基于机器学习的睡眠剥夺注意力易损性分类研究
2022-07-01王晨毋琳常英娟朱军强杨庆玲李磊磊孙泽恒赵萌萌方鹏朱元强
王晨,毋琳,常英娟,朱军强,杨庆玲,李磊磊,孙泽恒,赵萌萌,方鹏,朱元强
1.空军军医大学第一附属医院放射科,陕西西安 710032;2.空军军医大学军事医学心理学系,陕西西安 710032;3.甘肃省白银市第二人民医院放射科,甘肃白银 730914;4.西安市阎良区人民医院放射科,陕西西安 710089
前言
睡眠剥夺是指由于各种原因导致的睡眠缺失,进而引起一系列生理及心理功能改变的状态[1-2]。以往研究表明,睡眠剥夺经常会增加清醒时的睡眠倾向和神经行为的不稳定性,从而导致认知功能缺陷[2-3]。多项研究已经证实持续性注意力会因睡眠不足而严重受损[4-6]。同时,研究发现睡眠剥夺引起的持续性注意力损伤存在显著的个体差异性,即睡眠剥夺易损性[7]。然而,对于这种个体易损性差异背后的神经影像机制目前尚不清楚。
弥散张量成像(Diffusion Tensor Imaging, DTI)是一种基于水分子扩散原理,定量、非侵入反映大脑白质纤维束完整性的磁共振成像技术。Cui等[8]利用DTI技术通过白质纤维束追踪方法发现,额顶网络内部的纤维束连接与睡眠剥夺后个体注意力受损严重程度显著相关。Zhu 等[3]利用纤维束空间统计学方法发现,睡眠剥夺易损者较耐受者在胼胝体膝部、左侧内囊后肢与右侧前放射冠的FA 值显著降低。同时,基于纤维束空间统计学方法的研究也发现胼胝体膝部与上纵束的各向异性分数(Fractional Anisotropy, FA)值与个体对抗睡眠剥夺的能力之间呈显著相关[9]。这些研究表明睡眠剥夺后个体注意力的受损程度可能与大脑白质纤维完整性有关。在此基础上,本研究采用DTI 技术结合机器学习方法,构建睡眠剥夺易损性分类模型,以期寻找睡眠剥夺后易损性差异的生物影像学标志物,进而为临床筛选睡眠剥夺易损个体提供必要的辅助参考。
1 材料与方法
1.1 研究对象
本试验根据《赫尔辛基宣言》进行,并得到空军军医大学第一附属医院临床试验伦理委员会批准(批件号:KY20183005-1 号)。试验开始前每个参与者均被告知本研究的目的和意义,并签署知情同意书。本试验共招募54 例健康志愿者,男29 例,女25例,年龄(22.46±1.81)岁。所有参与者均来自西安市各大院校的本科生及研究生。纳入标准:①右利手;②18~30 岁;③智力水平正常;④无磁共振扫描禁忌。排除标准:①酒精或药物滥用史;②存在神经系统或精神疾病;③存在严重的器质性疾病;④睡眠障碍者;⑤幽闭恐惧症;⑥极端“清晨型”或者极端“夜晚型”。所有研究对象被告知在磁共振扫描前的1周不能摄取任何含有咖啡因或酒精的刺激性饮料。睡眠日记结果显示研究对象平均入睡时间为22:29±0:17,起床时间为6:27±0:11,夜间持续睡眠时间为7:57±0:21。此外,所有研究对象匹兹堡睡眠质量指数(Pittsburgh Sleep Quality Index,PSQI)测验得分均小于5分,表明睡眠质量良好。
1.2 研究流程
在试验期间每名参与者总共接受正常睡眠后和睡眠剥夺后两次磁共振扫描。磁共振扫描前的一天,试验人员向参与者发放腕表并说明使用方法,用于监测参与者的睡眠情况。为了排除睡眠昼夜节律对试验结果的影响,两次试验磁共振扫描时间都设定在早上8点。同时,为了防止睡眠剥夺后的持续效应干扰,两次磁共振扫描的时间间隔设为1周。正常睡眠后和睡眠剥夺后试验的磁共振扫描顺序均采用伪随机的试验设计,以尽量减少扫描先后顺序对最终试验结果的影响。
睡眠剥夺过程从第一天的早上8:00 点开始至第二天的早上8:00结束,总共持续24 h。在此过程中试验参与者可以正常工作学习,但是不能进行任何短暂或长时的睡眠休息(通过检查参与者腕表记录情况来确认)。每个参与者在8:00 进入试验室进行封闭式管理,此后在试验室可以学习、上网等,但不能进行剧烈活动,亦不能离开实试验室。在此期间,试验室为所有参与者提供同样的餐饮食物,22:00 后不再提供,同时由两名试验人员共同监视避免参与者入睡。在第二天早上8:00睡眠剥夺过程结束后进行磁共振扫描。正常睡眠试验要求参与者经过充足的睡眠后(通过腕表记录情况来确认)进行,每个参与者7:40前抵达实验室,8:00开始磁共振扫描。
1.3 行为学数据采集
本试验利用精神运动警觉性任务(Psychomotor Vigilance Test, PVT)来评估每个参与者正常睡眠或睡眠剥夺后的持续性注意力。PVT 是一种广泛应用于测量被试注意状态、唤醒水平、警觉变化的行为测试任务,该测试主要通过对视标刺激及时响应的检测,评估持续注意能力[10]。PVT 通过计算机化行为研究实验生成系统E-prime 3.0 呈现(http://e-prime3.com/)。在任务态磁共振扫描过程中,首先出现试验预备提示语,之后进入正式的试验视觉刺激,视觉刺激为2~10 s随机时间间隔内屏幕中心出现的红点,参与者被要求在屏幕中心出现红点的瞬间按键回应,以记录其反应时间[10]。反应时间大于500 ms的试次定义为脱漏,将睡眠剥夺后被试与正常睡眠后被试脱漏频次的差值作为衡量持续性注意力受损严重程度的指标。
1.4 磁共振数据采集
本试验使用GE Discovery MR750 3.0T 超导型磁共振扫描仪及8 通道头线圈采集数据。扫描过程中,使用橡胶垫与专用耳塞来固定被试头部和降低噪音。在扫描前,告知被试扫描过程中任务态试验相关的试验流程及注意事项。扫描过程中,要求被试睁眼。扫描完成后,询问被试在扫描过程中是否保持清醒。具体的扫描参数如下,DTI结构像:采用单次激励自旋平面回波成像序列,轴位扫描,扫描角度与前后联合连线平行,TR=10 000 ms,TE=83 ms,翻转角=90°,轴位采集70 层,FOV(Field of View)=240 mm×240 mm,层厚2 mm,连续扫描无间隔,矩阵256×256,体素大小0.9 mm×0.9 mm×2.0 mm,包括64个b值=1 000 s/mm2非线性扩散敏感梯度加权方向及一个b值=0 s/mm2非扩散加权方向。扫描完后获得DICOM格式影像数据,用于获取大脑白质弥散特征。
1.5 磁共振数据处理
DTI 数据预处理:使用磁共振数据处理软件FSL(FMRIB Software Library, https://fsl.fmrib.ox.ac.uk/fsl/fslwiki)、基于图像张量配准工具包DTI-TK(Diffusion Tensor Imaging ToolKit, http://dti-tk.sourceforge.net)以及高级图像配准工具ANTs(Advanced Normalization Tools, https://www.nitrc.org/projects/ants)对DTI数据进行预处理以获得大脑白质弥散特性参数。具体步骤如下:①使用FSL中的FDT(FMRIB's Diffusion Toolbox v2.0)工具包对个体空间中的DTI图像进行头动校正、涡流校正及梯度方向旋转;②使用FSL 中的bet 命令对大脑DTI 图像进行去脑壳操作;③使用FDT 工具包中的DTIFIT 命令计算反映白质弥散特性的各向异性分数(Fractional Anisotropy, FA)、轴向扩散系数(Axial Diffusivity,AD)、径向扩散系数(Radial Diffusivity, RD)以及平均扩散系数(Mean Diffusivity, MD);④使用DTI-TK工具包中的仿射变换及非线性变换算法构建基于特定群体空间的张量模板;⑤使用ANTs 工具包中的基于空间形变配准算法及线性插值算法将大脑白质图谱JHU ICBM-DTI-81 变换到步骤④得到张量模板;⑥利用步骤④张量模板构建过程中产生的变换矩阵将JHU ICBM-DTI-81 图谱变换到个体空间[11];⑦最后利用个体空间中的JHU ICBM-DTI-81 模板提取对应48条纤维束的FA、AD、RD及MD值作为特征值进行机器学习[11]。
1.6 机器学习流程
考虑到本研究样本量较小的情况,本研究采用留一法交叉验证评估分类模型的泛化能力。具体流程如下:①在留一法交叉验证中,选取FA值作为机器学习特征,每次选择1 个受试者作为测试集,其余受试者作为训练集,进行迭代训练测试;②将原始特征数据映射到[0,1]上进行归一化处理;③为避免特征维数过多导致的过拟合,使用LASSO(Least absolute shrinkage and selection operator)回归方法进行特征选择,保留回归模型中非零系数对应的特征;④基于选择的特征使用线性支持向量机分类器进行训练以得到分类模型;⑤根据训练出来的分类模型对测试集进行预测,计算FA 作为特征时的准确率、特异性、敏感性、阳性预测值、阴性预测值、受试者工作特性(Receiver Operating Characteristic, ROC)及曲线下面积(Area Under Curve,AUC);⑥分别选取AD、RD、MD 值作为机器学习特征,重复步骤①~⑤,分别计算AD、RD、MD 所对应的准确率、特异性、敏感性、阳性预测值、阴性预测值、ROC 及AUC;⑦将FA、AD、RD、MD 值进行合并作为新的特征,采取与FA 相同的方法进行模型训练,得到新的分类模型,并计算准确率、特异性、敏感性、阳性预测值、阴性预测值、ROC 及AUC;⑧使用1 000次置换检验来评估最佳分类模型在真实情况下的准确率和AUC是否显著高于随机置换分类标签后得到的准确率和AUC。置换检验显著性被计算为大于真实统计量的随机置换次数除以总次数(比如1 000次)。
1.7 统计学分析
使用IBM SPSS Version 24.0软件进行人口统计学信息统计分析。使用K-S检验确定两组被试的年龄、受教育程度及客观睡眠质量(通过腕表测量)等计量资料是否符合正态分布,对符合正态分布的计量资料使用独立样本t检验,用均数±标准差表示;对不符合正态分布的计量资料进行非参数Mann-WhitneyU检验,以中位数(四分位间距)表示。采用卡方检验比较计数资料组间性别的差异。使用相关分析对分类模型决策值与脱漏频次差值进行相关性检验,符合正态分布使用Pearson相关分析,非正态或分类资料使用Spearman相关分析。P<0.05为差异有统计学意义。
2 结果
2.1 人口统计学信息结果
在磁共振数据采集过程,有5 名被试由于头动过大图像伪影较重被排除,因此最后总共有49 人进入后续分析。根据睡眠剥夺后及正常睡眠后脱漏频次的差值对被试进行分组,频次差值较大的为睡眠剥夺易损组,差值较小的为睡眠剥夺耐受组[3]。两组被试在性别、年龄、受教育年限及客观睡眠质量方面差异均无统计学意义(P>0.05);易损组两种睡眠状态间(正常睡眠vs睡眠剥夺)脱漏频次差值明显大于耐受组,差异有统计学意义(P<0.05)。见表1。

表1 人口统计学信息、斯坦福睡眠量表及客观睡眠测量组间比较Table 1 Comparison of demographic characteristics,Stanford sleepiness scale and objective sleep measures
2.2 机器学习分类结果
为了比较不同特征组合对分类结果的影响,在机器学习过程中分别使用白质单一类型特征及其组合特征构建分类模型,以确定分类性能最佳的模型所对应的特征组合。不同特征组合的分类结果见表2。结果表明:与使用单一类型的特征相比,使用白质组合特征所构建的分类器表现性能最佳。具体见表2。ROC 分析表明组合特征分类器的AUC 明显高于其他单一特征分类器。见图1。置换检验分析表明,真实情况下组合特征分类器的准确率和AUC 显著高于随机置换分类标签后得到的准确率和AUC。具体见图2。
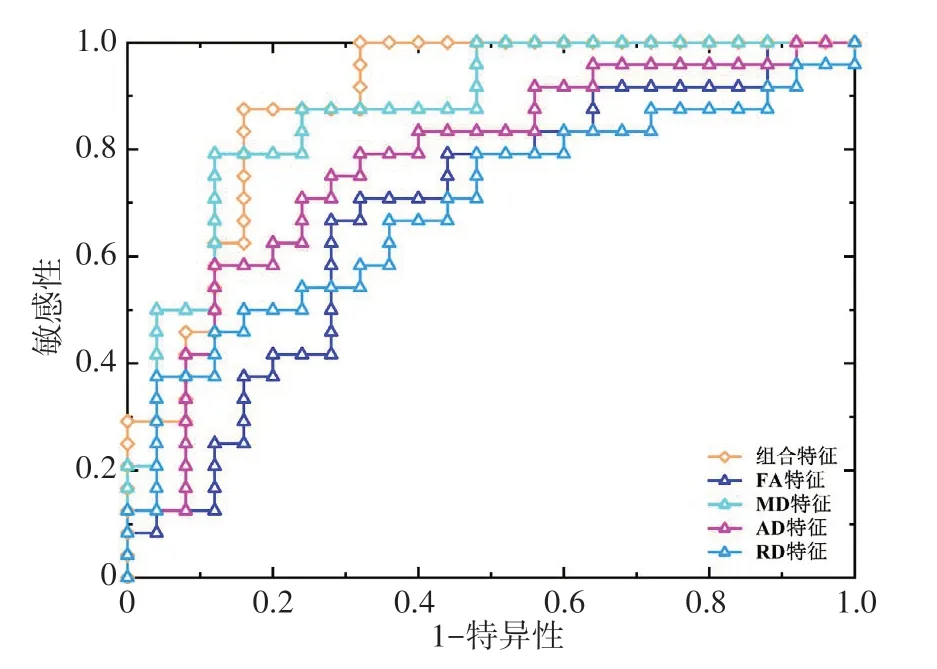
图1 使用不同类型特征构建的分类器ROC曲线结果Figure 1 ROC curve results of classifier constructed with different types of features
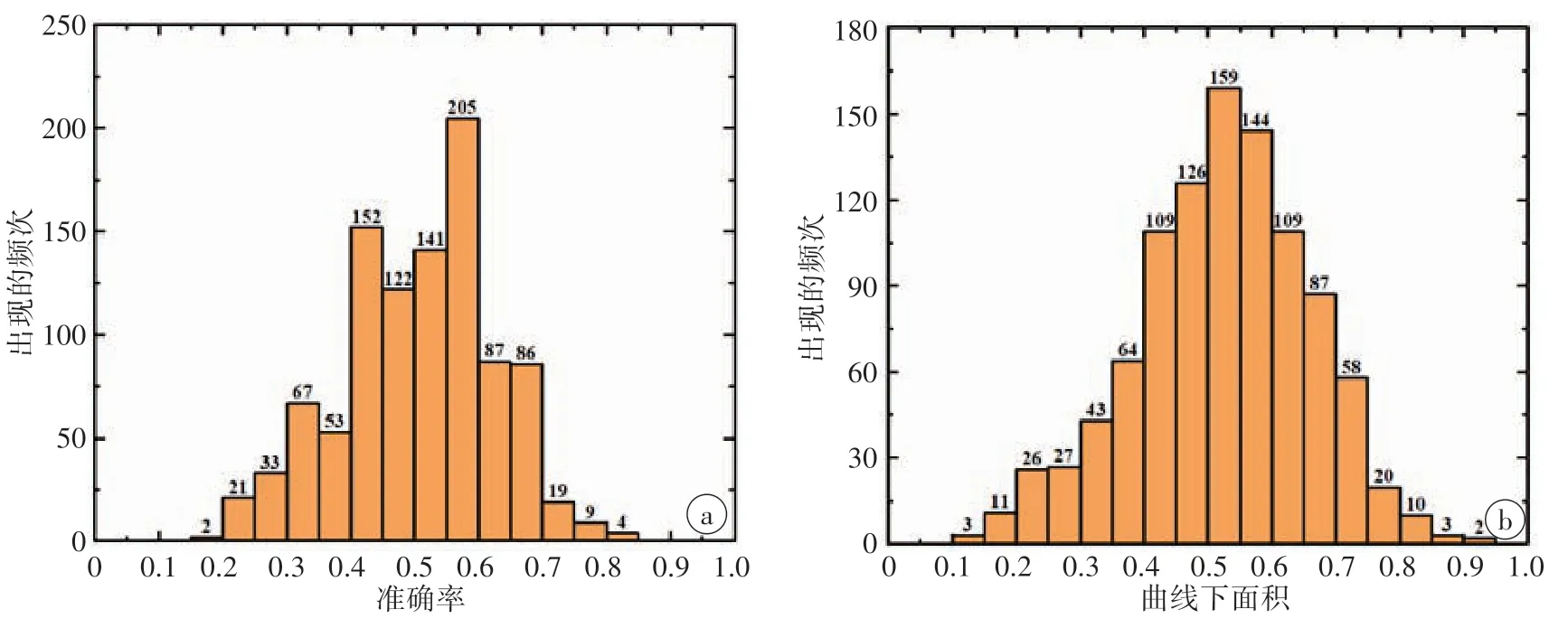
图2 组合特征分类器准确率(a)和曲线下面积(b)的置换检验结果分布直方图Figure 2 Histogram of permutation test results for accuracy and area under the curve of combined features-based classifier
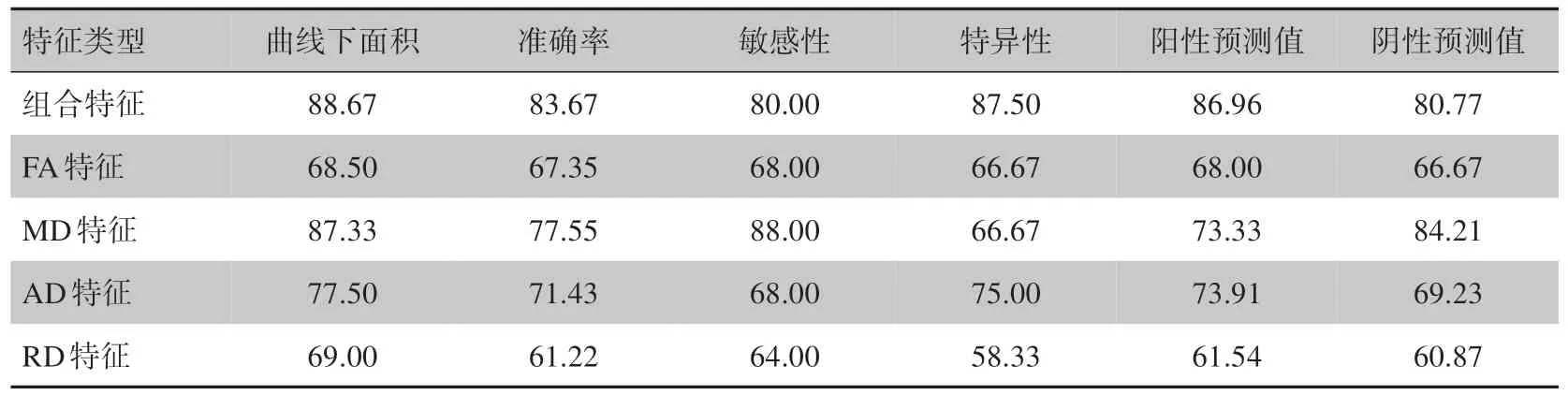
表2 使用不同类型特征的线性支持向量机分类器性能表现(%)Table 2 Classification performances of liner support vector mochine classifiers using different types of features (%)
2.3 分类模型特征权重
由于每次外层留一法交叉验证训练集的样本略有不同,因此每次特征选择过程筛选的特征也不一样。这里将每次外层交叉验证都出现的特征进行报道,特征的最终权重被定义为每次特征权重取和再进行平均。具体特征及最终权重见表3。
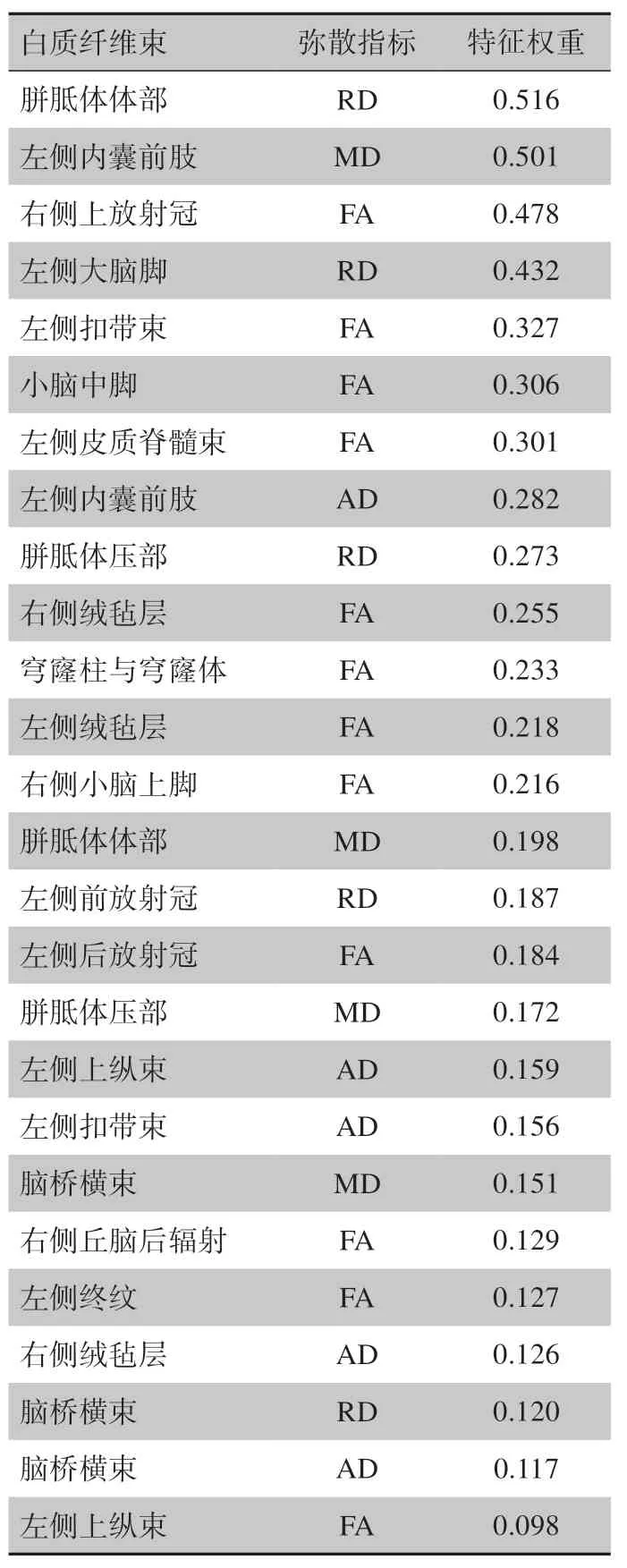
表3 基于留一法交叉验证筛选的图像特征Table 3 Selected discriminative features using the above nested LOOCV strategy
2.4 分类模型决策值与脱漏频次差值的相关性
采用Spearman 相关分析方法考察分类模型决策值与脱漏频次差值的相关性。结果表明,决策值与脱漏频次差值显著负相关(r=-0.549,P<0.01)。见图3。
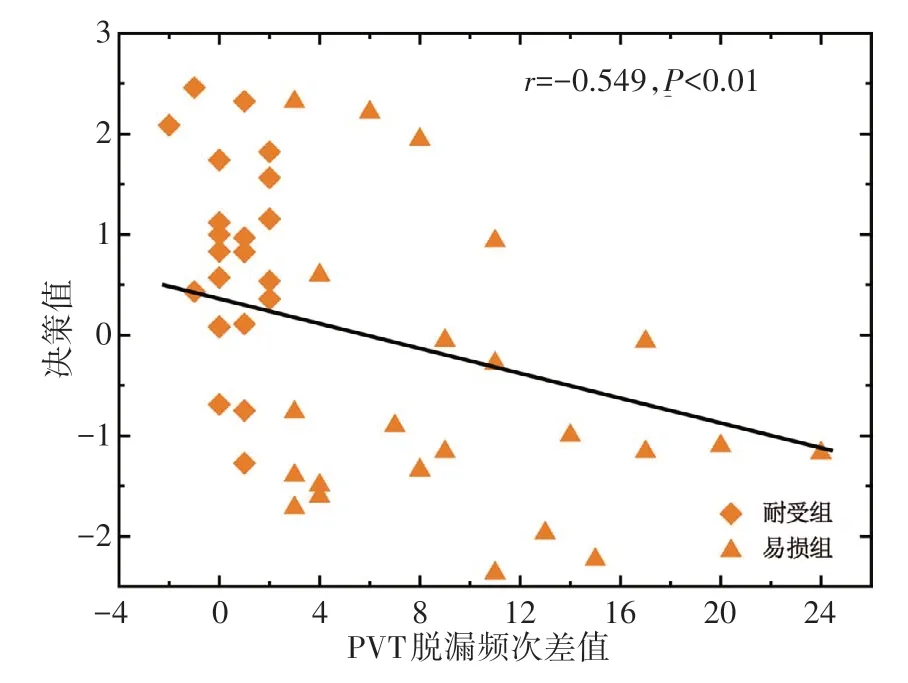
图3 相关性分析散点图Figure 3 Scatter diagram of correlation analysis
3 讨论
本研究基于大脑白质弥散特征采用支持向量机算法探索睡眠剥夺后持续性注意力受损的个体差异性背后的神经影像学机制。研究结果发现,通过不同类型及不同脑区的白质弥散组合特征可以很好地区分睡眠剥夺后持续性注意力易损个体和耐受个体,这表明大脑白质微观结构特性与睡眠剥夺后个体认知不稳定性有关。当前结果发现对分类模型贡献较大的白质纤维束包括放射冠、内囊前肢、丘脑后辐射与皮质脊髓束等投射纤维、上纵束和扣带等联络纤维以及胼胝体和穹窿联合等联合纤维。
神经影像学研究表明睡眠剥夺导致的认知缺失主要由大脑多个脑区间分工合作的协调性破坏所导致,这些脑区包括额顶叶控制区、次级感觉处理区以及丘脑等[12]。同时,脑电图研究也表明持续注意力缺失与包括前扣带回、前额叶区、颞顶叶连接区以及感觉皮层等不同皮层区域的事件相关电位有关[3-4]。这些研究结果表明睡眠剥夺引起的注意力损伤涉及多个皮层区域间的失连接[5,13]。作为连接大脑皮层的主要联合纤维,胼胝体在大脑半球间的沟通和协调中扮演重要角色,其结构损伤可严重影响认知、情感、感觉和运动信息的正常整合[14-16]。最近几项研究也证实了胼胝体完整性和认知不稳定性之间的潜在联系[17-19]。上纵束为连接同侧大脑半球额叶与顶叶的双向神经纤维束,是各脑区间进行信息交互的重要通道。有研究发现,在睡眠剥夺后,睡眠剥夺耐受个体比易损个体在负责高级认知功能的额顶网络表现出更高的激活,这表明额顶网络与睡眠剥夺背景下认知能力的维持有关[20]。一项使用fMRI 激活脑区作为种子点进行白质纤维跟踪的研究发现,PVT缺失的个体差异与上纵束的微观结构特性存在紧密的联系[8]。考虑到上纵束是连接额顶注意网络的主要白质通路[3,21],上纵束结构特性的变异可能是构成睡眠剥夺后个体注意力差异的重要因素。放射冠、内囊前肢与丘脑后辐射中的部分纤维束组成了大脑皮层与丘脑的信号传导通路—丘脑皮质束。丘脑皮质束包括特异性投射系统和非特异性投射系统两个电生理系统,其中非特异性投射系统被认为与睡眠和觉醒调控等意识维持机制有关[10]。因此,丘脑皮质束的结构连接可能决定了睡眠剥夺后个体注意力受损的严重程度。
当前机器学习研究也发现了以往睡眠剥夺注意力研究很少提及的白质区域,比如皮质脊髓束、扣带束及穹窿。造成这种结果差异的原因可能是因为采用的研究方法不同。本研究采用了一种机器学习多变量分析方法—支持向量机,与传统的单变量分析方法相比,它提供了一种更敏感的方法来捕捉区分两组被试的白质纤维束特征。作为边缘系统的核心脑区,扣带回通过扣带束纤维广泛联系额叶、顶叶、颞叶等区域,进而参与多种生理活动和认知活动[22-24]。因此,扣带束的完整性可能对控制行为与情绪状态起着重要作用。穹窿是起自海马内侧的投射纤维,主要沟通颞叶与丘脑之间的联系。有研究发现,与正常睡眠相比,睡眠剥夺后颞叶与丘脑的功能连接显著下降[25],这提示穹窿可能是构成睡眠剥夺后颞叶与丘脑失连接的结构基础。
目前,对睡眠剥夺后个体注意力受损严重程度的评估主要依赖于行为学试验,通常耗时且高度依赖于被试的依从性和试验人员的专业素养。相比之下,基于神经影像的自动判别流程可以有效地避免人工干预和潜在偏差。更重要的是,基于神经影像的机器学习模型可以为睡眠剥夺后注意力损伤的神经基础提供有价值的信息,即“生物影像学标志物”,这是目前尚缺乏但非常需要的[26-27]。在临床实践中,这些“生物标记”信息可以被用来对睡眠剥夺后个体状态进行预判并提前采取相应的干预措施。
本研究还存在几点不足之处:首先,研究样本量较小,缺少独立的测试数据集,进而无法对分类模型的泛化能力进行评估。其次,从数据分析角度出发,特征属性相对单一,当前研究仅仅计算每条纤维束的弥散特性平均值,后续研究可以使用影像组学方法获取更多图像参数,比如:峰度、偏度与标准差等统计量。最后,从模型构建角度出发,特征数量相对较少,当前研究使用的白质图谱仅仅包括主要的48条纤维束,后续研究可以使用相应的纤维追踪算法构建白质网络来进行机器学习研究。
