小鼠不同脂肪组织对冷应激的适应性反应
2022-06-15向李倩
耿 蓉,罗 岩,孙 翠,向李倩,刘 煜
南京医科大学附属逸夫医院内分泌科,江苏 南京 211166
肥胖已成为全球公共卫生危机,中国是全球受肥胖影响人数最多的国家之一,约46%的成年人和15%的儿童肥胖或超重[1-2]。脂肪组织是调节机体能量代谢平衡中重要的靶器官,不仅能够通过能量贮存和释放,还可以通过调节产热、改变局部炎症状态来参与机体能量代谢平衡[3]。冷刺激治疗肥胖的潜在作用包括诱导脂肪组织适应性产热[4]、促进2 型免疫、促进脂肪分解[5]等。
脂肪组织分为以下3 种类型:棕色脂肪组织(brown adipose tissue,BAT)、白色脂肪组织(主要是附睾白色脂肪,epididymal white adipose tissue,eWAT)和米色脂肪组织(主要是腹股沟脂肪组织,inguinal white adipose tissue,iWAT)[6]。BAT 主要由棕色脂肪细胞组成,含有丰富的血管和高度神经支配,是机体非战栗性产热的主要器官[7];eWAT 的主要功能是储存和释放能量,血管和神经分布最少;iWAT 在腹股沟区较为明显,交感神经纤维相对BAT 较少[8]。iWAT 中除脂肪细胞外还存在一些免疫细胞,比如嗜酸性粒细胞、巨噬细胞等[9]。iWAT响应寒冷和其他某些刺激而出现适应性改变,称为适应性产热[6,10],这一过程伴随着神经重塑[11]。在肥胖个体中,脂肪组织呈现出慢性、低度炎症的状态,其中巨噬细胞呈现出促炎性M1型极化的状态[12]。冷刺激后iWAT 中嗜酸性粒细胞、巨噬细胞被选择性激活,选择性激活的巨噬细胞可以通过分泌儿茶酚胺增加脂解、促进适应性产热[13]。与BAT 相比,iWAT 具有更大的表型灵活性,可以根据外界环境调整产热或脂质储存表型。BAT和iWAT能够增加能量消耗、促进产热,有益于探究肥胖及其相关代谢综合征的治疗。
本研究通过观察C57BL∕6J小鼠3种脂肪组织对冷刺激响应的区别,为研究冷刺激条件下脂肪组织功能及作用机制提供基础。
1 材料和方法
1.1 材料
10 周龄雄性C57BL∕6J 小鼠购于南京医科大学医药实验动物中心,体重24~28 g,实验动物的饲养、使用和操作均获得南京医科大学医药实验动物中心伦理委员会批准(伦理审批号为IACUC⁃1901036)。蛋白marker(ThermoFisher 公司,美国),逆转录试剂(南京诺唯赞生物科技股份有限公司),SYBR Green(上海翌圣生物科技股份有限公司),引物由苏州金唯智生物科技有限公司合成。
1.2 方法
1.2.1 动物分组和处理
将小鼠适应性喂养1 周,待小鼠体征稳定后随机分为2 组:对照组和冷刺激组。对照组小鼠喂养于室温(22 ℃)环境;冷刺激组小鼠于6 ℃环境饲养,两组均单笼单只。
1.2.2 RNA提取及相对定量分析
取20~30 mg 小鼠脂肪组织,加500 μL RNA⁃easy Isolation Reagent 在高速组织研磨仪中研磨,70 Hz,120~180 s,按说明书抽提RNA,OneDrop 微量分光光度计进行定量及纯度分析。
用5×HiScript ⅡqRT Super Mix逆转录为cDNA,-20 ℃保存。
用去离子水10 倍稀释cDNA,实时荧光定量PCR 按照稀释后的cDNA 4.6 μL,SYBR Green 5 μL,上、下游引物(10 mmol∕L)各0.2 μL,混匀,反应在ThermoFisher Quant Studio5荧光定量PCR仪上进行,反应程序为:预变性95 ℃5 min;95 ℃10 s,60 ℃30 s,40 个循环;熔解曲线:95 ℃15 s;60 ℃1 min,95 ℃15 s。以2-ΔΔCT计算目的基因相对表达量。
目的基因和内参引物如下:解偶联蛋白1(un⁃coupling protein 1,Ucp⁃1)上游引物5′⁃AGGCTTC⁃CAGTACCATTAGGT⁃3′,下游引物5′⁃CTGAGTGAG⁃GCAAAGCTGATTT⁃3′;细胞死亡诱导DNA 片段化因子样效应子A(cell death inducing DFFA like effec⁃tor A,Cidea)上游引物5′⁃TGCTCTTCTGTATCGCC⁃CAGT⁃3′,下游引物5′⁃GCCGTGTTAAGGAATCT⁃GCTG⁃3′;过氧化物酶体增殖受体γ共激活因子1α(peroxisome proliferator⁃activated receptor gamma co⁃activator 1⁃α,Ppargc1α)上游引物5′⁃AGCCGTGAC⁃CACTGACAACGAG⁃3′,下游引物5′⁃GCTGCATG⁃GTTCTGAGTGCTAAG⁃3′;内参基因β⁃actin上游引物5′⁃GTTGGTTGGAGCAAACATC⁃3′,下游引物5′⁃CTTATTTCATGGATACTTGGAATG⁃3′。
1.2.3 Western blot
各脂肪组织加裂解液后匀浆,冰上充分裂解后,4 ℃12 000g离心10 min,转移上清至新FP 管中。加入5×SDS上样缓冲液,变性后电泳,转膜,室温封闭1 h,一抗过夜,孵育二抗,FCL化学发光法检测蛋白表达。
1.2.4 流式细胞术
脂肪组织用消化酶消化,细胞过筛,离心后重悬1 次,再次离心,Fc 受体封闭液封闭,加入相应抗体anti⁃CD45⁃PF∕Cy7、anti⁃CD11b⁃PerCP∕Cy5.5、anti⁃F4∕80⁃AF488、anti⁃CD11c⁃APC,孵育30 min。流式细胞染色缓冲液洗涤,细胞活性染料染色。离心去染料,重悬细胞,加入固定剂,室温下避光孵育。洗涤后离心弃上清,重悬细胞。加入破膜剂和anti⁃CD206⁃PF,避光孵育后洗涤,离心弃上清,再涡旋使细胞完全重悬,用0.1%甲醛固定,2~8 ℃避光保存,并在24 h内对固定的细胞进行分析。
1.3 统计学方法
所有数据均使用Fxcel 进行统计学分析。定量数据用均数±标准差()进行描述,两组定量数据比较采用双尾t检验,P<0.05 为差异有统计学意义。
2 结果
2.1 冷刺激增加进食量,对体重及脂肪组织重量影响不显著
C57BL∕6J 小鼠冷刺激实验设计如图1A 所示,小鼠适应性喂养24 h后,分别记录对照组和冷刺激组小鼠体重、血糖、原始食物量,冷刺激组小鼠6 ℃、单笼单只饲养;冷刺激48 h 后,记录对照组和冷刺激组小鼠体重、血糖、食物余量;冷刺激70 h 后,安乐死小鼠,分别取肩胛区BAT、腹股沟区iWAT、附睾eWAT。
冷刺激48 h,对照组和冷刺激组体重(图1B)、脂肪组织及肝脏重量(图1C)无显著差异。冷刺激组小鼠进食量显著增加(P=0.006 8,n=5,图1D),这与其他文献报道结果一致[14]。观察各脂肪组织切片的HF 染色结果,发现与对照组相比,冷刺激组BAT 中细胞大小及胞内脂滴无明显变化,iWAT 组织内呈现褐色化改变,eWAT 内脂肪细胞体积略减少(图1F),说明冷刺激显著诱导iWAT褐色化。
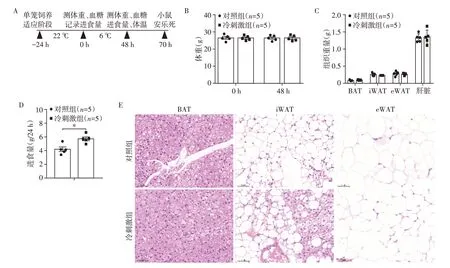
图1 冷刺激后小鼠体重及脂肪组织的变化Figure 1 Changes of body weight and adipose tissues in mice after cold stimulation
2.2 冷刺激后3种脂肪组织产热基因表达有所不同
为了明确冷刺激对不同脂肪组织产热及线粒体功能是否存在不同,对不同脂肪组织进行了Ucp⁃1的免疫组化染色(图2A)。发现在BAT和eWAT中,对照组和冷刺激组Ucp⁃1 无显著差异;而在iWAT中,冷刺激组Ucp⁃1 表达显著高于对照组。通过qRT⁃PCR和Western blot检测了BAT、iWAT和eWAT中不同基因mRNA和蛋白表达水平:在BAT中,qRT⁃PCR结果显示冷刺激组中Ppargc1α、Cidea、肿瘤坏死因子受体超家族成员9(tumor necrosis factor receptor superfamily member 9,TNFRSF9∕CD137)等褐色化相关基因和肉碱棕榈酰转移酶1A(carnitine palmitoyl⁃transferase 1A,Cpt1a)、肉碱棕榈酰转移酶2(carni⁃tine palmitoyltransferase 2,Cpt2)等脂肪酸氧化相关基因的表达显著高于对照组(图2B),而对照组和冷刺激组Ucp⁃1的mRNA和蛋白水平差异均无统计学意义(图2B、C),考虑在3种脂肪组织中,BAT中Ucp⁃1 表达最高,冷刺激不能引起其进一步升高。在iWAT 中,qRT⁃PCR 结果显示冷刺激组Ucp⁃1、Ppargc1α、Cidea 等褐色化相关基因和乌头酸酶2(aconitase 2,Aco2)等线粒体功能相关基因的表达显著高于对照组(图2D);Western blot结果显示冷刺激组Ucp⁃1、CCAAT∕增强子结合蛋白β(CCAAT∕en⁃hancer⁃binding protein β,C∕FBPβ)等产热相关基因和激素敏感型脂肪酶(hormone⁃sensitive lipase,HSL)的表达显著高于对照组(图2F),表明冷刺激能够促进iWAT 产热和线粒体功能显著增加。在eWAT中,qRT⁃PCR结果显示冷刺激组Aco2、ATP 合成酶⁃H+转运⁃线粒体F1复合体α亚基1(ATP synthase,H+transporting,mitochondrial F1 complex,alpha subunit 1,Atp5α1)、琥珀酸脱氢酶B 亚基(succinate dehydrogenase complex iron sulfur subunit B,Sdhb)等线粒体功能相关基因和Cpt2 的表达显著升高(图2F);Western blot 结果显示对照组和冷刺激组在产热相关基因表达上无显著差异(图2G)。以上结果提示冷刺激提高3种脂肪组织线粒体的功能,但仅能提高BAT和iWAT而非eWAT中产热基因的表达。
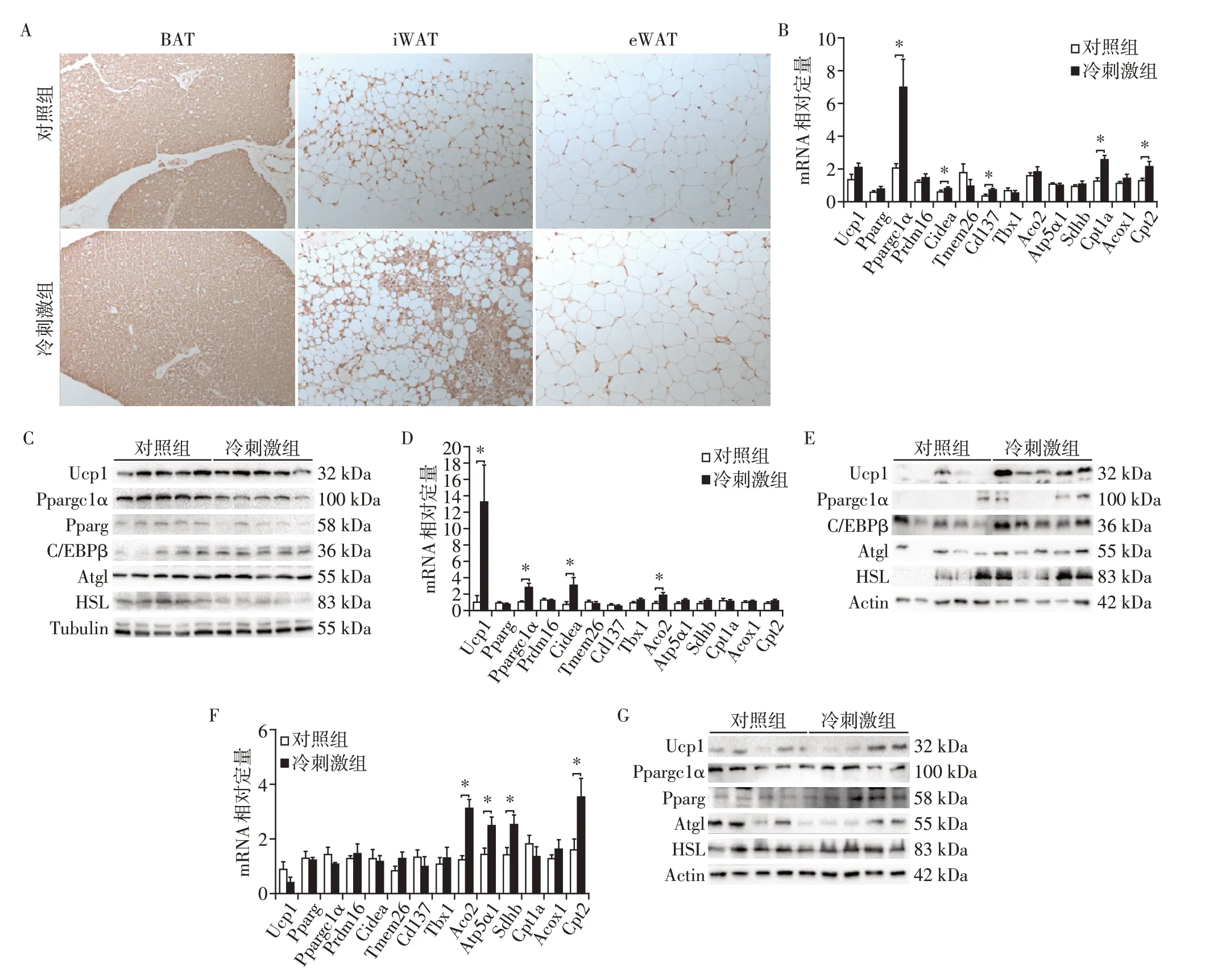
图2 冷刺激后3种脂肪组织产热基因表达水平Figure 2 Expressions of thermogenic genes in three adipose tissues after cold stimulation
2.3 冷刺激后3种脂肪组织交感神经基因表达变化
研究表明脂肪组织产热主要受交感神经释放去甲肾上腺素调节,酪氨酸羟化酶(tyrosin hydroxy⁃lase,TyrH)主要表达于交感神经,是儿茶酚胺类激素生物合成过程中的关键限速酶。免疫组化结果显示,在BAT 和iWAT 中,冷刺激组TyrH 表达显著高于对照组(图3)。
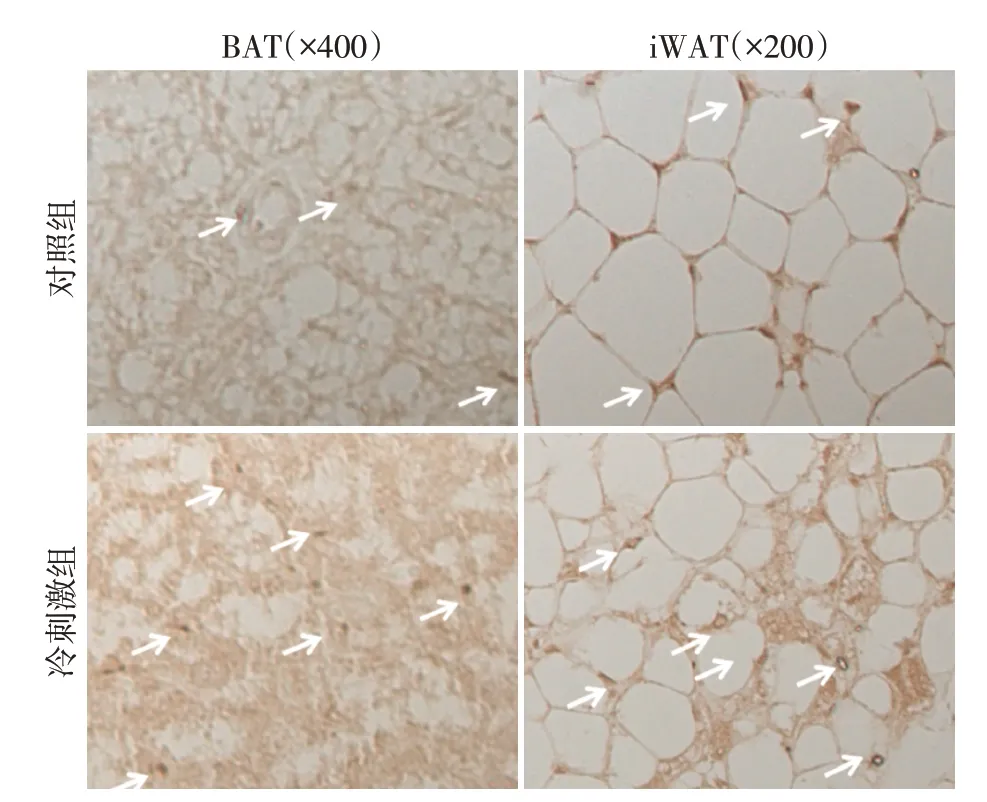
图3 免疫组化检测产热脂肪组织TyrH表达Figure 3 TyrH expression in thermogenic adipose tissues detected by immunohistochemical staining
2.4 冷刺激后3种脂肪组织中巨噬细胞的M2型极化状态变化
巨噬细胞极化在脂肪组织炎症中起着重要作用。M1型巨噬细胞是经典型巨噬细胞,具有促炎活性,其标志物是CD11c[15];M2 型巨噬细胞是选择性激活的巨噬细胞,呈现出抗炎作用,其标志物是CD206[15]。为了明确冷刺激对脂肪组织中巨噬细胞的影响,通过流式细胞仪检测巨噬细胞亚型。与对照组相比,冷刺激组BAT和iWAT中M1型巨噬细胞的比例有所降低(图4A),而M2 型巨噬细胞比例显著升高(图4B),表明冷刺激可以改善BAT 和iWAT中的炎症状态。eWAT 中巨噬细胞极化改变不明显。以上结果表明冷刺激能够促进BAT和iWAT中巨噬细胞M2型极化,对eWAT中巨噬细胞极化影响不大。
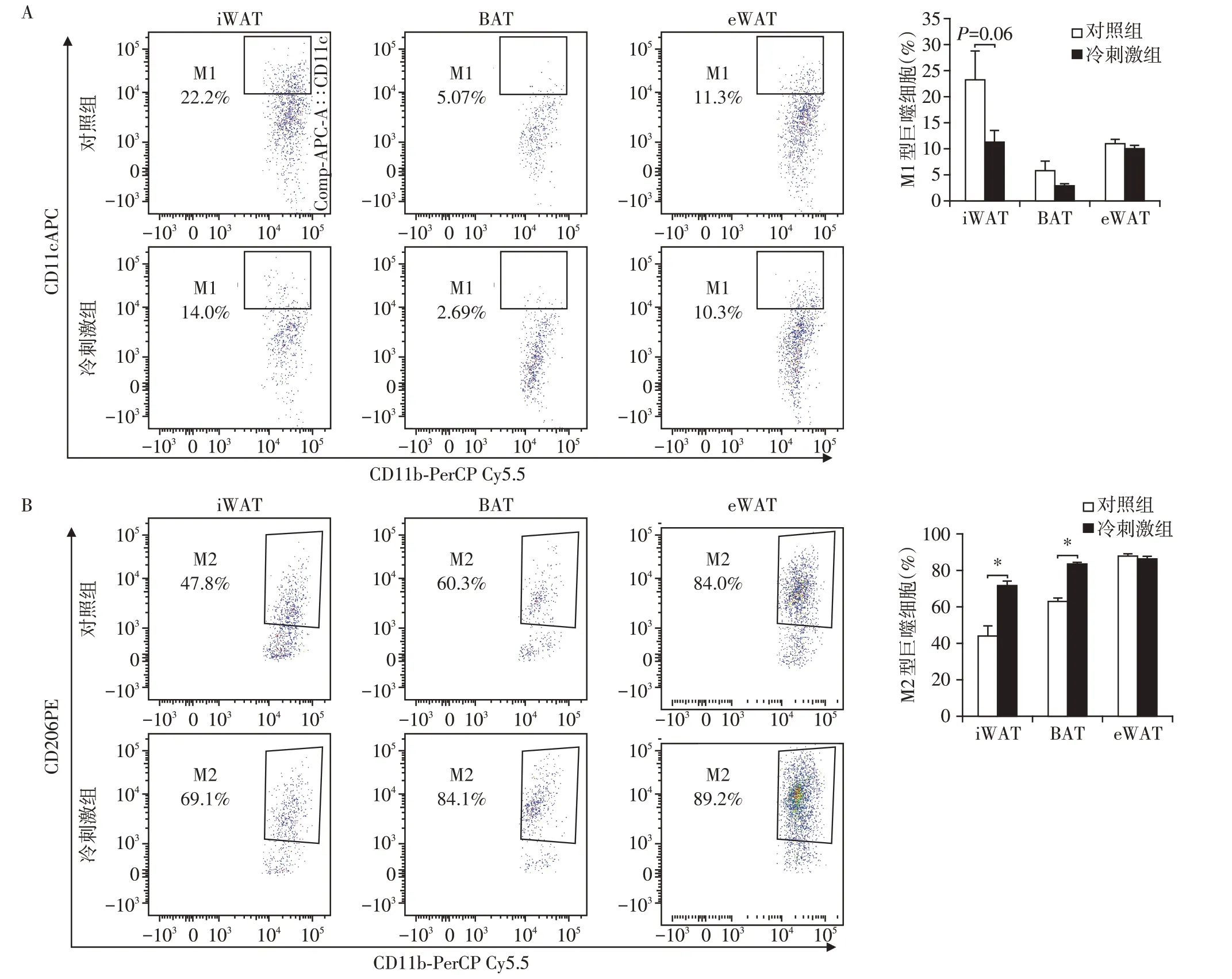
图4 流式分析冷刺激后3种脂肪组织巨噬细胞Figure 4 Flow cytometric analysis of three kinds of adipose tissue macrophages after cold stimulation
3 讨论
本研究旨在探讨C57BL∕6J 小鼠中不同类型的脂肪组织对冷刺激适应性应答的相似点与差异。结果发现3 种脂肪组织对冷刺激的应答有所不同。BAT 作为机体非战栗性产热的主要组织,产热基因Ucp⁃1表达在3种脂肪组织中最高,冷刺激不能进一步提高其表达水平,但组织内交感神经支配显著提高,巨噬细胞M2型极化增加,这可能与产热少量增加有关。iWAT 对冷刺激响应最为积极,产热基因和M2 型巨噬细胞都显著提高,组织内交感神经支配显著增加。eWAT 中产热基因表达和M2 型巨噬细胞比例均无明显变化,对冷刺激响应不显著。考虑冷刺激时间较短,且内脏脂肪的主要作用是储存能量,受环境温度调控较小。冷刺激后所有脂肪组织的线粒体功能都有所增强,但脂肪组织重量和体重无显著变化,考虑冷刺激后小鼠进食量增加,脂肪产热的增加来源于摄入食物的增加。
综上所述,不同类型的脂肪组织在冷刺激后,产热、交感神经支配、脂肪组织巨噬细胞极化方面均存在显著差异,但具体机制还有待进一步研究。此外本研究仅观察了C57BL∕6J 小鼠冷刺激后脂肪组织的差异,未观察肥胖小鼠冷刺激后不同脂肪组织的差异,需进一步造模研究(ob∕ob 鼠或者高脂饮食诱导肥胖[16])。不同脂肪组织对冷刺激不同响应的研究有助于了解脂肪组织在冷刺激后机体代谢过程中的作用,且对研究肥胖与代谢综合征等疾病的诊断、治疗过程极为重要。随着不同类型脂肪组织差异性研究的不断推进,脂肪组织的代谢功能及对机体生理和病理状态影响的了解会更加深入。
