基于遗传算法优化BP神经网络的石漠化区土壤水分动态预测模型
2022-06-29杨佳琦郭建斌汤明华周金星
杨佳琦,郭建斌,汤明华,周金星,万 龙†
(1.北京林业大学水土保持学院 云南建水荒漠生态系统国家定位观测研究站, 654300,云南建水;2.北京林业大学水土保持国家林业和草原局重点实验室,100083,北京)
岩溶断陷盆地地貌是我国八大岩溶地貌组合类型之一,主要分布于云贵高原,长江、珠江上游地区,因其脆弱而特殊的生态环境和人类不合理的经济社会活动,生态问题日益严重,进而演变成喀斯特岩溶石漠化现象[1]。云南省红河州建水县为典型的岩溶断陷盆地地貌,该地区岩石裸露率高,土壤渗透性强,持水力差,严重制约了生态、经济的可持续发展[2]。植被恢复是岩溶地区生态恢复的关键[3],了解土壤水分动态变化格局,才能对岩溶石漠化进行有效的植被恢复[4]。国内外对于喀斯特岩溶区土壤水分动态变化过程已开展了一些研究[5],但这些研究更多是针对单一石漠化程度下的土壤水分动态变化过程,缺乏对不同石漠化程度和不同植被恢复模式等条件的综合考虑以及较长期的持续性监测,因此预测该区不同石漠化程度土壤水分动态变化对指导区域水土保持,恢复石漠化地区生态具有重要的意义。
土壤水分预测是一个高度非线性的过程,具有复杂性、多变性以及不确定性,预测方法目前主要有模型预测和数据驱动预测2种[6]。BP神经网络模型可以准确度较高地处理复杂的输入输出关系,已逐渐应用于土壤水分预测[7]。目前,众多学者在非喀斯特地区进行了土壤水分预测及其环境响应的研究,如林洁等[8]基于BP神经网络建立太湖典型农田土壤水分预测模型,并利用对气象因子进行敏感性分析来判断影响土壤水分动态的主要气象因子,以主要气象因子建立土壤水分动态模拟模型。丁从慧[9]建立南京典型夏玉米农田的BP神经网络模型模拟土壤水分动态,并讨论了各气象因子对土壤水分的敏感性差异。郝姗姗等[10]在六盘山地区的高建堡、虎沟等11个小流域构建基于BP神经网络的土壤侵蚀因子模型,并分别探究了降雨侵蚀力、坡度坡长等影响因子对土壤侵蚀因子的显著性。李柳阳等[11]构建长三角地区农田站点土壤水分和气象因子的BP神经网络模型预测土壤水分的长时间序列,并对比了基于全部站点和分站点构建的模型精度差异。岩溶断陷盆地季节性干旱严重,土壤水分变化敏感,然而在该区域对土壤水分动态模拟研究薄弱。
大量研究发现,BP神经网络算法的权值和偏置由随机初始化产生,因而模型在训练过程中存在收敛速度慢,易遇到局部最小陷阱的缺点,导致训练效果不佳[12]。遗传算法(genetic algorithms)将网络中所有的权值和偏置组成种群个体,通过计算个体的适应度并选取最优个体,最终将最优个体赋值给网络的初始权值和偏置,提高网络的整体拟合程度[13-14],因此,笔者选取研究区2种程度石漠化样地,分层建立基于BP神经网络的土壤水分动态预测模型,引入遗传算法来优化模型。采用缺省因子法进行敏感性分析,确定影响该区土壤水分动态的主要气象因子,为喀斯特地区生态恢复以及石漠化综合治理提供理论依据。
1 研究区概况
以建水县九标国有林场石漠化治理恢复区为试验研究区域,该区位于云南省红河州建水县(地理坐标E102°54′, N 23°37′),小流域内地貌为典型的石漠化岩溶断陷盆地地貌。海拔230~2 515 m,南亚热带季风气候,年均气温19.8 ℃,年均相对湿度72%,蒸发量2 311.19 mm,年均降水量805 mm,全年无霜期307 d。在2008年,建水县被列入国家首批100个石漠化综合治理试点县之一。试验选取恢复区2种石漠化区样地:中度石漠化区和轻度石漠化区进行研究。试验区于1985年进行了人工植被恢复,其中中度石漠化区样地植被为灌-草结构,主要覆盖植被为车桑子(Dodonaeaviscosa)和硬秆子草(Capillipediumassimile);轻度石漠化区样地为灌-草结构,主要覆盖植被为车桑子和荩草(Arthraxonhispidus)。土壤类型为红色石灰土,2种石漠化样地详细信息见表1,土壤的基本理化性质信息见表2。
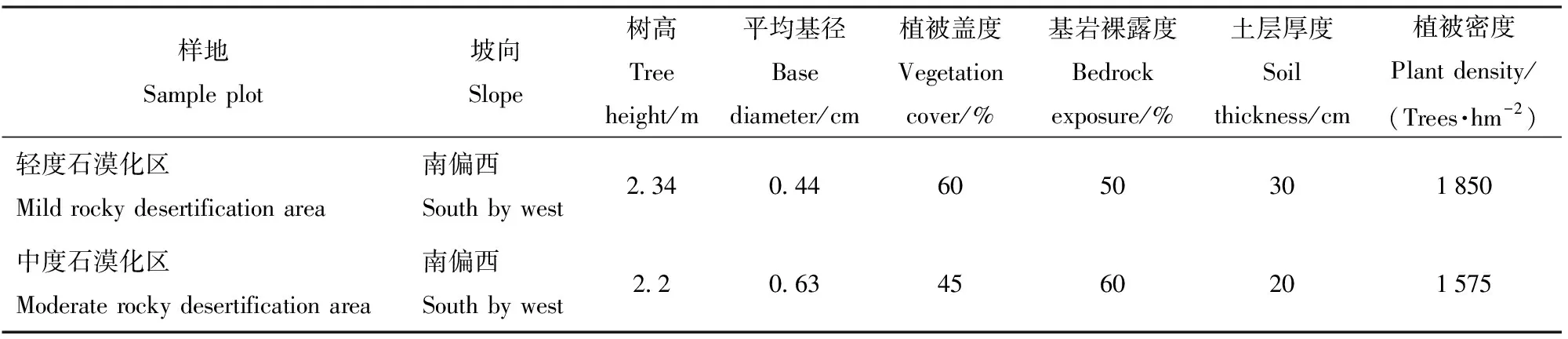
表1 2种石漠化程度样地的基本概况Tab.1 Basic situation of two kinds of Rocky desertification erosion degree

表2 2种石漠化程度样地土壤的基本理化性质Tab.2 Basic physical and chemical properties of soils in two types of rocky desertification plots
2 数据与方法
2.1 数据来源
数据采集于2016年4月16日—2020年12月1日。使用Enviro-SMART CS616土壤水分定位监测系统(Campbell Scientific Inc. USA)每10 min 1次对研究区进行实时监测,测定土壤深度分别为0~10 cm、10~20 cm和20~30 cm。选取同期小型气象站(WatchDog 2900ET, Spectrum Technologies Inc. USA),按照每10 min 1次的频率对研究区气象进行测定。经合并换算得到土壤水分和气象要素日数据,共计1 691组数据。
2.2 BP神经网络模型的建立
2.2.1 BP神经网络模型的建立 笔者采用Matlab软件编程,以6个气象因子作为输入因子,中度石漠化区和轻度石漠化区0~10 cm,10~20 cm,20~30 cm土层土壤含水率分别作为输出因子,共1 691组数据,其中在前1 245组随机抽取800组作为训练数据集,用于训练土壤含水量预测模型,其余445组为验证数据集,用于检验土壤含水量预测模型,最后446组为测试组用于土壤水分动态模拟的性能检验。隐含层的最优节点取值范围根据经验公式求得为[3,12],通过多次网络训练精度,确定最优拓扑结构。轻度石漠化区3土层按照土层深度依次为6∶9∶1、6∶4∶1和6∶3∶1结构;中度石漠化区3土层依次为6∶7∶1、6∶4∶1和6∶7∶1结构。设置最大迭代次数为1 000次; 学习率0.01;输入输出的目标误差0.000 1。在模型训练过程中,将训练数据集输入进行误差的反向传播,不断更新网络权值和偏置,得到最小误差时建立预测模型。在此基础上,引入具有全局搜索能力的遗传算法优化 BP 神经网络。通过6个气象因子逐一缺省进行敏感性分析,得到影响该地区土壤水分动态的主要气象因子。
2.2.2 遗传算法优化BP神经网络模型的建立 遗传算法是由美国人提出的一种模仿生物自然进化过程的随机并行自适应优化方法[15]。具有简单、随机性、鲁棒性和全局性以及不受搜索空间限制性条件约束等特点,可视为介于确定性与随机性优化方法之间的一类新的优化方法[16]。
笔者采用十进制编码,优化过程如下。
1)种群初始化。将神经网络模型所有的权值和偏置建立种群信息,权值与偏置按一定顺序级联起来相当于一个染色体,其长度
M=N(N+O)+Y+O。
(1)
式中:M为染色体长度;N为输入节点数;O为输出节点数;Y为隐含层结点数。
2)个体适应度计算。通过计算找出种群中的每一代进化中的最优染色体并记录保留。笔者选用的适应度函数
(2)
式中:n为样本数;(i)和y(i)分别为样本i的模拟值和实测值。
3)遗传进化。循环进行选择、交叉、变异,个体适应度计算的遗传叠代操作,笔者采用算数交叉,交叉概率为0.4,变异概率为0.08。
4)得到最高个体适应度个体,实现优胜劣汰。最终赋值给BP神经网络模型的权值和偏置,来提高网络的整体拟合程度,使其得到优化。
2.3 模拟效果评价方法
笔者采用平均相对误差(YMARE)、均方根误差(YRMSE)和决定系数(R2)评价土壤水分预模型的精度[17]。
(3)
(4)
(5)

2.4 敏感性评价方法
根据敏感性指数(Yi)来判断神经网络内部输出因子对各个缺省因子的响应强度。敏感性指数的数值越大则土壤水分动态对i气象因子越敏感。
Yi=YRMSE(-i)/YRMSE。
(6)
式中:Yi为第i个因子敏感性指数;YRMSE(-i)为不包括i气象因子的模型均方根误差;YRMSE为全因子模型均方根误差。
3 结果与分析
3.1 BP神经网络模型的检验
将训练所得的轻度石漠化区和中度石漠化区不同土层的BP神经网络模型用于预测2019年9月13日—2020年12月1日的土壤体积含水率,并绘制了土壤含水率实测值和预测值对比图(图1)。显示土壤含水量的动态变化趋势模拟结果与实测值有较好的一致性。

图1 2种石漠化区基于BP神经网络的土壤水分动态模拟与实测值对比Fig.1 Comparison of dynamic simulated and measured values of soil moisture in two rocky desertification erosion areas based on BP neural network
总体来说,中度石漠化样地模拟精度高于轻度石漠化样地。从测试样本模拟结果来看,中度石漠化样地YMARE比轻度石漠化低42%~50%,YRMSE低5%~54%,R2高6%~14%。不同土层模拟结果表明,2个样地均为表层土壤水分0~10 cm模拟精度最高,YMARE<20%,YRMSE<4%,R2>0.704 2。随着土层深度增加,模拟精度降低。在20~30 cm土层土壤水分模拟精度较差,YMARE在1%~25%之间,YRMSE在4%~5%之间,R2>0.547 6。由此可以得出,中度石漠化区土壤水分动态模拟效果优于轻度石漠化区,并且随土层深度增加模拟效果呈现降低趋势的结论。
总体上,轻度石漠化区和中度石漠化区的3个数据集YMARB均<25%,YRMSE均<5%,R2均>0.547 6,BP神经网络模型对该区土壤水分进行预测精度有待于优化。
3.2 遗传算法优化BP神经网络模型的检验
将训练所得的轻度石漠化区和中度石漠化区不同土层经遗传算法优化的BP神经网络模型用于预测2019年9月13日—2020年12月1日的土壤体积含水率,绘制土壤含水率实测值和预测值对比图(图2)。对比图1可知通过遗传算法优化BP神经网络模型后各土层土壤预测值和实际值的拟合度更高,故该地区应用遗传算法优化BP神经网络模型可以更好地进行水分动态模拟。
模型检验效果由表3和表4对比可知: 经遗传算法优化后与优化前的预测模型在该地均呈现相同的规律,即中度石漠化区土壤水分动态模拟效果高于轻度石漠化区,并且随土层深度增加模拟效果呈现降低趋势。
优化前(表3),轻度石漠化区测试样本集的YMARE均<26%,YRMSE均<5%,R2均>0.547 6;中度石漠化区YMARE均<14%,YRMSE均<4%,R2均>0.599 6。经遗传算法优化后(表4),轻度石漠化区测试样本集的YMARE均<13%,YRMSE均<5%,R2均>0.696 3;中度石漠化区的YMARE均<6%,YRMSE均<3%,R2均>0.701 2。经遗传算法优化后,测试样本20~30 cm深层土壤水分模拟精度提升最为明显。轻度石漠化和中度石漠化YMARE分别提升45%和63%,YRMSE分别提升3%和12%,R2分别提升约27%和17%。
由表3和表4可以看到轻度石漠化区和中度石漠化区样地经过遗传算法优化后3个数据集的模拟精度评价指标整体较优化前均有明显提升,且对于测试样本模拟的效果优化后较优化前得到较高程度提升。总体来看,轻度石漠化区和中度石漠化区的3个数据集YMARE均<14%,YRMSE均<4%,R2均>0.70。可以得出,表明该地区应用经遗传算法优化的BP神经网络模型可以更好地进行石漠化区土壤水分模拟,且更适合进行土壤水分动态模拟预报。
3.3 敏感性分析
笔者利用缺省因子法确定影响轻度石漠化区和中度石漠化区不同土层土壤水分预测的主要气象因子。通过对6个输入因子(平均气温、降水量、平均风速、太阳辐射、平均相对湿度及气压)的缺省分别进行神经网络模型的建立、检验。该5因子模型的建立采用与6因子模型相同的训练样本、检验样本与算法,使得 6因子模型和5因子模型具有可比性。

图2 2种石漠化区基于遗传算法优化BP神经网络的土壤水分动态模拟与实测值对比Fig.2 Comparison of dynamic simulated and measured values of soil moisture based on BP neural network optimized by genetic algorithm in two rocky desertification erosion areas

表3 2种石漠化区基于BP神经网络的土壤水分模拟效果统计分析
从表5中可以看出,轻度石漠化区和中度石漠化区样地不同深度土层的土壤水分含量均不同程度的受到6种气象因子的影响,总体上看,2样地每层土壤水分含量均对降雨量的敏感度最高,土壤水分对降雨的敏感性指数在1.317 0~1.783 5之间,其次为平均气温,敏感性指数在0.880 9~1.071 2之间,再次为大气压和太阳辐射。不同石漠化程度和土层深度对气象要素的敏感程度都存在着一定的差异。2种石漠化程度样地均表现出随土层深度增加,模拟效果降低的规律。敏感性分析表明, 轻度石漠化在0~10 cm表层土壤对气象因子响应敏感性最高, 而中度石漠化区样地在10~20 cm中层土壤对气象因子响应敏感性最高。

表4 2种石漠化区基于神经算法优化BP神经网络的土壤水分模拟效果统计分析Tab.4 Statistical analysis of soil moisture simulated effect based on BP neural network optimized by neuralalgorithm in two rocky desertification erosion areas

表5 基于遗传算法优化BP 神经网络模型土壤水分动态的敏感性分析Tab.5 Sensitivity analysis of soil water dynamics based on BP neural network model optimized by genetic algorithm
4 讨论
1) BP神经网络模型已在土壤水分预测普遍应用,但应用于土壤水分动态预测的研究较少。在非喀斯特地区,邓建强[18]在红壤丘陵区建立的BP神经网络的土壤水分动态预测模型,预测精度YRMSE为0.814;郭守平[19]在北方风沙区建立的基于BP神经网络的土壤水分动态预测模型R2为0.982,YMARE均<15%;白冬妹[20]在西北黄土高原区建立NARX递归神经网络模型,预测精度R2>0.92,YRMSE<0.77。对比笔者结果可以看到,在非喀斯特地区利用BP神经网络进行土壤水分动态预测,预测精度普遍高于喀斯特地区。这可能由于喀斯特地区具有岩石裸露率高,土壤瘠薄,持水力差的特点,即使在降水较多的雨季,也只能在每次降雨后维持短时间的水分充足状态有关[21]。笔者研究结果所揭示的基于遗传算法优化的BP神经网络模型能够很好地应用于该地区石漠化区土壤水分模拟,对指导区域水土保持、喀斯特地区生态恢复以及石漠化综合治理具有重要的意义。
近年来,有大量研究从不同角度提升了神经网络的预测精度[22-23],如将灰狼算法、粒子群算法、遗传算法、模拟退火法与BP神经网络相结合,但将多种优化方法进行比较,并应用于土壤水分预测模型的研究较少。在今后的研究中,可以考虑应用其他优化方法改进土壤水分动态预测模型,探究更适合于优化该地区的土壤水分预测模型的算法。
2)在喀斯特地区,何兴潼等[24]在贵州喀斯特地区应用滑动平均法和回归分析法得到降水、气温、地表温度为影响贵州喀斯特地区土壤水分变化的最主要因子。在非喀斯特地区韩湘云等[25]在江西省低丘红壤坡面应用冗余分析发现气温、降水、相对湿度、水汽压是土壤水分季节变化的主控因子;林洁等[26]在江苏太湖典型农田应用缺省因子法进行敏感性分析,得到土壤水分含量对气象因子的敏感性大小依次为降水量、蒸发量、平均气温、平均地表温度、平均风速。基于以上研究,可以得出降雨为影响喀斯特地区土壤水分变化最主要的气象因子,笔者获得相同的结论。而在非喀斯特地区,不同月份的主控因子不同,降雨在雨季对土壤水分变化的影响较大,但在旱季气温为影响土壤水分变化最主要的气象因子。这主要与非喀斯特区地域旱雨季气候特点有关。
笔者仅研究了土壤水分与气象因子的关系,基于气象因子的遗传算法BP神经网络模型可以揭示70%左右的土壤水分变化,其他相关的影响因子对土壤水分的影响有待进一步研究。有研究表明,除气象因子外,植被,土壤初始含水量及理化性质及地形、地貌和土地利用等影响因素对土壤水分预测结果的影响较大[27]。在植被覆盖度低,土壤持水性差的中度石漠化样地土壤水分受气象因子的影响更大,因此笔者基于气象因子与土壤水分建立的遗传优化BP神经网络模型对中度石漠化样地的预测精度高于轻度石漠化样地。可以在今后对土壤水分建立预测模型时更细致地探究不同石漠化程度植被覆盖度,土壤特性和地形等相关因子对模型精度的影响。此外,本研究应用敏感性分析方法确定影响土壤水分动态预测的主要气象因子,也可供其他地区进行参考。
5 结论
1)对云南建水喀斯特岩溶断陷盆地2种不同程度石漠化区的0~10 cm、10~20 cm和20~30 cm土层分别建立基于BP神经网络的土壤水分动态预测模型,并采用遗传算法对模型的初始权值和偏置进行优化,经验证,遗传算法优化后的模拟拟合效果较未优化模型明显提高,克服了传统BP神经网络模型存在的收敛速度慢、易陷入局部极小点的缺点。结果表明中度石漠化样地的土壤水分预测模型精度高于轻度石漠化样地,模拟效果随土层深度增加而降低。基于遗传算法优化的BP神经网络模型能够很好地应用于石漠化区土壤水分模拟,更适合进行土壤水分动态模拟预报。
2)通过遗传算法优化的BP 神经网络对气象要素逐一缺省进行敏感性分析,分析土壤水分与气象要素之间的非线性关系,发现影响该研究区土壤水分动态预测精度的最主要的气象因子为降雨,其次为气温。轻度石漠化表层土壤水分对气象因子的敏感性最高, 中度石漠化区样地中层土壤水分对气象因子敏感性最高。中度石漠化样地对各气象因子的敏感性均高于轻度石漠化样地,且随着土层深度的增加,对气象因子的敏感性呈降低趋势。
