稻鱼共生系统农田边界的植物和丛枝菌根真菌调查*
——以首批全球重要农业文化遗产稻鱼共生系统核心保护地浙江青田龙现为例
2022-06-10周静怡赵璐峰唐建军裘子悦娄李懿陈超巍林佳辰胡亮亮
周静怡,赵璐峰,唐建军,裘子悦,娄李懿,陈超巍,吴 静,林佳辰,郭 梁,胡亮亮,陈 欣
(浙江大学生命科学学院生态学系 杭州 310058)
联合国粮农组织(FAO)组织和实施的全球重要农业文化遗产系统(Globally Important Agricultural Heritage Systems,GIAHS)计划,旨在建立全球重要的传统农业及其相关的景观、生物多样性、知识和文化保护体系,并在世界范围内得到认可与保护。稻鱼共生系统是人类利用稻田浅水环境将水稻()和鱼类种养在同一稻田空间中而形成的独特农业系统,分布在浙南地区的“青田稻鱼共生系统”具有千年历史,2005年被联合国粮农组织等国际机构列为首批“全球重要农业文化遗产”。研究表明,青田稻鱼共生系统保持高的水稻品种多样性和田鱼遗传多样性,但青田稻鱼共生系统在野生植物资源及其相关生物的保育方面是否也有重要作用? 有多大作用? 这个问题仍需深入研究。
农业系统中,边际土地(如荒地、草坡、田埂等)发挥了重要作用。边际土地不仅构建了农田景观多样性,同时也是野生物种相对富集的地点,并作为生物多样性富集的“野生生境岛屿”保存在农业区域中,对农业生态系统具有重要的生态意义。农田边界是边际土地的主要类型,即农田过渡带,通常由草带、篱笆、树、沟渠、堤等景观要素组成。研究表明,农田边界是农业系统野生植物、昆虫和微生物的重要栖息地,维持有丰富的物种多样性。农田边界植物多样性的维持有助于天敌和传粉者的保育。研究还表明,农田边界的多样性有利于提高农田系统的生态服务功能,例如,由人工混播多种禾本类和非禾本类植物构建的边界植被带的长期存在,能减少恶性杂草的危害。
丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)是土壤中一类重要共生真菌,AMF 的宿主植物分布十分广泛,包括苔藓植物、蕨类植物、裸子植物和被子植物,能与地球上80%的陆地植物形成共生关系。研究表明,AMF 帮助宿主植物扩大对营养元素尤其是难移动的元素(如磷等)的吸收,促进宿主植物生长和提高宿主植物的抗逆能力,AMF 在维持植物群落多样性和稳定性方面也具有重要作用。因而AMF 物种多样性及功能多样性状况已成为衡量生态系统可持续性的一项重要指标。宿主植物类群、土壤生境、农业措施(如耕作、施肥和农药等)均对AMF 物种组成和群落结构产生影响。在宿主植物的影响方面,由于不同种宿主植物生理代谢、根系内环境及分泌物不同,必然会影响不同AMF 的侵染和共生体的形成,进而影响AMF 群落物种的组成。此外,植物群落组成会显著影响AM 真菌的多样性,如Börstler 等发现不同管理类型高山草甸的AMF 群落的变化与地上植物多样性显著相关;Hausmann 等发现,邻体植物及建植次序均会影响AM 真菌群落组成及多样性;Alguacil 等对西班牙东南部半干旱区灌木丛的AM 真菌群落研究发现,灌丛群落复杂性调控着AMF 的多样性。土壤条件也显著影响AM 真菌群落的组成和多样性,如Landis 等发现,AMF 多样性和物种丰富度与土壤氮含量呈正相关;Fitzsimons 等和Liu 等发现,土壤pH 和土壤硝态氮能够很好地预测AM 真菌群落的变化;Alguacil 等研究报道土壤磷有效性与AM 真菌多样性呈负相关。农业措施(如耕作、施肥和农药等)对AMF 多样性的影响也受到关注,如Santos 等研究发现,无机氮肥的使用降低了AM真菌的多样性;Zeng 等发现氮素施用显著改变AMF 群落。农田边界的特殊生境下,AMF 群落组成和物种多样性状况如何? 目前相关研究报道较少。
本研究对青田稻鱼共生系统农田边界的植物和根际土地的土壤丛枝菌根真菌进行调查研究,旨在从农田边界的角度,了解青田稻鱼共生系统对生物多样性的保育状态,揭示全球重要农业文化遗产系统的生物多样性保护意义。
1 材料与方法
1.1 研究区概况
本研究在青田稻鱼共生系统核心保护地浙江省丽水市青田县方山乡龙现村(120°18′E,27°59′N)进行(图1)。该地属亚热带季风气候,年平均气温17~18 ℃,年平均降水量1400~1450 mm,主要种植一季稻。该地所属及周边5 个县的4.5 万hm稻田中,大约1 万hm用于稻鱼共生。本研究区域土地总面积300 hm,稻鱼共生类型的稻田有约30 hm。

图1 研究区域地理位置图(a: 浙江省行政区域,陆上部分;b: 青田县区域;c: 龙现村内本次调查研究的区域)Fig.1 Geographic location map of the study area (a: administrative region of Zhejiang Province,land part only;b: Qingtian County area;c: the study area in Longxian Village)
1.2 样地和样方设计
2020年8月在稻鱼共生系统农田边界采样。梯田海拔高度在300~360 m,田块平均面积344.84 m,农田边界平均宽度(田埂宽度) 61.33 cm,平均高度(塝坡高度)为123 cm,边界面积占梯田总面积的8.49%。在山坡的底部、中部和顶部选择大小相对一致的稻鱼共生梯田为样地,坡底、坡中和坡顶的样地各3 个,共9 个样地(图2a)。
1.3 植物样方调查、植物根系和土壤样本采集
在每个样地(梯田田块)的边界内布设3 个1 m×1 m 样方(图2b),记录每个样方中植物的种类及盖度。样方内的每种植物取3~5 个代表性植株的根系作为样本,根样放入冰盒带回实验室。样方内土壤样本用5 点取样法取5~20 cm 土层的土壤,混匀约20 g,装入无菌自封袋做好标记,放入冰盒(内有冰袋)带回实验室。部分土样风干后过100 目筛用于土壤理化性质分析,部分土样存于-80 ℃用于DNA 提取,高通量测序鉴定。
图2 采样样地、样方分布图(a)和样方位置(b)Fig.2 Diagram of sampling settings in Longxian Village,Qingtian,Zhejiang,China
1.4 丛枝菌根真菌侵染率测定
采用酸性品红染色法对菌根进行染色,植物根系经过10% KOH 透明、3% HCl 酸化后置于酸性品红乳酸甘油染色液中染色30 min。菌根侵染率的测定采用网格交叉法,计算公式为: 菌根侵染率=菌根侵染的交叉点数/根段与方格线总交叉点数×100%。
1.5 丛枝菌根真菌测定
采用DNA 测序技术对根际土壤中AMF 群落进行研究。使用Fast DNASPIN Kit (MPBIO,USA)试剂盒从根际土壤中提取总DNA。提取的总DNA 浓度和纯度用NanoDrop 2000 UV-vis 分光光度计(Thermo Scientific,Wilmington,USA)测定,DNA 质量用2%琼脂糖凝胶电泳检测。经检测确认的总DNA 样品,送上海美吉生物Majorbio公司进行AMF 群落组成分析(https://www.i-sanger.com/)。鉴定AMF 群落组成分析方法: 首先使用真菌通用引物对AML1F_AML2R(F 端引物AML1: 5′-ATCAACTTTCGATGGTAGG ATAGA-3′;R 端引物AML2: 5′-GAACCCAAACA CTTTGGTTTCC-3′),然后用丛枝菌根真菌特异性引物对AMV4.5NF_AMDGR (F 端引物AMV4.5N: 5′-AA GCTCGTAGTTGAATTTCG-3′;R 端引物AMDG: 5′-CCCAACTATCCCTATTAATCAT-3′)进行巢式PCR扩增。Thermocycler PCR 系统(Gene Amp 9700,ABI,USA)程序如下: 第一轮: 95 ℃,3 min 预变性;再95 ℃30 s 变性,55 ℃ 30 s 退火,72 ℃ 45 s 延长,循环32次;最后在72 ℃条件下延长10 min。第二轮: 95 ℃条件下延长3 min;然后95 ℃ 30 s 变性,55 ℃ 30 s 退火,72 ℃ 45 s 延长,30 个循环;最后在72 ℃条件下延长10 min。委托上海美吉生物Majorbio公司通过Illumina MiSeq 平台(Illumina,San Diego,USA)测 定AMF 的DNA。
1.6 根际土壤碳氮磷钾含量测定
土壤理化性质测定依据NY/T 1121-2006 土壤检测标准进行。重铬酸钾氧化-外加热法测定土壤有机质、土壤有机碳含量;碱解扩散法测定土壤速效氮含量;氟化铵-盐酸溶液浸提,钼锑抗比色法测定土壤有效磷含量;乙酸铵浸提-火焰光度计法测定土壤速效钾含量;硫酸-加速剂消解,凯氏法测定土壤全氮含量。
1.7 数据分析
1)物种丰富度指数()。用单位面积的物种数目,即物种密度来表示物种的丰富程度。一般用每平方米的物种数目表示。

式中:p为总物种数中第物种的比例,用每种植物的种群盖度代替;为群落中的总物种数。

式中:表示两个群落中共有的物种数,和分别表示两个群落中特有的物种数。
结果使用Microsoft Excel 2010 和SPSS Statistics 23 进行数据统计和分析。
AMF 群落组成分析由上海美吉生物Majorbio公司完成。对测得序列进行拼接(FLASH 软件)和质控(Trimmomatic 软件),以测定最小样本序列数为标准进行抽平,并按照97%相似性对非重复序列(不含单序列)进行OTU (operational taxonomic unit)聚类(Usearch 7.0 软件)。使用Quantitative Insights Into Microbial Ecology (QIIME,16.0),采用RDP classifier 贝叶斯算法对97%相似水平的OTU 代表序列进行分类学分析,AMF 基因序列比对maarjam081/AM数据库(https://www.maarjam.botany.ut.ee)。AMF 群落组成利用R(4.0.5)软件分析,计算OTU 的相对丰度,即每个样本中序列数占总序列的百分比,并计算反映微生物群落多样性的指数(包括Ace 指数、Chao 指数、Simpson 指数和Shannon 指数),具体计算方法参照Callahan 等。
利用软件SPSS Statistics 23 计算边界植物群落与AMF 群落的多样性指数的相关系数。
使用R 的Pheatmap (1.0.12)软件包绘制热图,Vegan (2.5-7)软件包进行冗余分析(RDA/CCA 分析),使用Car(3.0-13)软件包进行VIF 方差膨胀因子分析筛选剔除共线性的环境因子,使用Vegan 软件包进行db-RDA 分析。
2 结果与分析
2.1 农田边界植物种类
研究区域农田边界样方中共发现维管植物109 种,分属于53 科97 属(见二维码中的附录1)。主要有菊科(Compositae)(11 属)、禾本科(Poaceae)(11 属)、唇形科(Labiatae) (5 属)、茜草科(Rubiaceae) (4 属)、蔷薇科(Rosaceae) (3 属)、凤尾蕨科(Pteridaceae E.D.N.Kirchn) (3 属)。从植物盖度的角度分析(图3),农田边界植物主要有乌蕨()、芒萁()、荩草()、白茅()等。农田边界植物资源较丰富,不同坡位的多样性均较高(表1),且分布较均匀。从α 多样性指数(物种丰富 度=0.995,=0.398;Shannon-Wiener=0.431,=0.660;Simpson=0.291,=0.752;Pielou=0.229,=0.799)、β 多样性指数(Jaccard=6.255,=0.130;Whittaker=4.997,=0.155;Sorenson=4.997,=0.155)看,不同坡位间无显著差异(表2)。

图3 农田边界主要植物种的盖度Fig.3 Coverages of main plant species in the field margin

表1 不同坡位农田边界植物群落α 多样性指数Table 1 Species α diversity indexes of vegetation community in the field margins in different positions of hillside

表2 不同坡位农田边界植物群落β 多样性指数Table 2 Species β diversity of vegetation community in the field margins in different positions of hillside
2.2 植物根系的AMF 侵染率
通过对青田龙现稻鱼共生系统农田边界常见植物的根样进行染色和显微镜检测,发现植物根部均有侵染。不同植物的AMF 侵染率存在显著差异(<0.01),样本的侵染率范围为6.67%~44.44%,平均为27.33%,其中积雪草()的侵染率最低(6.67%),络石()、斑鸠菊()的侵染率最高(44.44%)(表3)。

表3 农田边界植物的丛枝菌根真菌(AMF)侵染率Table 3 Arbuscular mycorrhizal fungi (AMF) colonization rates of plants in the field margin
2.3 农田边界的土壤特征
表4为根际土壤理化性质测定结果,参照第二次全国土壤普查推荐的土壤肥力分级标准,发现稻鱼共生系统农田边界的土壤肥力均较高。对照土壤肥力分级指标可得,所调查地的全氮含量均大于2 g·kg,有机质均大于40 g·kg,属于一级肥力;速效钾含量均大于150 mg·kg,属于二级肥力。单因素方差分析结果表明,仅不同坡位的土壤含水率存在显著差异(<0.01)。

表4 农田边界根际土壤碳氮磷钾特征Table 4 Soil properties of field margin
2.4 农田边界的土壤AMF 群落
群落组成分析结果表明,样本中检测到的OTUs分属于球囊菌门、球囊菌纲的5 目9 科14 属。在科分类水平上(图4a),按照相对丰度大小依次为: 球囊霉科(Glomeraceae,87.29%~89.64%)、只分类到球囊菌纲的物 种(unclassified_c_Glomeromycetes,3.32%~4.19%)、类球囊霉科(Paraglomeraceae,2.88%~4.46%)、巨孢囊霉科(Gigasporaceae,0.38%~1.78%)、无梗囊霉科(Acaulosporaceae,0.85%~2.00%)、其他(0.35%~1.85%)。
在属分类水平(图4b),球囊霉属(,87.29%~89.64%)相对丰度最高,是优势属。其他按照相对丰度大小依次为: 只分类到球囊菌纲的物种(unclassified_c_Glomeromycetes,3.32%~4.19%)、类球囊霉属(,2.88%~4.46%)、无梗囊霉属(,0.85%~1.92%)、其他(0.73%~2.17%)和巨孢囊霉属(,0.38%~1.55%)。对AMF 群落丰富度和多样性指数的分析表明,基于OTU 的物种丰富度指数和多样性指数在不同取样地(坡底、坡中、坡顶)间无显著差异(表5)。
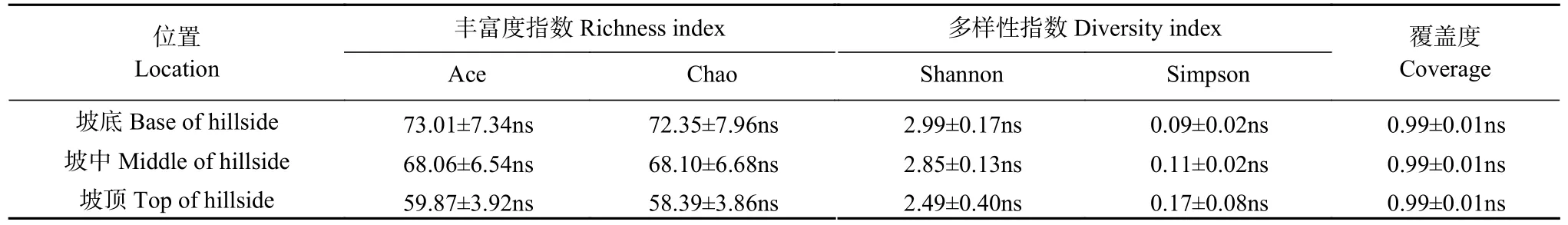
表5 土壤丛枝菌根真菌(AMF)群落丰富度与多样性指数(基于OTU)Table 5 Species richness and diversity indexes of arbuscular mycorrhizal fungi (AMF) community in field margin (based on OTU)
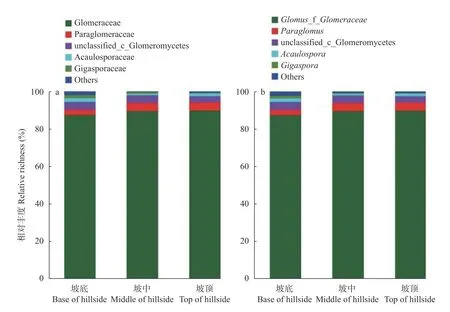
图4 农田边界土壤科(a)、属(b)分类水平上丛枝菌根真菌(AMF)群落组成图Fig.4 Species composition diagram of arbuscular mycorrhizal fungi (AMF) community at the taxonomic level of family (a) and genus (b) in soil of field margin
AMF 群落的多样性与其宿主植物多样性有一定的关系(表6),AMF 群落的Shannon 指数与植物物种丰富度指数表现为极显著正相关(<0.01)。

表6 植物物种丰富度和丛枝菌根真菌(AMF)群落Shannon 指数相关性Table 6 Correlation between plant species richness and Shannon index of arbuscular mycorrhizal fungi (AMF) community
不同环境因子对AMF 群落不同物种的影响存在差异。土壤性状(水分含量、总氮、有效碳、有机碳、有机质、有效磷、有效钾)对AMF 群落中不同物种(基于属水平)影响的热图结果如图5所示,无梗囊霉属()与土壤含水率(MC)存在显著正相关(<0.05)。多孢囊科球囊霉属(f)与土壤总氮(TN) (<0.01)、土壤有效氮(AN) (<0.01)、土壤有机碳(OC) (<0.05)、土壤有机质(OM) (<0.05)均显著正相关。双型囊霉属()与TN (<0.05)、多孢囊霉属()与土壤有效磷(AP) (<0.05)均存在显著负相关。

图5 丛枝菌根真菌群落物种组成在属水平(a)、OTU 水平(b)上与土壤环境因子的Spearman 相关系数热图Fig.5 Spearman correlation coefficient heat maps between species composition of arbuscular mycorrhizal fungi (AMF) community classified at genus level (a) and OTU level (b) and soil environmental factors
而OTU 水平显示AMF 群落中较多物种与速效磷(AP)显著相关(<0.05)。OTU49 与AN 极显著负相关(<0.001),与OC 和OM 呈显著负相关(<0.01)。同时AN 对OTU94、OC 和OM 对OTU94 也有极显著或者显著负影响。
通过方差膨胀因子分析对所有土壤性状进行共线性分析发现,有机碳和有机质之间以及总氮和速效氮之间存在明显的共线性关系,基于这一点,剔除有机质、速效氮后再进行后续分析。又因Axis lengths=2.4105 (<3.0),故采用RDA 分析。结果表明,土壤速效磷对AMF 群落组成的影响最大(<0.05),而速效钾影响较小。对OTU 贡献度较大的主要是球囊霉属。
3 讨论和结论
本研究表明,全球重要农业文化遗产青田稻鱼共生系统的农田边界有丰富的植物种类(见二维码中的附录1),有C植物和C植物、直根系植物和须根系植物、豆科和非豆科植物、禾草类植物和双子叶草本植物等。农田边界的植被组成是由土壤特性、景观结构和农事管理及发展历史等多种因素共同决定的。农田边界植物组成影响植被响应和效应性状,并维护不同植被类型多样性效应特征及其生态系统服务。水资源条件较好的水渠边植物多样性指数和生态优势度较高,而经常受人类活动干扰的作物边界植物多样性指数和生态优势度均显著小于其他农田边界。在本研究中,稻鱼共生系统由于生物种间的互惠效应,田鱼的取食活动对稻田杂草起着很好的抑制效应,降低了除草剂的使用,从而间接地保育了田埂植被多样性。本研究的样地主要是坡地田埂,位于农田外部边缘,是人为修筑的高出田面的台阶与其外侧坡的复合体,主要作用是界定和稳定田面、拦蓄水土、增加耕地水分、提高农田土壤肥力。此外,青田稻鱼共生系统边界的水资源相对较充沛,植物资源较丰富。从坡底到坡顶,植物群落多样性虽然有降低的趋势,但不存在显著差异,这与梯田上方为覆盖良好的森林系统,具有良好的水土保持和水源涵养功能有关。
不同生境AMF 群落的状况,已对湿地、喀斯特地区、岩溶区、荒漠带等进行了较多研究,农业生态系统中AMF 分布特征也有相关研究报道,但农田边界AMF 特征的研究较少。本研究从农田边界根际土壤共分离出AM 真菌9 科14 属62 种,AMF群落物种丰富(图4)。宿主植物是影响AMF 群落物种多样性的重要因素,虽然AMF 和宿主植物间的共生关系并没严格的专一性,但AMF 对宿主植物具有一定的偏好性,并会随着宿主植物演化而发生变化。一方面,不同种类AMF 对于同种宿主植物的作用效应不同,另一方面,同种AMF 对不同种类宿主植物的作用方式和程度也不同,因而植物的功能类群如C和C植物、直根系和须根系植物等均对AMF 群落产生不同的影响。研究还表明,植物多样性和生物量是AMF 多样性的最有力预测因子,植物α 多样性较高时,AMF 共生程度显著增加。在我们的研究样地中,农田边界生境保持有丰富的植物种类(图3,二维码中附录1)和较高的α 多样性(表1)。且AMF 群落的Shannon 指数与植物物种丰富度指数表现为极显著正相关(表6)。因此,农田边界较高的植物物种多样性可能是AMF 物种多样性较高的决定因素。田埂植物有助于加固梯田田埂,物种多样性起到促进田埂植物群落稳定性的作用,也有利于梯田田埂的保护;而多样化的AMF 群落,与植物多样性形成了良好的互惠共生,促进了植被群落的稳定和梯田田埂的保护。我们的研究还发现,AMF 优势属是球囊霉属,这一结果与其他很多研究一致。球囊霉属为AMF 类型最大的群体,适应性强,有较强产孢能力和侵染植物能力。
土壤含水量和有效N/P 比是影响AMF 多样性和群落结构的关键因素,海拔、土壤pH 的变化也是影响AMF 多样性的因素。有研究报道,AMF多样性和物种丰富度与土壤氮含量呈正相关,而与土壤磷有效性呈负相关。本研究中,农田边界的土壤碳、氮、磷、钾含量较高,且与AMF 物种多样性指数均呈正相关(图5),RDA 分析显示,土壤有效磷对AMF 物种多样性影响最大。可见,农田边界根际土壤的AMF 群落组成和多样性主要受宿主植物和土壤特性两方面的影响,较高的植物物种多样性和较高的土壤碳氮含量可能是农田边界保育有较高AMF 物种多样性的主要原因。
现代集约化农业系统农田边界简化,生境和生物多样性下降,相比之下,传统农业系统保留有多样化的农田边界。本研究对青田稻鱼共生系统核心区的研究表明,农田边界保育着多样性的宿主植物和与之共生的丛枝菌根真菌,启示着农田边界生境保护的重要意义。在现代农业系统中,设计和保留适当的农田边界生境,将有利于丰富农业生态系统的生物多样性并发挥其功能。
