华顶杜鹃叶绿体psbA-trnH序列的克隆与分析
2022-06-23汤紫依田盛野何海叶张慧娟
汤紫依,吴 倩,田盛野,何海叶,朱 晏,张慧娟,蒋 明
(台州学院 生命科学学院,浙江 台州 318000)
0 引言
杜鹃属(Rhododendron)是杜鹃花科(Ericaceae)最大的属,全世界约有1200种,除非洲和南美洲外,其他洲均有分布。我国的杜鹃属植物种类较为丰富,约有720种,主要分布在西南和华南地区[1-2]。浙江省的杜鹃属植物种类相对较少,仅19种,它们具有观赏或药用价值,其中的云锦杜鹃(R.fortunei)、猴头杜鹃(R.simiarum)和刺毛杜鹃(R.championiae)等具有很好的观赏性,可用于庭院栽培或园林美化;羊踯躅(R.molle)、映山红(R.simsii)和毛果杜鹃(R.seniavinii)等有一定的药用价值,具有清热解毒、化痰止咳和祛风除湿的功效[3]。华顶杜鹃(R.huadingense)为浙江特有种,最早发现于天台华顶山,后在宁波、金华、临海等地发现新的分布点。华顶杜鹃生于海拔700 m以上的黄山松林或针阔叶混交林中,植株数量十分稀少,为国家Ⅱ级重点保护野生植物、浙江省重点保护野生植物和浙江省极小种群保护植物[3-5]。华顶杜鹃树姿优美,花先于叶,花色艳丽,花朵硕大,具有很好的观赏价值和开发前景(如图1所示)。

图1 华顶杜鹃
叶绿体基因组为双链环状DNA,长度范围为35~217 kb,绿色植物中叶绿体基因组是大小约为115~165 kb、有130个左右编码的基因,包括tRNA、rRNA和蛋白质编码基因等[6]。叶绿体基因及其基因间隔区通常十分保守,经常被用于植物系统发育分析或物种的鉴定,其中的ndhF、matK、atpB、rpl16、trnL内含子、trnK内含子、trnL-trnF和psbA-trnH等应用较广[7-8]。Givnish等[9]结合形态学和ndhF基因序列对泽蔺花科(Rapateaceae)开展研究,明确了它们的进化关系;Hao等[10]利用trnL-trnF理清了红豆杉科(Taxaceae)、三尖杉科(Cephalotaxaceae)和罗汉松科(Podocarpaceae)植物之间的亲缘关系;Feng 等[11]则借助psbA-trnH序列对酸浆属(Physalis)植物进行了分子鉴定。近年来,有关华顶杜鹃的研究主要集中在群落特征、种间关系和亲缘关系等方面,与叶绿体基因间隔区相关的研究未见报道[12-14]。本文在克隆华顶杜鹃叶绿体psbA-trnH序列的基础上进行了序列分析和系统发育分析。
1 实验材料与方法
1.1 实验材料
华顶杜鹃叶片分别采自浙江省临海市兰辽(2019年)、天岗尖(2020年)、括苍山(2021年)和天台县华顶山(2021年),将健康叶片置于自封袋带回实验室。叶片先用大量的自来水冲洗,再用无菌水冲洗3~4次,晾干后置于-80℃超低温冰箱中备用。
1.2 基因组DNA的提取
每个样地取10个单株的叶片用于DNA的提取,基因组DNA的提取采用试剂盒法,“柱式植物基因组DNA提取试剂盒”购自生工生物工程(上海)股份有限公司,操作按照说明书步骤进行。DNA用1%的琼脂糖凝胶电泳检测,再稀释成35 ng/μL的溶液,置于-20℃冰箱中备用。
1.3 psbA-trnH序列的克隆
用于扩增psbA-trnH序列的上游和下游引物分别为5'-CGCGCGTGATGAATTCACAATCC-3'和5'-GTTATGCATGAACGTAATGCTC-3',以DNA为模板进行PCR扩增。在20 μL体系中,分别加入2 μL 10×Taq缓冲液(含 Mg2+)、0.40 μL dNTP mix(10 mmol/L)、上/下游引物(20 μmol/L)各 0.45 μL,DNA 模板 0.8 μL(35 ng/μL)和 0.45 μLTaqDNA 聚合酶(2 U/μL),最后加入 ddH2O 15.45 μL。PCR 反应程序为:95℃ 5 min;95℃ 30 s,65℃ 45 s,72℃ 45 s,共32个循环;72℃延伸10 min。PCR产物经1.0%琼脂糖凝胶电泳检测、连接、转化和PCR验证后测序。
1.4 序列比对和系统发育分析
从 NCBI下载张家界杜鹃(R.zhangjiajieense)(HQ707040)、灰被杜鹃(R.tephropeplum)(JX989278)、银叶杜鹃(R.argyrophyllum)(KM605490)、褐叶杜鹃(R.pseudociliipes)(KM605509)、糠秕杜鹃(R.sperabiloides)(KM605519)、柳条杜鹃(R.virgatum)(KM605526)、毛棉杜鹃(R.moulmainense)(KM605648)、弯果杜鹃(R.campylocarpum)(KM605669)、猩红杜鹃(R.fulgens)(KM605680)、滇隐脉杜鹃(R.maddenii)(KM605702)、米杜鹃(R.tschonoskii)(LC612422)、天城杜鹃(R.amagianum)(LC631172)、羊踯躅(R.molle)(MH837814)和长蕊杜鹃(R.stamineum)(HQ707026)的psbA-trnH序列。采用Clustal X 1.81进行多重比对,参数取默认值;根据比对结果,利用GeneDoc生成序列比对图;利用Mega 7.0计算遗传距离,并生成系统发育树,建树方法采用邻接法,自举检测次数为1000。
2 结果与分析
2.1 华顶杜鹃psbA-trnH序列信息
利用PCR法克隆了华顶杜鹃的psbA-trnH序列,电泳结果表明,PCR产物条带清晰、单一,可用于连接、转化和测序(如图2所示)。测序结果表明:来自同一样地10个单株的psbA-trnH序列完全一致,不同样地样品之间则存在一定的差异。多重比对结果表明:兰辽样品psbA-trnH的序列最长,为353 bp,而华顶、天岗尖和括苍山样品的序列长度一致,均为352 bp:兰辽样品psbA-trnH的GC值为29.2%,另三地样品psbA-trnH的GC值为29.3%。兰辽样品与其他三地样品的psbA-trnH仅存在1个碱基的差异,该变异发生在+260 bp处,兰辽样品在该位点有1个碱基T的插入(如图3所示)。

图2 华顶杜鹃psbA-trnH的克隆

图3 华顶杜鹃样品psbA-trnH序列的比对
2.2 杜鹃属植物psbA-trnH的序列比对
如表1所示,18个杜鹃属植物材料psbA-trnH的序列长度范围为346~359 bp,天城杜鹃和米杜鹃的序列最长,均为359 bp;华顶杜鹃(兰辽)、张家界杜鹃、弯果杜鹃和滇隐脉杜鹃次之,均为353 bp;柳条杜鹃的序列最短,仅346 bp。psbA-trnH序列的GC值大小为24.4%~29.3%,华顶杜鹃的GC值最大,长蕊杜鹃其次,为28.6%,张家界杜鹃的GC值最小,仅24.4%。4个样地华顶杜鹃psbA-trnH的相似性最高,兰辽样品与天岗尖样品、华顶样品、括苍山样品的相似性均为99.7%,与长蕊杜鹃的相似性次之,为97.2%,与张家界杜鹃、银叶杜鹃的相似性最低,均为91.5%。
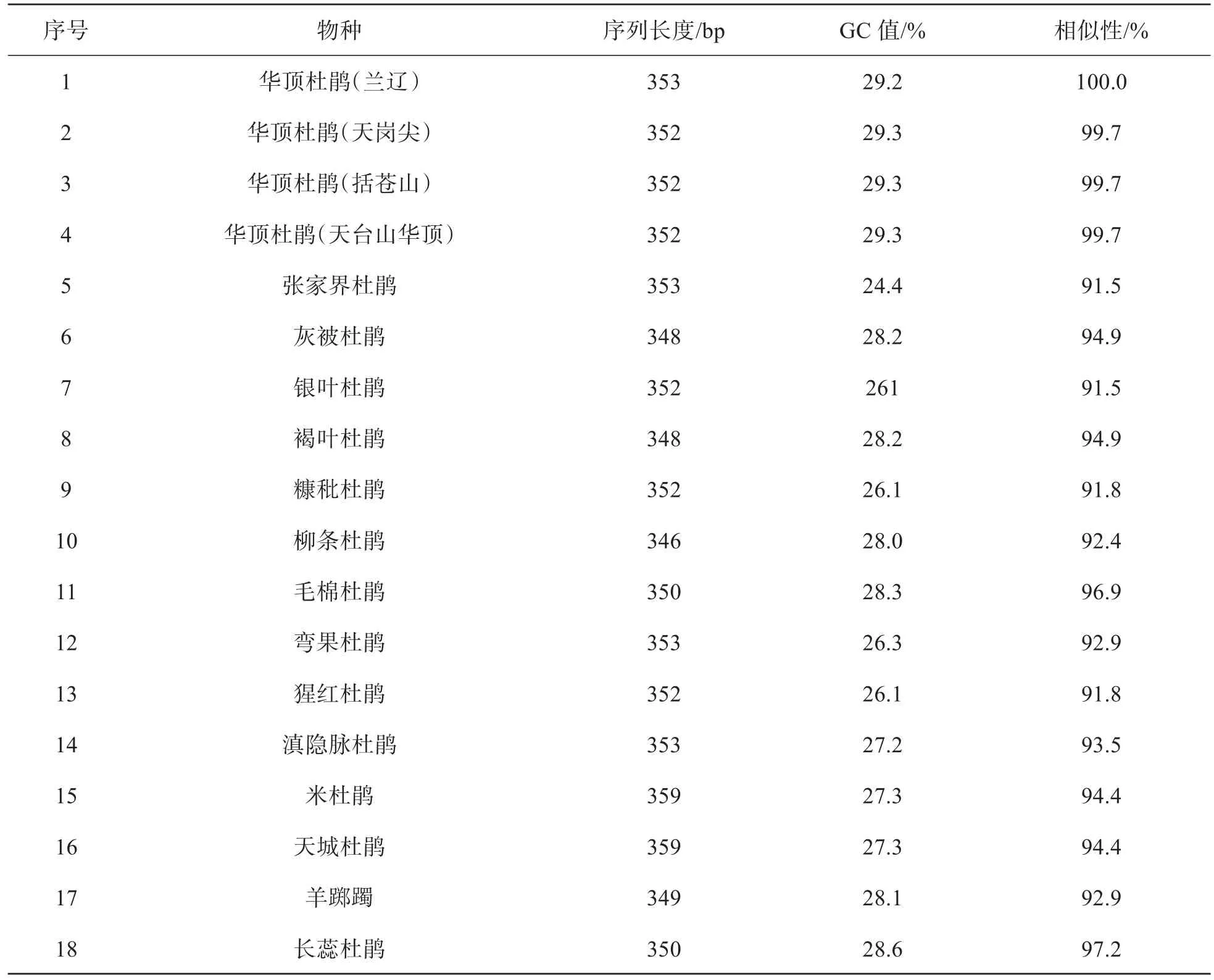
表1 杜鹃属植物psbA-trnH序列的特征
华顶杜鹃序列与其他杜鹃属植物序列之间存在大量的插入、缺失、转换和颠换现象。如长蕊杜鹃、毛棉杜鹃、米杜鹃、天城杜鹃和华顶杜鹃psbA-trnH的+7位为G,而其他杜鹃属植物在该位点的碱基为A,发生了1次转换;羊踯躅的+36位为G,而其他杜鹃属序列为A,该位点也发生了转换现象(如图4所示)。灰被杜鹃和褐叶杜鹃在+55位的碱基为A,而其他杜鹃属序列中为C,发生了颠换现象。滇隐脉杜鹃的+75位发生缺失,银叶杜鹃、糠秕杜鹃、弯果杜鹃、猩红杜鹃、张家界杜鹃的psbA-trnH序列在该位点为T,而其他杜鹃属植物为G,发生了颠换现象。

图4 杜鹃属psbA-trnH序列的多重比对
2.3 系统发育分析
利用Mega软件计算遗传距离,结果表明:18个材料之间的平均遗传距离为0.047,华顶杜鹃与张家界杜鹃的遗传距离最大,为0.084,与柳条杜鹃和滇隐脉杜鹃的遗传距离次之,为0.078;4个样地华顶杜鹃材料之间及灰被杜鹃和褐叶杜鹃之间的遗传关系最近,遗传距离最小,值为0。利用邻接法构建了系统发育树,如图5所示。从图5中可以看出,15种杜鹃在发育树上可分为6组:4个华顶杜鹃材料聚于组I,支持率达99.9%;马银花亚属的长蕊杜鹃和毛棉杜鹃聚于组Ⅱ,支持率相对较低,为69.7%;映山红亚属的米杜鹃和天城杜鹃聚于组Ⅲ,支持率为94.4%;杜鹃亚属的灰被杜鹃、褐叶杜鹃、柳条杜鹃和滇隐脉杜鹃聚于组Ⅳ,支持率为93.3%;羊踯躅亚属的羊踯躅单独聚于组V;常绿杜鹃亚属的5种植物聚于组Ⅵ,支持率为95.3%。

图5 基于psbA-trnH序列构建的系统发生树
3 讨论
叶绿体基因及基因间隔区是植物系统发育分析和分子鉴定的重要DNA条形码,与核基因条形码相比,叶绿体DNA标记为母性遗传,进化速率小,序列组成和长度相对保守,适合种以上水平的系统发生研究,近年来在谱系地理学、系统发育、亲缘关系及遗传多样性等方面得到广泛应用[15]。psbA-trnH是最为常用的叶绿体DNA条形码之一,该序列的长度变异大,变异位点多,在真双子叶植物(eudicotyledons)、单子叶植物、裸子植物、蕨类植物和苔藓植物中的长度范围分别为152~851 bp、151~905 bp、283~1006 bp、167~547 bp和103~265 bp[16]。文中15种杜鹃属植物psbA-trnH的长度变异不大,介于346~359 bp之间,而不同地理来源华顶杜鹃样品的psbA-trnH长度则更为保守,仅存在1个碱基的差异。
psbA-trnH的变异丰富,可用于种及种级以上物种的鉴定。赵容等[17]以穿龙薯蓣(Dioscorea nipponica)及23种来自NCBI数据库的薯蓣属植物psbA-trnH序列为材料,发现利用该DNA条形码可区分供试的24种植物。刘涛等[18]利用psbA-trnH对滇重楼(Paris polyphyllavar.yunnanensis)及其同属物种进行鉴定,表明该序列可用于分辩这些物种,并且有较好的稳定性和准确性。Loera-Sánchez等[19]研究表明:psbA-trnH可用于区分5个供试豆科(Leguminosae)牧草和3种饲用牧草材料,但它不能正确区分羊茅属(Festuca)、黑麦草属(Lolium)和霞禾属(Poa)牧草。本文实验中利用psbA-trnH序列可将华顶杜鹃和其他杜鹃属植物区分开,但该序列不能区分灰被杜鹃和褐叶杜鹃,因为它们的序列长度和碱基组成完全一致,需要其他DNA条形码解决这一问题。
4 结语
本文以珍稀濒危植物华顶杜鹃的叶片DNA为材料,在克隆叶绿体基因组psbA-trnH序列的基础上,利用生物信息学手段分析它的序列特征,明确了华顶杜鹃与其他杜鹃属植物之间系统发育关系,为后续进一步开展华顶杜鹃的遗传多样性研究奠定了基础。
