸速率的影响
土壤呼吸包括植物根系呼吸、土壤微生物呼吸和土壤动物呼吸及含碳物质的化学氧化作用,这些复杂的过程受到土壤温度、土壤水分、降水、有机质含量、植被生长状况和土壤微生物酶活性[23-26]等多种因素共同影响,而土壤温度与含水量通过直接或间接调控上述复杂的生物、非生物过程,对土壤呼吸碳通量交换产生重要影响[7]。
4.2.1 土壤呼吸速率与土壤温度的关系
图6为2020 年生长季土壤呼吸速率与土壤温度的散点图,其中指数和线性模型的拟合方程分别为FCO2=0.7e0.05TS和FCO2=1.16+0.09TS,决定系数R2分别为0.605 和0.612(P<0.001),即指数和线性模型分别可解释土壤呼吸速率变异的60.5%和61.2%,线性模型略优于指数模型。结果表明,土壤呼吸速率随着土壤温度的增加(降低)而呈现明显的升高(减小)趋势,二者存在显著正相关关系,即土壤温度是影响呼吸速率季节变化的重要驱动因子,主要通过直接控制植被根系代谢过程和土壤微生物的分解作用,从而对土壤呼吸产生影响。
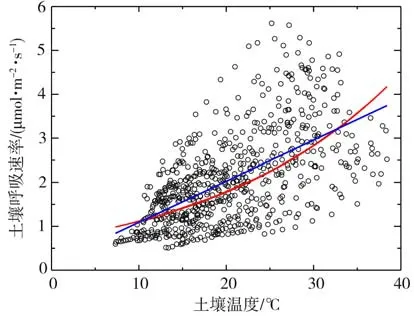
图6 2020年生长季土壤呼吸速率与土壤温度的散点图Fig.6 The scatter diagram between soil respiration rate and soil temperature during the growing season of 2020
4.2.2 土壤呼吸速率与土壤含水量的关系
图7为2020 年生长季土壤呼吸速率与土壤含水量的散点图,其中指数和线性模型的拟合方程分 别 为FCO2=1.35e2.03WS和FCO2=1.41+4.45WS,决 定系数R2分别为0.27 和0.29(P<0.001),对土壤呼吸速率变异的解释程度均小于30.0%。与土壤温度相比,土壤含水量与生长季呼吸速率的相关性较弱。

图7 2020年生长季土壤呼吸速率与土壤含水量的散点图Fig.7 The scatter diagram between soil respiration rate and soil water content during the growing season of 2020
4.2.3 水热因子的协同作用
结合土壤温度与含水量建立的双变量非线性模型能更好地解释土壤呼吸速率的变化[9]。分别使用线性、交叉、幂次、复合4种拟合模型,对生长季土壤呼吸速率与土壤温度和含水量的所有小时平均数据进行多元回归拟合(表1)。可以看出,4种模型的决定系数R2分别为0.663、0.735、0.733、0.740(P<0.001),说明双变量非线性方程的拟合效果明显优于单变量模型。另外,分别计算白天和夜间水热因子对土壤呼吸速率变异的解释程度,发现夜间时段水热因子对土壤呼吸速率变异的解释程度为57.5%(幂次模型),而白天仅为44.3%(交叉模型),均远低于全天的74.0%。因此,综合考虑全天时段水热因子协同作用的非线性模型(交叉、幂次、复合模型)对呼吸速率变异的解释程度更好,改进后双因子模型的最优拟合效果达74.0%。这与刘丽雯等[27]、马涛等[28]、王建国等[29]分别在黄土高原中西部的永寿(75.3%)、兰州(63.6%~80.6%)、神木(69.7%~77.4%)等草地系统的研究结论较接近。

表1 2020年生长季土壤呼吸速率与土壤温度和含水量的多元回归模型Tab.1 Multiple regression model of soil respiration rate with soil temperature and soil water content during the growing season of 2020
从上述4 种模型中选取决定系数较高的交叉和复合模型来验证其适用性。挑选出2020 年生长季所有晴天,将晴天条件下的土壤温度和含水量数据代入上述模型,得到的土壤呼吸模拟值和实测值对比如图8 所示。可以看出,2 种模型的模拟结果与实测值变化趋势较为一致,且数值也较为接近。交叉模型拟合结果的平均值、标准差分别为2.02、0.9 µmol·m-2·s-1,而复合模型的对应值分别为2.20、0.98 µmol·m-2·s-1,均与实测值(2.01±0.105 µmol·m-2·s-1)十分接近,且拟合度都大于87.6%,表明这2种模型均能较好地预估半干旱区生长季草地系统的土壤呼吸速率。另外,在植被返青期复合方程对极值的模拟偏高且误差略微偏大,交叉方程的模拟结果与实测值更吻合;而在植被生长茂盛期,复合方程对呼吸速率的拟合效果均优于交叉方程,后者对极大值的模拟存在一定的低估;到了植被枯黄期,受土壤温度、降水及植物生长状况的共同影响,2种拟合方程的模拟值都较实测值偏高。
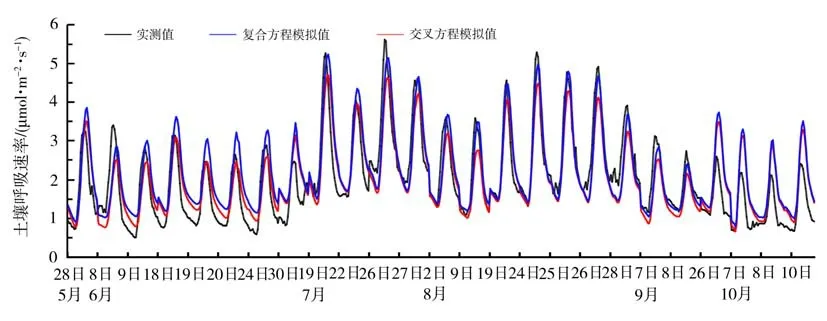
图8 2020年生长季晴天时土壤呼吸速率实测值与模拟值对比Fig.8 Comparison of measured and simulated soil respiration rate on sunny days during the growing season of 2020
5 Q10值变化特征
分别计算2020 年生长季5—10 月全天、白天和夜间的Q10值(表2)。可以看出,全天、白天、夜间的Q10值 变 化 范 围 分 别 为1.38~2.14、1.22~1.96、0.85~1.64,对应平均值分别为1.58±0.23、1.41±0.19、1.20±0.16。不同月份全天Q10值均高于白天和夜间,且除5月外,6—10月白天Q10值也均高于夜间,说明白天时段土壤呼吸速率对土壤温度的敏感性低于全天但高于夜间。通过定量计算发现,如果仅用白天Q10平均值代替总平均值将造成约10.8%的低估。因此,在讨论土壤温度敏感性时,使用全天连续时段观测数据计算的Q10值更准确。由表2还可知,半干旱区草地系统的Q10值从返青期(5 月)开始逐渐降低,到草盛期(6—9 月)下降到最低值后又逐渐升高,在植被枯黄初期(10 月)达到最大值,即草地土壤呼吸的温度敏感性随着植被生长旺盛(枯黄)而减小(增大),这是由于当植物处于不同生长阶段时,昼夜土壤温差和植物根系呼吸的贡献也有所差异,从而使Q10值发生变化[30]。另外,不同月份地上植被凋落物被土壤微生物的分解程度不同,使得土壤有机质也发生了改变[31],这会对土壤微生物的呼吸产生影响,因此未来在研究生长季土壤呼吸速率的温度敏感性时,除了考虑土壤温度和土壤含水量外,还要考虑凋落物的数量与植物的生理指标等因素的影响。LI 等[32]利用LI-6400 型便携式碳通量仪对SACOL 站2008—2010 年3—10 月围封草地土壤呼吸进行了测量,发现围封草地土壤呼吸Q10值的变化范围为1.27~5.76,平均值为1.75,通过对比发现本文在2020 年生长季测得Q10值的全天变化范围及平均值都略低于SACOL 站早期的测量结果,这可能是由测量仪器不同及不同年份之间气候差异造成的,2008—2010 年测量时段土壤的水热条件、呼吸速率、降水量均高于2020 年,使得Q10值也有所升高。此外,LI等[32]仅选择在生长季晴天的上午(08:00—11:00)进行测量而忽略了下午及夜间时段的结果,导致其测量的Q10值偏高。总之,Q10值对土壤水热条件、植被生长状况等因素的变化非常敏感,从而表现出较大的时间差异[33]。
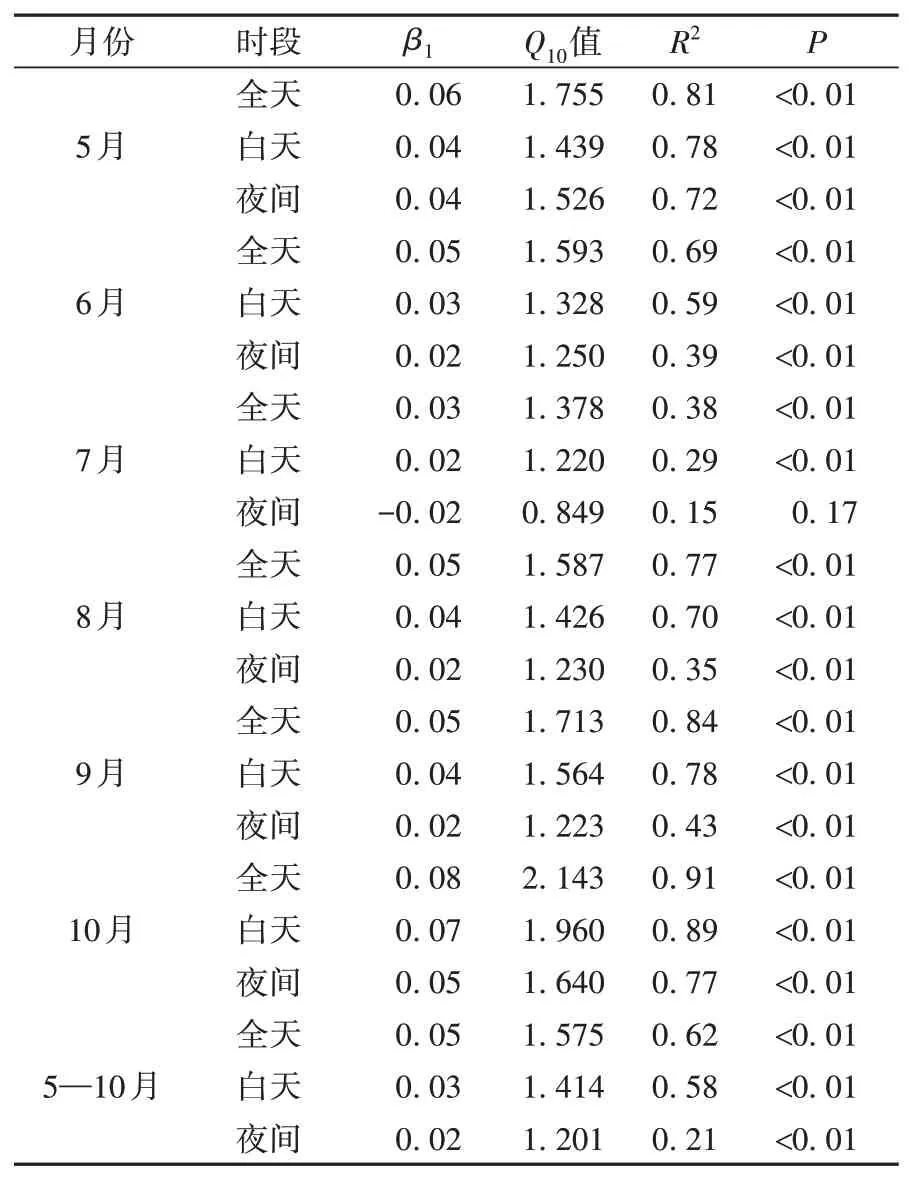
表2 2020年生长季逐月不同时段Q10值Tab.2 The Q10 values in different periods in each month during the growing season of 2020
6 结 论
(1)不同天气条件下土壤呼吸速率存在明显的动态日变化差异,晴天和多云天均呈“单峰型”曲线,而阴天变化不明显,其平均值和变化范围都低于晴天与多云天。
(2)整个生长季不同月份土壤呼吸速率的平均日变化也呈明显的“单峰型”曲线,表现为中午高、早晚低的规律;不同月份的土壤呼吸速率在0.71~4.40µmol·m-2·s-1之间波动,总体趋势为8 月>7 月>5 月>9 月>6 月>10 月。平均日变化的最高值通常出现 在12:00 或13:00,而最低值出现 在05:00 或06:00,日均值接近于10:00或19:00观测值。
(3)白天和夜间土壤呼吸速率在5—6月处于较小值,从6 月开始逐渐增大,8 月达峰值(白天3.31±0.98 µmol·m-2·s-1、夜间1.80±0.39 µmol·m-2·s-1),9—10 月始终呈下降趋势,最低值(白天1.55±0.55 µmol·m-2·s-1、夜 间0.81±0.12 µmol·m-2·s-1)出现在10 月;受太阳辐射和土壤温度等因素影响,白天的呼吸速率总是高于夜间;各月份夜间土壤呼吸对土壤总呼吸的贡献约为27.2%~32.4%,因此,在生态模型的模拟中夜间的土壤呼吸不应被忽略。
(4)土壤温度、土壤含水量等单变量模型对生长季土壤呼吸作用变化的解释程度远低于考虑水热因子协同作用的多元回归模型,改进的交叉和复合模型对土壤呼吸变异的解释程度明显提高,分别为73.5%和74.0%。
(5)2020 年生长季全天、白天和夜间Q10值的变化范围分别为1.38~2.14、1.22~1.96、0.85~1.64,平均值分别为1.58±0.23、1.41±0.19、1.20±0.16,即白天时段的温度敏感性低于全天,若只选择白天时段的Q10值近似代表全天的总平均值,将造成约10.8%的低估。因此,为更加准确地估计半干旱区草地土壤呼吸的温度敏感性,应对全天各时段开展长期连续的综合观测。
