江西武夷山珍稀濒危植物南方铁杉种群动态与空间分布
2022-06-27赵家豪叶钰倩孙晓丹兰文军关庆伟
赵家豪,叶钰倩,孙晓丹,兰文军,方 毅,陈 斌,关庆伟,*
1 南京林业大学生物与环境学院, 南京 210037 2 南京林业大学南京现代林业创新协同中心, 南京 210037 3 自然资源部第一海洋研究所海洋生态研究中心, 青岛 266061 4 江西武夷山国家级自然保护区管理局, 上饶 334515
植物种群动态和空间分布格局是植物种群生态学研究的核心内容之一,主要研究植物种群数量在时间和空间的变化规律、密度的大小、分布与空间结构和种群动态调节机理等[1]。研究植物的种群结构与动态研究可以客观反映植物种群历史动态特征及现状,并预测未来演变发展趋势[2]。目前学者通常对濒危植物的种群数量动态特征和空间分布格局进行分析,能够对濒危种群的现状认知和保护策略探讨提供强有力的科学依据。例如:刘浩栋等[3]通过探讨珍稀濒危植物陆均松(Dacrydiumpectinatum)天然种群的龄级结构及分布格局,为陆均松的保护和恢复提出了合理的适应性策略;任毅华[4]发现了色季拉山急尖长苞冷杉(Abiesgeorgeivar.smithii)不同龄级立木的空间格局分布规律,为该种群保护与实践提供理论依据依据。因此分析珍稀濒危植物的种群动态和空间分布格局很有必要,对珍稀濒危植物的保护和管理具有重要的指导意义[5]。
南方铁杉(Tsugachinensisvar.tchekiangensis)属松科铁杉属,为铁杉(Tsugachinensis(Franch.)Pritz)的变种,南方铁杉分布广,但数量少而分散[6]。江西武夷山国家级自然保护区海拔1500—1900 m范围内的南方铁杉的分布面积达1560 hm2,是这一特有珍稀物种在我国少有的较大天然种群分布区域[7]。目前已有学者报道了关于南方铁杉地种群数量动态[8]、更新能力[9]、种间关系[10]、生态位特征[11]、林隙干扰规律[12]及菌根生态学[13]等方面的研究。关于南方铁杉的种群动态研究最早由杜道林[14]报道,发现贵州茂兰的南方铁杉种群呈衰退状态,更新能力极差。江西武夷山猪母坑地区[15]、广西猫儿山[16]及湖南桃源洞牛石坪地区[17]南方铁杉种群同样存在种群衰退、无法天然更新的现象。张志祥等[18]发现浙江九龙山的南方铁杉种群发育具有波动性,但仍属于稳定型种群。另外,王大来[19]和何建源等[20]利用几项聚集度指标测定发现莽山和福建武夷山的南方铁杉呈随机分布。杨清培等[6]用点格局分析发现南方铁杉幼苗分布随研究尺度增长由聚集分布向随机分布演变。浙江九龙山[21]的南方铁杉在幼苗和幼树阶段呈集群分布,中树和大树向随机分布发展。综上可以发现,由于影响南方铁杉更新和分布的因素较为复杂,不同地区的南方铁杉种群动态特征和空间分布结果具有较大争议。
为进一步诠释南方铁杉种群动态和空间分布特征,本文以江西武夷山国家级自然保护区内180年生的南方铁杉种群为对象,通过年龄结构分析、静态生命表、存活曲线、生存分析和时间序列预测等方法,探讨建群种南方铁杉的种群结构和动态,阐明其天然更新能力。目前点格局分析方法应用于种群的空间分布研究较多,比只能分析单一尺度的传统方法更能揭示种群在连续样地中的扩散过程[4],因此本文采用点格局分析Ripley′sK函数衍生的成对相关函数g(r)明晰各生长阶段的南方铁杉在不同尺度下的空间分布格局[22],综合探讨该地南方铁杉种群的保护策略,以期为珍稀濒危植物南方铁杉的就地保护提供依据。
1 材料与方法
1.1 试验区概况
江西武夷山国家级自然保护区位于江西省上饶市铅山县南沿、武夷山脉北段西北坡,地理位置117°39′30″—117°55′47″N,27°48′11″—28°00′35″E,平均海拔约1200 m,属于海洋性气候和大陆性气候的过渡地带,区内地貌形态为强烈侵蚀的岩浆岩中山地貌,土壤是典型的亚热带中山土壤[23]。年均气温14.2℃;年均降水量1814—3545 mm;年均蒸发量553—941 mm;年均湿度80%—92%。保护区主要为中亚热带中山山地森林生态系统,植物地理区系呈现由温带向亚热带过渡的现象[24];植物种类十分丰富,主要有南方铁杉、红豆杉(Taxuschinensis)、香榧(Torreyagrandis)、鹅掌楸(Liriodendronchinensis)、川黄檗(Phellodendronchinensis)等21种重点野生植物保护对象[25]。
1.2 样地设置
根据史密斯桑尼亚热带研究所的热带森林科学研究中心(The Center for Tropical Forest Science, CTFS)样地建设标准[26],2014年于江西武夷山国家级自然保护区的核心区黄岗山设置了南方铁杉针阔混交林动态监测固定样地,中心点坐标为27°50′26" N,117°45′43" E,固定样地面积为6.4 hm2(400 m×160 m),运用全站仪将其分成160个20 m×20 m的样方。2015年8月在每个20 m×20 m样方内对胸径(DBH)≥1 cm的木本植物进行物种鉴别、坐标定位、胸径测量和编号挂牌,记录每个调查对象的胸径、树高、冠幅等指标[24]。在每个20 m×20 m样方中,随机设置16个1 m×1 m的小样方,记录每个小样方内草本种名及盖度等指标。样地调查结果如表1.

表1 江西武夷山固定样地基本情况表
1.3 种群动态特征计算方法
1.3.1年龄结构
种群年龄结构不仅可以反映种群不同个体的组成情况,也反映了种群数量动态及发展趋势,对其分析是揭示种群结构现状和更新策略的重要途径之一[27]。由于南方铁杉生长缓慢,缺少解析木资料,因此以立木级结构代替种群年龄结构分析种群数量动态[28]。本研究中种群径级结构为:胸径5 cm以下的南方铁杉为第1龄级,胸径5—10 cm为第2龄级,胸径10—20 cm为第3龄级,之后以10 cm为一间隔,以及类推至100 cm以上为第12龄级数。根据南方铁杉生长规律[29],其中1—4龄级为幼龄级,5—8龄级为中龄级,9—12龄级为老龄级。
1.3.2种群结构动态量化分析
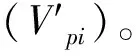
式中,Sn与Sn+1分别为第n与第n+1年龄级种群个体数,K为最大龄级。
1.3.3静态生命表和生存分析
根据吴承祯等[31]的计算方法对南方铁杉静态生命表进行计算和绘制,且采用“匀滑技术”解决生命表绘制时出现死亡率为负的情况。同时本文引入生存分析中的4个函数:生存函数S(t)、累积死亡率函数F(t)、死亡密度函数f(t)、危险率函数λ(t)[32],用于分析南方铁杉的种群动态。
1.3.4种群数量动态的时间序列预测模型
采用线性平稳时间序列分析中的一阶移动平均模型[33]对南方铁杉种群年龄结构动态进行预测。其模型如下:
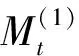
1.3.5空间分布格局
点格局分析广泛应用于固定大样地内的种群空间格局研究,通过种群个体的空间坐标数据分析不同尺度下种群的分布格局。南方铁杉种群在我国数量稀少,多为零星分布,但在保护区内有较大区域的分布,采用点格局分析更为合适。因此使用单变量成对相关函数g(r)分析不同尺度下各龄级的种群空间分布格局。成对相关函数g(r)由Ripley′sK函数衍生[22],计算公式如下:
式中,r为空间尺度,即研究半径,A为固定样地面积,n为样地内南方铁杉种群总数量,uij为种群个体i到个体j之间的距离,Ir为指示函数,Wij为边缘校正系数。
通过100次Monte-Carlo随机模拟得到置信区间为99%的上下两条包迹线,若g(r)值在上包迹线之上,则种群成聚集分布;若g(r)值在上下包迹线之内,则种群成随机分布;若g(r)值在下包迹线之下,则种群成均匀分布。
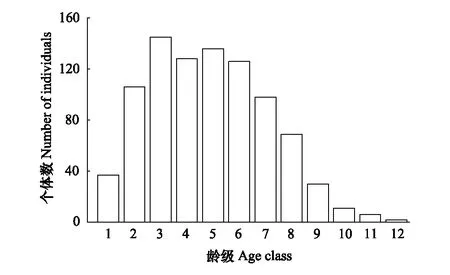
图1 江西武夷山固定样地南方铁杉种群年龄结构 Fig.1 Age structure of T. chinensis var. tchekiangensis population in permanent plot in Jiangxi Wuyishan
1.3.6数据处理与分析
通过Microsoft Excel 2016用于数据校对和整理,并用于静态生命表编算、动态量化分析、生存函数估算和时间序列预测计算。点格局数据计算和分析则使用生态学软件Programita完成,所选空间尺度为0—80 m,步长为1 m。其余数据制图利用Origin 9.0软件完成。
2 结果与分析
2.1 南方铁杉种群年龄结构及动态量化分析
本样地中,南方铁杉种群共894株,各龄级的个体数量如图1所示,种群年龄结构分布总体上呈“金字塔”型,显示南方铁杉种群属于衰退型种群。个体数在第4龄级出现波动,其个体数低于第3及第5龄级,但仍高于第6龄级。种群个体数主要集中在2—8龄级,所占比例约90.38%。


表2 南方铁杉种群龄级结构动态变化指数/%
2.2 静态生命表与存活曲线、死亡率和消失率曲线
南方铁杉种群的静态生命表如表3所示,随着龄级的增加,南方铁杉种群各龄级存活量(lx)逐渐减少,各龄级个体的生命期望(ex)逐渐降低,仅在第10龄级种群个体生命期望出现细微波动,其值略高于第9龄级的个体生命期望。

表3 江西武夷山固定样地南方铁杉种群静态生命表
图2中,南方铁杉种群的存活曲线显示随着龄级增长,lnlx值逐渐减少,总体呈现出“凸”型,表现为Deevey-I型。从图3中可知,南方铁杉种群的死亡率和消失率曲线变化趋势基本一致,皆在第9龄级出现峰值;在第10龄级出现谷值。有可能是南方铁杉种群在9龄级之后进入生理死亡年龄,种群个体快速减少,因此出现一定的波动。
2.3 南方铁杉种群生存分析
表4为南方铁杉种群的4个生存函数估算值,结合图4和图5,可以看出南方铁杉种群的生存率随龄级增加而逐渐下降,累积死亡率逐渐递增,且二者互补。在第9龄级前,二者增加或下降的幅度较大,其中,在第7—9龄级之间幅度最大,第9龄级之后,生存率S(t)和累积死亡率F(t)变化幅度有所放缓。

图2 南方铁杉种群存活曲线 Fig.2 Survival curve of T. chinensis var. tchekiangensis population

图3 南方铁杉种群死亡率(qx)和消失率(Kx)曲线 Fig.3 Mortality rate (qx) and vanish rate (Kx) value curve of T. chinensis var. tchekiangensis population
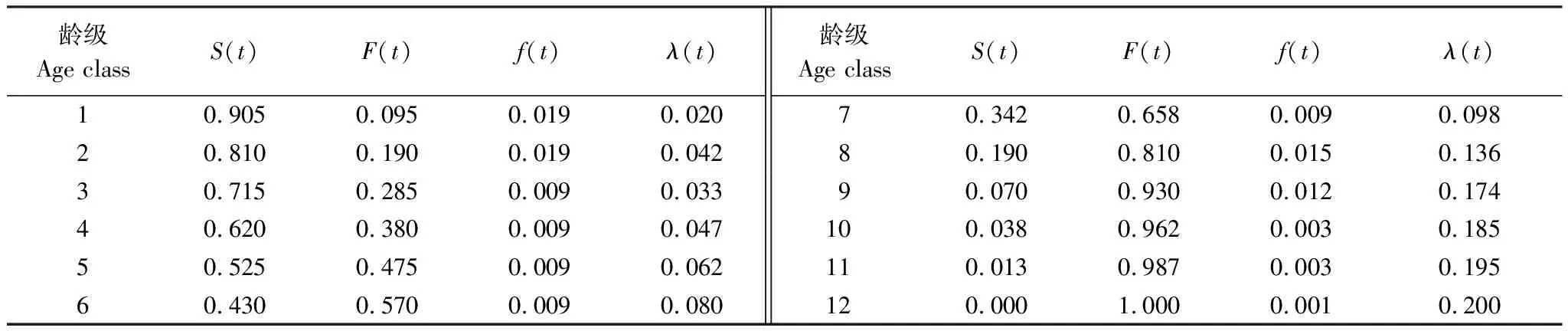
表4 南方铁杉种群生存函数估算值
死亡密度f(t)整体偏低,在第1和第2龄级出现最大值,为0.019,最小值出现在第12龄级,为0.001,总体波动幅度较小。危险率λ(t)曲线总体呈现上升的趋势,在第2龄级出现波动,其值高于第3 龄级而低于第4龄级。第7—9龄级上升幅度较快,第3—7、9—12龄级上升幅度较低。
2.4 种群时间序列分析
利用时间序列预测模型对未来3、6、9个龄级时间后南方铁杉个体数进行推演,得出表5。表中可知,在3个龄级时间之后,第3、第4龄级个体数降低,第5龄级个体数持平,第6龄级之后个体数的预测值皆上升。在6个或者9个龄级之后,仅第6、9龄级的个体数降低,其余龄级个体数皆增加。总体来说,幼龄树不足导致不同龄级的时间后南方铁杉个体数出现少许波动。

表5 南方铁杉种群数量的时间序列分析
2.5 空间分布格局
南方铁杉种群在不同龄级的空间分布格局如图6所示。南方铁杉幼龄树g(r)值皆高于上包迹线,在0—80 m的尺度下都呈聚集分布。中龄树聚集程度较老龄树相比降低,在0—1 m的尺度下呈随机分布,在1—80 m尺度下显示为聚集分布。老龄树g(r)值皆处于两条包迹线之间,在0—80 m的尺度下呈随机分布。样地中南方铁杉种群的所有个体g(r)值趋势与幼龄树类似,皆呈聚集分布。
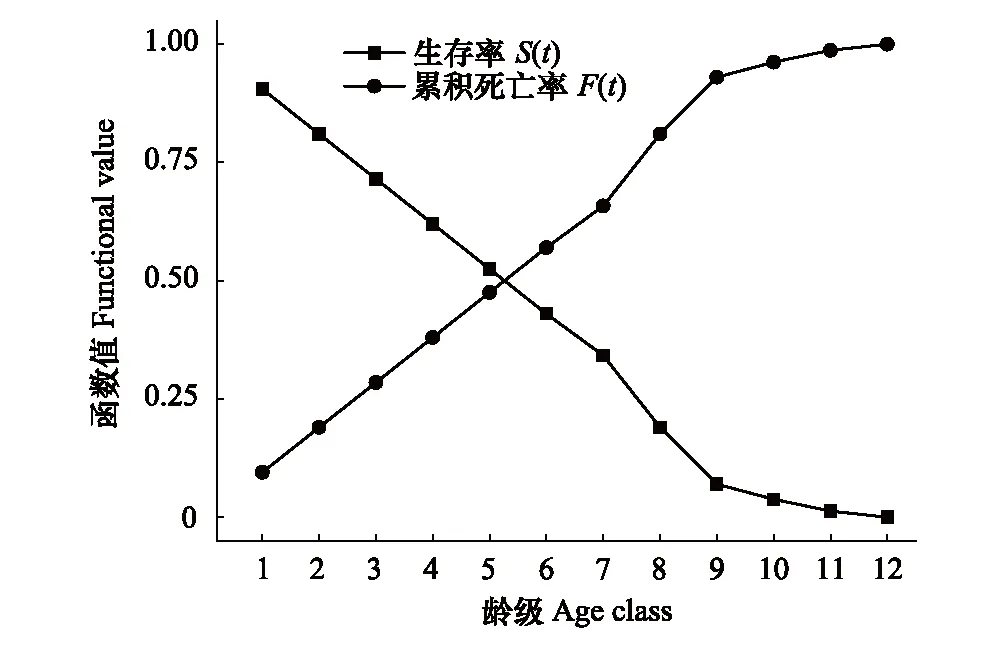
图4 南方铁杉种群生存率函数(S(t))和累积死亡率(F(t))函数曲线Fig.4 Survival rate (S(t)) and cumulative death rate (F(t)) curves of T. chinensis var. tchekiangensis population

图5 南方铁杉种群死亡密度(f(t))和危险率(λ(t))函数曲线Fig.5 Mortality density (f(t)) and risk functional (λ(t)) curves of T. chinensis var. tchekiangensis population
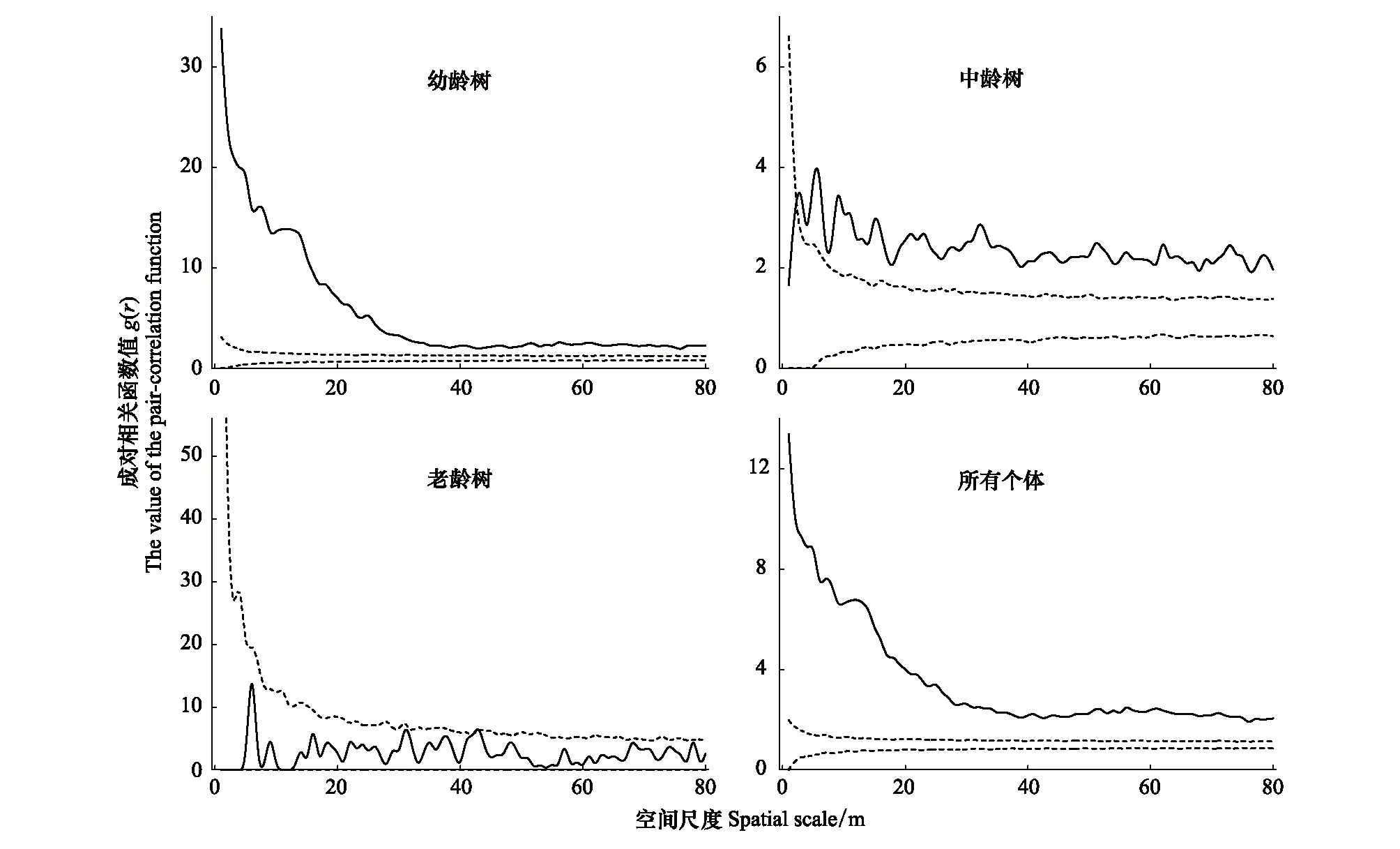
图6 南方铁杉种群空间点格局分析Fig.6 Point pattern analysis of T. chinensis var. tchekiangensis population图中实线为g(r)函数值,虚线为Monte-Carlo随机模拟得到的上下两条包迹线
3 结论与讨论
3.1 南方铁杉种群动态和更新能力
南方铁杉属阳性慢生树种,在早期生长较慢[29],一旦摆脱不利条件,南方铁杉的生长将趋于稳定。本样地中南方铁杉种群的径级结构总体呈“金字塔”型,属于衰退型种群,主要在幼龄时存在衰退状态。南方铁杉种群存活曲线呈现出Deevey-I型,目前属于稳定型种群,与李林等[16]对广西猫儿山及郭连金等[15]在武夷山猪母坑地区等对南方铁杉的调查研究类似,再结合时间序列预测分析,表明现阶段南方铁杉各龄级个体数量在经过不同龄级时间后略有增长,易出现波动,种群自然更新能力较差。这与浙江九龙山[18]和湖南桃源洞梨树洲[17]的南方铁杉种群研究结果相反,其原因可能是本研究固定样地高海拔且是人为干扰较少的原始林,林下的高郁闭的现状无法得到改善,不利于南方铁杉生长。
生存分析函数能很好地表现种群的结构和动态变化,结果表明南方铁杉种群具有前期缺乏、中期波动、后期减少、总体衰退的特点。祁红艳等[34]认为南方铁杉生长缓慢、林下受压“早衰”、甚至死亡是南方铁杉缺乏竞争优势、更新困难的原因。树种在不同生长阶段对资源和空间的需求和耐性会表现出差异性[35],南方铁杉在林下不具有优势,无法获得充足的空间的光照等条件导致其幼苗和幼树的生态位宽度较低[11];郭连金[15]发现南方铁杉种子散布能力弱,自然迁移能力低,且休眠期较长,这也使样地中个体数量不足。随着龄级的增长,生长期的南方铁杉对光照、空间等资源的需求越来越迫切,出现强烈的环境筛;另外,由于群落中种内和种间竞争的加强,尤其是林冠层多脉青冈、青冈等大乔木的竞争[11],导致种群数量在中龄级出现波动;后期老龄树伴随着南方铁杉进入生理死亡年龄,虽然具有较发达的根系,但仍无法提供老龄树庞大的生物量[29],因此种群进入衰退阶段。
3.2 南方铁杉种群的空间分布
植物种群分布格局主要受物种自身特性、种群相互关系及外部生境差异性的影响,可以反映种群个体在水平空间上彼此的关系及其在群落中的作用和地位[36]。已有研究表明随着种群年龄的增加,种群的空间格局的聚集程度逐渐降低,表现为由聚集分布向随机分布转变[37],同样,南方铁杉种群在不同生长阶段也会呈现不同的分布状态[6]。南方铁杉幼龄树和中龄树在样地基本呈聚集分布,老龄树呈随机分布,且幼龄树的聚集程度比中龄树大,这很可能由南方铁杉的种子扩散机制、种内或种间竞争决定。南方铁杉种子扩散能力较差[15],使得幼苗幼树多聚集在母树周围,表现出较高的聚集程度。
随着龄级的增长,环境筛作用对种群动态的影响越发明显,呈聚集分布的幼龄树在生长过程中受到环境因子、种内种间竞争的干扰后个体数量锐减,逐渐降低聚集程度,最后呈随机分布。种群进入老龄阶段后呈随机分布,是因为此时南方铁杉多为大径级个体,对周围环境干扰的抵抗能力较强,具有稳定的资源获取和空间占用能力,生长或存活主要受自身生理特性影响,该结果也与本文种群动态特征的研究结果一致。
3.3 南方铁杉种群保护策略
南方铁杉种群在多地出现“断层”的现象[15—17],将会导致这一种群趋于濒临灭绝,但是至今仍没有切实有效的保护政策。江西武夷山的南方铁杉种群动态特征和空间格局显示该种群不具备自我更新和维持的能力和潜力,其更新依赖于母树周围充足的种子源。原始林林冠层的高度郁闭性现状和来自林下植被的持续性种间竞争,很有可能对该地区的南方铁杉种群造成不可逆的影响。
近年来,我国对濒危植物的保护研究集中在两个方面,一是重点考虑濒危植物的种群动态监测和遗传多样性保护[38]。二是减轻或消除胁迫性生态因子如人为干扰对珍稀植物种群维持的不利影响[39],而本研究所得结论恰好相反,亟需人为的良性干扰促进南方铁杉幼苗更新。因此,可以在保护南方铁杉现有种群数量的基础上,适度地采取除灌、清理病腐木等科学措施保证林内南方铁杉种子萌发和幼苗定居具有良好环境,为该地区的南方铁杉种群续存创造有利的生境条件。
