大豆GmNF-YA8的生物信息学及盐胁迫表达分析
2022-06-15李铭杨张军翟莹何佳琦邱爽于海伟陈炯辛马婷婷
李铭杨,张军,翟莹,何佳琦,邱爽,于海伟,陈炯辛,马婷婷
大豆的生物信息学及盐胁迫表达分析
李铭杨1,张军2,翟莹1*,何佳琦1,邱爽1,于海伟1,陈炯辛1,马婷婷1
(1.齐齐哈尔大学 生命科学与农林学院,黑龙江 齐齐哈尔 161006;2.黑龙江省农业科学院畜牧兽医分院,黑龙江 齐齐哈尔 161005)
本实验对大豆基因进行了生物信息学分析和高盐胁迫下表达量的检测。位于大豆基因组9号染色体上,cDNA全长1121bp,开放阅读框615bp,编码204个氨基酸。GmNF-YA8蛋白分子量22.7 kD,pI 6.16。GmNF-YA8蛋白氨基酸序列中含有1个CBF保守结构域。GmNF-YA8蛋白无信号肽,定位在细胞核中,属于亲水性蛋白。蛋白系统进化分析结果表明,GmNF-YA8与拟南芥AtNF-YA4和AtNF-YA7的亲缘关系最近。实时荧光定量PCR结果显示,高盐胁迫处理后的表达量先上升后下降,在处理10h时表达量最高,约为对照组的4倍,表明在大豆幼苗中能够响应高盐胁迫。
大豆;核因子Y;生物信息学;盐胁迫;表达分析
大豆中的蛋白质含量丰富,是谷类作物的两倍左右。中国作为大豆的原产国,一直将大豆作为食用油的主要原料之一[1]。目前,极端温度、土壤盐渍化、干旱等非生物胁迫对大豆产量影响越来越严重。盐胁迫会使细胞渗透压发生改变,导致植物养分流失,不利于植物的生长发育。因此,培育出对土壤盐渍化有抗性的大豆品种意义重大。
核因子Y(Nuclear factor Y, NF-Y)通常也被称为CCAAT结合因子(CCAAT-binding factor, CBF)或亚铁血红素激活蛋白(Heme activator protein, HHAP),由NF-YA、NF-YB和NF-YC三个亚基构成,是一种普遍存在于真核生物中的一个庞大的转录因子基因家族[2]。研究发现NF-Y在植物应对不同环境胁迫(如干旱、盐害、极端温度等)中起着重要作用。目前,已有大量植物NF-YA基因耐盐功能鉴定的相关报道。例如,甘蓝型油菜和基因受盐胁迫的强诱导,且BnNF-YA中的其他成员也不同程度的在盐胁迫下诱导表达[3]。在模式植物拟南芥中,过表达的拟南芥和小麦基因均可以使拟南芥的耐盐性增强[4-5]。沉默棉花和会大幅度降低棉花幼苗的耐盐性[6]。大豆中存在21个NF-YA基因[7]。但目前仅的功能被鉴定,其表达受高盐、干旱、低温和ABA诱导,过表达提高了转基因拟南芥的抗旱性,但对高盐胁迫和外源ABA的敏感性增强[8]。本课组前期发现大豆能被干旱胁迫诱导表达(已另文发表),本研究对的基因及蛋白序列进行生物信息学分析,并对其高盐胁迫下的表达量进行检测,为后续基因的功能鉴定及应用提供理论依据。
1 实验部分
1.1 材料
大豆“北豆9号”种子由齐齐哈尔大学植物分子育种实验室提供。
1.2 生物信息学分析
登陆PlantTFDB(http://planttfdb.gao-lab.org/index.php)在线网站,获得大豆转录因子的基因和蛋白序列。Expasy在线软件(https://web.expasy.org/compute_pi/)预测蛋白分子量及等电点。在线软件PROSITE(https://prosite.expasy.org/)分析大豆GmNF-YA8蛋白的功能结构域。PSORT在线软件(http://cello. life.nctu.edu.tw/)预测GmNF-YA8蛋白亚细胞定位情况。通过网站(https://web.expasy.org/cgi-bin/protscale/)分析GmNF-YA8蛋白的亲/疏水性。使用SignalP-5.0(https://services.healthtech.dtu.dk/)网站预测GmNF-YA8蛋白信号肽;在NCBI(https://www.ncbi.nlm.nih.gov/)数据库获得GmNF-YA3和拟南芥NF-YA1-10的蛋白序列,由MEGA5软件构建系统发育树。
1.3 大豆幼苗盐胁迫处理
将大豆种子播种在沙土和草炭土1∶1均匀混合的花盆中,待大豆幼苗生长至根长约5cm,将其移至Hoagland培养液中,培养至幼苗一片三出复叶彻底展开后开始胁迫处理。用含200 mmol/L NaCl的Hoagland营养液对大豆幼苗高盐胁迫。在胁迫处理的0, 1, 2, 5, 10, 24h收集0.1g的第一片三出复叶(三次重复),-80℃冰箱保存用于后续提取RNA。
1.4 基因表达量的检测
用RNAiso Plus(Takara公司)提取高盐胁迫后各时间点样品的RNA,将其反转录为cDNA(反转录试剂盒来自Novoprotein公司)。根据基因序列,通过Premier 5设计实时荧光定量PCR(qPCR)引物,上游引物5′-AGACAGTCCCGTGCTAAAGC-3′,下游引物3′-CGACCTCCACATCCTCTTGC-5′。用上述实验得到的cDNA为模板,大豆为内参基因,-引物同文献[9]。通过qPCR检测基因在高盐胁迫下的相对表达量,反应体系和程序设置同文献[10]。
2 结果与讨论
2.1 GmNF-YA8生物信息学分析
2.1.1 GmNF-YA8基因及蛋白序列
基因(Genebank登录号NM_001362821)位于大豆第9号染色体。序列分析结果如图1所示,基因cDNA全长1121bp,开放阅读框615bp,编码204个氨基酸,预测蛋白分子量22.7kD,pI 6.16。对蛋白序列功能结构域预测结果显示,GmNF-YA8蛋白在107-140位含有一个CBF结构域,此结构域是基因作为NF-YA转录因子家族一员的标志[11]。对GmNF-YA8蛋白进行信号肽结构预测,结果显示该蛋白无信号肽。

图1 GmNF-YA8核苷酸及氨基酸序列
下划线为CBF结构域;阴影为核定位信号;*为终止密码子
2.1.2 GmNF-YA8蛋白亚细胞定位分析
亚细胞定位预测结果表明,GmNF-YA8含有一个核定位信号(图1)。如表1所示,GmNF-YA8蛋白定位在细胞核的分值最高(4.622),推测GmNF-YA8蛋白定位在细胞核,这与大部分NF-YA蛋白的亚细胞定位结果相同[12]。也有NF-YA蛋白定位于细胞膜上的报道,例如SiNF-YA6蛋白定位在细胞膜和细胞核上,推测定位在细胞膜上该蛋白会被其他蛋白修饰后进入细胞核内发挥作用[13]。
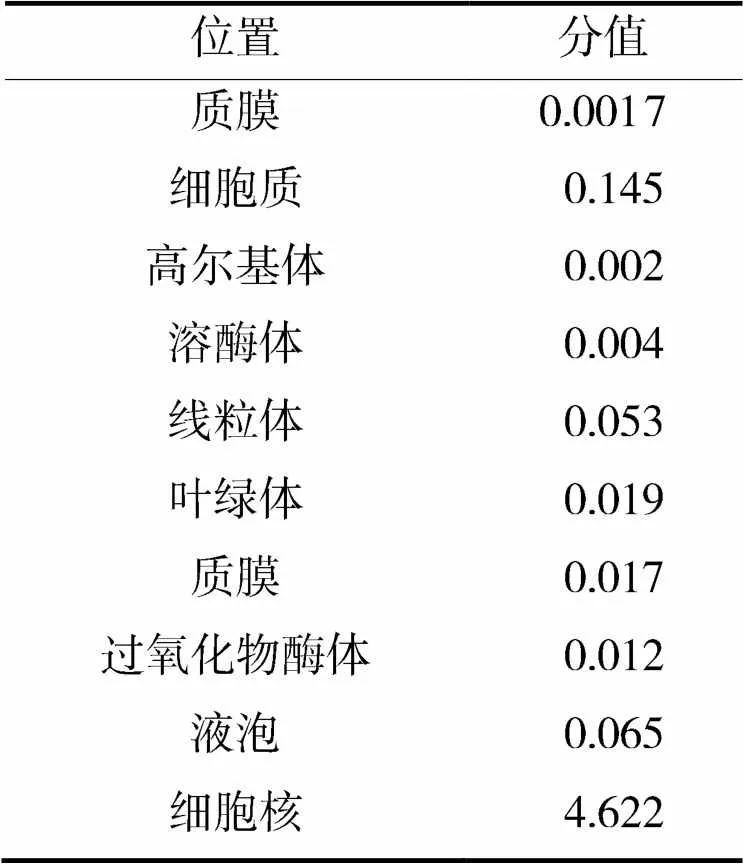
表1 GmNF-YA8蛋白亚细胞定位预测
2.1.3 GmNF-YA8蛋白亲/疏水性分析
对GmNF-YA8蛋白进行亲/疏水性分析,结果如图2所示,GmNF-YA8蛋白疏水性最强的位点是第92位(1.444),亲水性最强的位点是第173位(-3.589),GmNF-YA8蛋白的多肽链大多分布在0刻线以下区域。由此表明GmNF-YA8蛋白具有很强的亲水性,属于亲水蛋白。
2.2 NF-YA蛋白系统发育分析
将大豆GmNF-YA3、GmNF-YA8与10个拟南芥NF-YA蛋白构建系统进化树。如图3所示,GmNF-YA8与AtNF-YA4和AtNF-YA7的亲缘关系最近,由此推测它们的功能可能存在相似性。此外,GmNF-YA8与同为大豆的GmNF-YA3不在同一分支,亲缘关系较远,因此推测NF-YA转录因子家族的分化或许先于拟南芥与大豆之间物种的分化[14]。
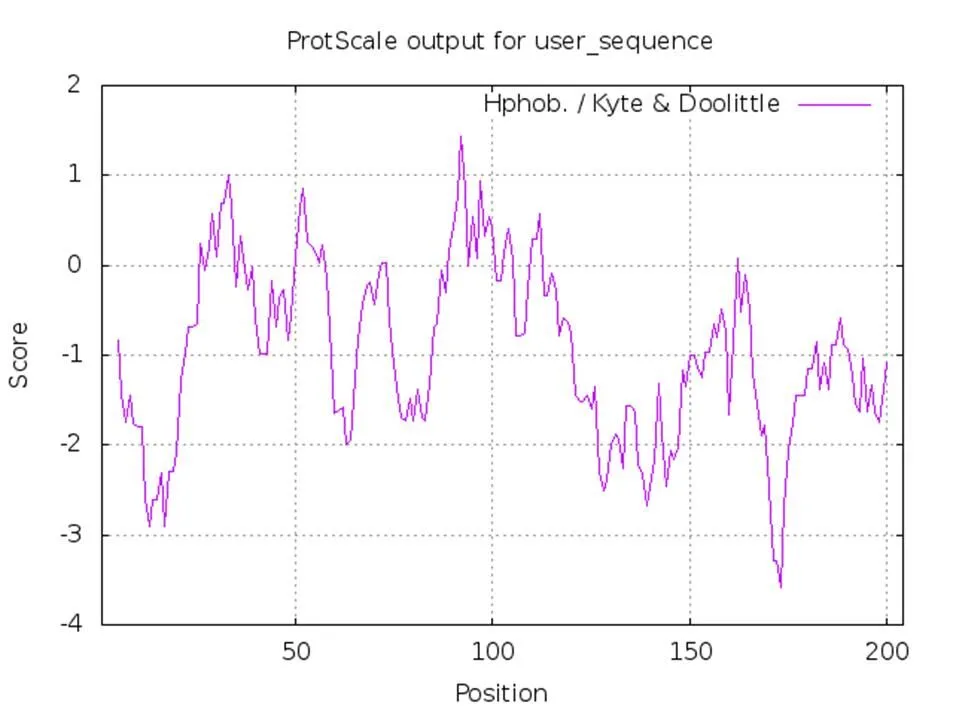
图2 GmNF-YA8蛋白亲/疏水性分布曲线
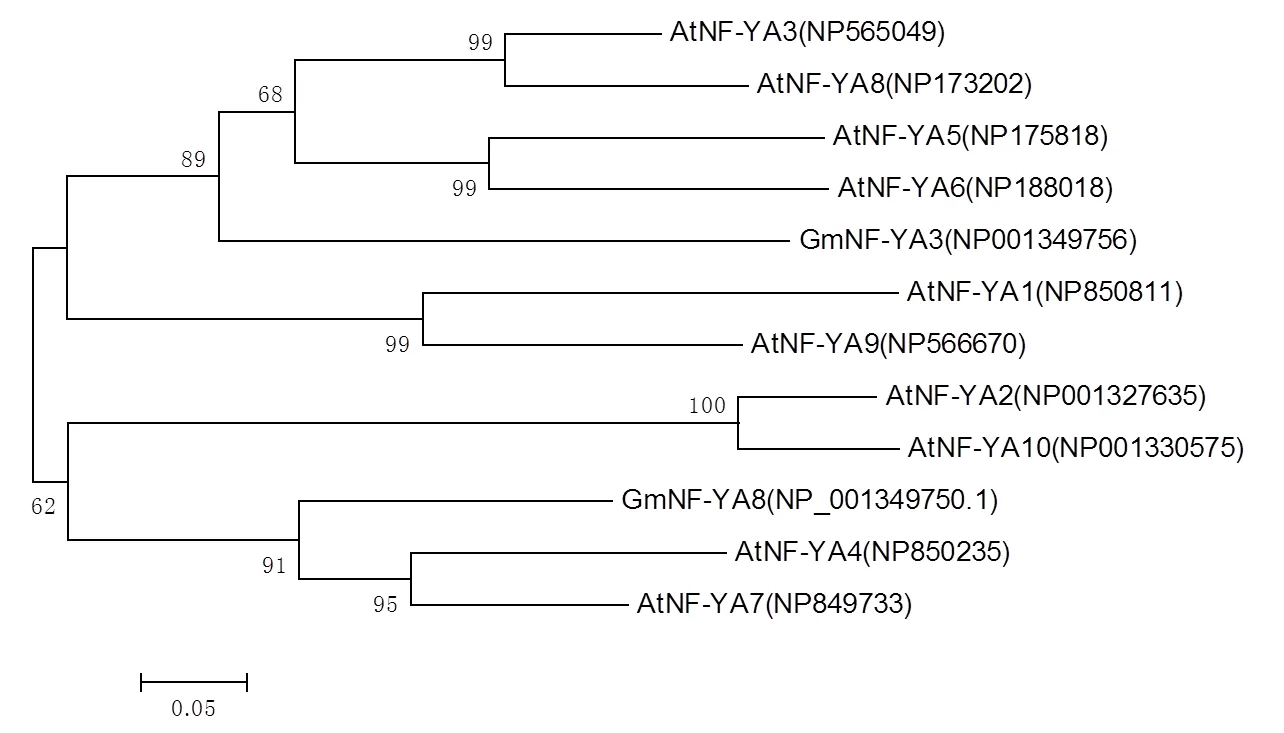
图3 植物NF-YA蛋白系统进化树
2.3 GmNF-YA8在高盐胁迫下的表达
以高盐胁迫处理0h的大豆幼苗为对照,通过qPCR对在盐胁迫各时间点的表达量进行检测。结果显示(图4),在盐胁迫下的表达量呈先上升后下降的趋势,在处理10 h表达量出现峰值,大约是对照组的4倍。这一结果与狗牙根在盐胁迫下的表达情况相似[15],但谷子在盐胁迫处理1~24h的表达量是逐渐升高的,并无下降趋势[13]。由此推测NF-YA转录因子家族成员在不同物种中对盐胁迫的响应机制存在差异。
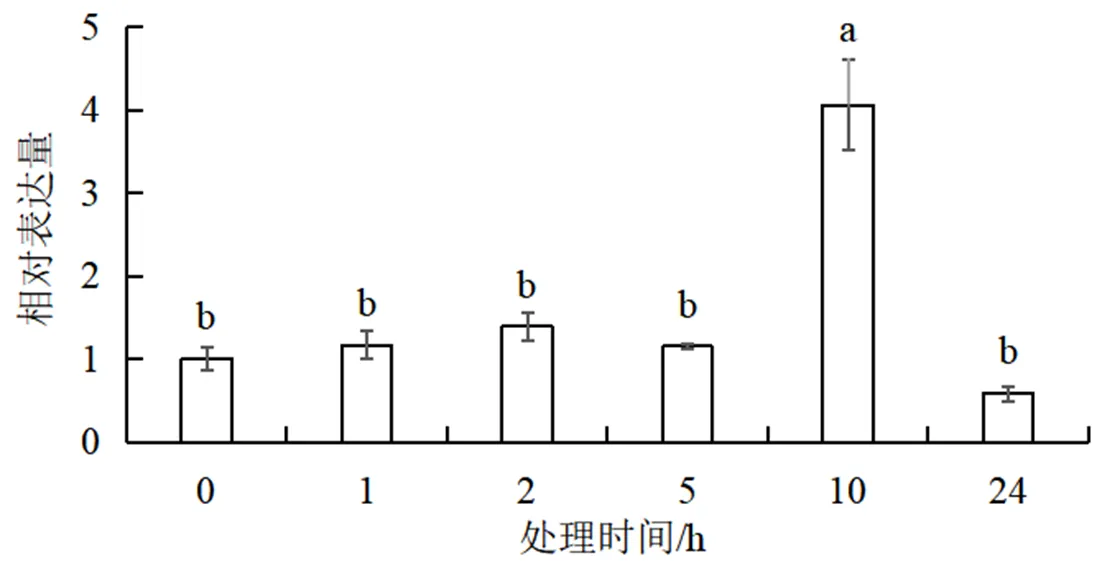
图4 GmNFYA8在高盐胁迫下的表达
不同字母表示差异显著(小于0.05)
3 结论
位于大豆基因组9号染色体上,开放阅读框615bp,编码204个氨基酸。蛋白分子量22.7kD,pI 6.16。GmNF-YA8蛋白无信号肽,定位在细胞核中,属于亲水性蛋白。GmNF-YA8与AtNF-YA4和AtNF-YA7的亲缘关系最近。高盐胁迫处理后,的表达量先上升后下降,表明在大豆幼苗中能够响应高盐胁迫。
[1] 周新安. 我国大豆生产与科研现状及其发展对策[J]. 作物杂志,2007(6): 1-4.
[2] MANTOVANI R. The molecular biology of the CCAAT-binding factor NF-Y[J]. Gene, 1999, 239(1): 15-27.
[3] 林钟员. 甘蓝型油菜NF-YA和NF-YC转录因子的克隆及其表达分析[D]. 南京:南京农业大学,2014.
[4] LI Y J, FANG Y, FU Y R, et al.is involved in regulation of postgermination growth arrest under salt stress in Arabidopsis[J]. PLoS ONE, 2017, 8(4): e61289.
[5] CHEN M, ZHAO Y, ZHUO C, et al. Overexpression of a NF-YC transcription factor from bermudagrass confers tolerance to drought and salinity in transgenic rice[J]. Plant Biotechnology Journal, 2015, 13(4): 482-91.
[6] ZHANG Q, ZHANG J, WEI H, et al. Genome-wide identification of NF-YA gene family in cotton and the positive role ofandin salt tolerance[J]. International Journal of Biological Macromolecules, 2020, 165(PB): 2103-2115.
[7] QUACH T N, NGUYEN H T, VALLIYODAN B, et al. Genome-wide expression analysis of soybean NF-Y genes reveals potential function in development and drought response[J]. Molecular Genetics and Genomics, 2015, 290(3): 1095-1115.
[8] NI Z Y, HU Z, JIANG Q Y, et al., a target gene of miR169, is a positive regulator of plant tolerance to drought stress[J]. Plant Molecular Biology, 2013, 82(1-2): 113-129.
[9] QIU S, ZHANG J, HE J, et al. Overexpression of, a soybean galactinol synthase gene, enhances transgenic tobacco drought tolerance[J]. Plant Cell Tissue and Organ Culture, 2020, 143(3): 507-516.
[10] 翟莹,何佳琦,张军,等. 大豆转录因子基因的克隆及非生物胁迫诱导表达分析[J]. 分子植物育种,2020, 18(18): 5894-5900.
[11] 汪晓东,陈洋,于月华,等. 大豆基因结构及原核表达分析[J]. 作物杂志,2015(4): 47-50.
[12] 白玉翠,王萍,倪志勇,等. 大豆基因克隆及特征分析[J]. 大豆科学,2019, 38(5): 719-725.
[13] 方广宁,胡利芹,王二辉,等. 谷子转录因子的过表达提高转基因植株对低氮胁迫的抗性[J]. 中国农业科学,2015, 48(20): 3989-3997.
[14] 张军,尹珺伊,王岩,等. 大豆GeBP转录因子家族基因生物信息学分析[J]. 齐齐哈尔大学学报(自然科学版),2021, 37(3): 58-61.
[15] 刘芳兵. 狗牙根基因响应盐胁迫的功能解析[D]. 武汉:中国科学院大学(中国科学院武汉植物园),2020.
Bioinformatics and salt stress expression analysis of the soybean
LI Ming-yang1,ZHANG Jun2,ZHAI Ying1*,HE Jia-qi1,QIU Shuang1,YU Hai-wei1,CHEN Jiong-xin1,MA Ting-ting1
(1.College of Life Science and Agroforestry, Qiqihar University, Heilongjiang Qiqihar 161006, China; 2.Branch of Animal Husbandry and Veterinary of Heilongjiang Academy of Agricultural Sciences, Heilongjiang Qiqihar 161005, China)
In this research, the bioinformatics analysis of soybeanand its expression levels under high salt stress were carried out.was located on chromosome 9 of the soybean genome. Its full-length cDNA was 1121bp with an open reading frame of 615bp and encoded 204 amino acids. The molecular weight of GmNF-YA8protein was 22.7 kD and pI was 6.16. The amino acid sequence of GmNF-YA8protein contained a CBF conserved domain. GmNF-YA8protein had no signal peptide and was located in the nucleus, which was a hydrophilic protein. Phylogenetic analysis showed that GmNF-YA8 is the most closely related toAtNF-YA4 and AtNF-YA7. Real-time fluorescence quantitative PCR results showed that the expression levels ofelevated firstly and then decreased after high salt stress treatment. The peak expression occurred 10 h after treatment, which was about 4 times that of the control group. These results indicated thatcould respond to high salt stress in soybean seedlings.
soybean;nuclear factor Y;bioinformatics;salt stress;expression analysis
2021-11-24
黑龙江省普通本科高等学校青年创新人才培养计划(UNPYSCT-2017153);齐齐哈尔大学研究生创新科研项目(YJSCX2020041)
李铭杨(1998-),女,黑龙江绥化人,本科,主要从事植物分子遗传育种研究,lmy879463529@163.com。
翟莹(1982-),女,吉林吉林人,教授,博士,主要从事植物分子遗传育种研究,fairy39809079@126.com。
S565.1
A
1007-984X(2022)03-0070-04
