利用基因编辑技术研究BRANCHED1参与油菜分枝过程的调控
2022-06-09OlalekanAmoo胡利民翟云孤范楚川周永明
Olalekan Amoo 胡利民 翟云孤 范楚川 周永明
(1.华中农业大学作物遗传改良国家重点实验室,武汉 430070;2.湖北洪山实验室,武汉 430070)
植物分枝是指叶腋处的腋生分生组织起始并形成腋芽,腋芽快速生长发育形成侧枝的过程。分枝是植物株型的一个重要特征,参与植物很多重要的生长过程,如植物叶面积的展开和分布,决定了光的截取和光合作用,进而影响花和果实的数量、果实的填充和产量[1-2];分枝还会影响植物与野草竞争光源、影响害虫的传播等[3-4]。此外,对于一些观赏植物,分枝决定了其观赏品质。
油菜是我国重要的油料作物,其单株产量由单株有效角果数、每角果粒数和千粒重三大产量因素构成。植株株型形态特征如分枝数、分枝位置等性状可以间接影响油菜产量。其中单株有效角果数与分枝密切相关,同时分枝还决定地上部分株型的形态。降低油菜分枝位置、增加有效分枝数不仅有利于植株抗倒性,还可以增加油菜单株产量[5-6]。目前关于油菜株型性状的研究还较少,挖掘可利用的理想株型相关基因在油菜高产育种和实现机械化收割中具有重要的意义。
近些年,在分离和鉴定直接参与植株株型形成的基因方面的研究取得了较多进展,特别在与控制腋芽起始和生长、茎伸长和花序构型相关的基因研究方面。这些株型调控基因大部分在双子叶植物和单子叶植物之间都是保守的,说明这些植物可能通过相似调控途径来调控株型[7]。在控制植物分枝的基因中,最突出的是TEOSINTE BRANCHED1(TB1),它的同源基因在许多物种中都非常重要[8-9]。TB1同源基因在水稻中被称为OsTB1或FINECULM1;在拟南芥、豌豆和番茄中被称为BRANCHED1(BRC1)。众所周知,BRC1基因在腋芽中特异表达,被认为是不同信号调控植物分枝的重要枢纽[10-14]。拟南芥中具有两个BRANCHED基因,分别为BRC1和BRC2,它们编码TCP(Teosinte branched/cycloidea/proliferating cell factor)转录因子。在拟南芥中,AtBRC1和 AtBRC2都负调控分枝[10,15]。然而,研究表明AtBRC1在腋芽发育中似乎比AtBRC2发挥更关键的作用[16]。目前在异源四倍体油菜中至今尚未有关于BRC1的研究报道,因此对该基因展开功能研究有助于揭示油菜中分枝形成的调控机理。
CRISPR/Cas9基因组编辑技术是目前在农作物遗传改良中应用最广泛的基因编辑技术。本研究欲利用CRISPR/Cas9技术在油菜中靶向突变BRC1基因,以期获得油菜BRC1突变体,为阐释BRC1的基因功能奠定基础,并为株型育种提供宝贵的种质资源。
1 材料与方法
1.1 材料
1.1.1 植物材料 本实验所用的遗传转化受体材料为半冬性甘蓝型油菜品系甲9707(J9707),由华中农业大学周永明教授提供。遗传转化获得的再生苗、繁殖的转化苗后代和小区试验在华中农业大学转基因基地试验田中进行。
1.1.2 菌株与载体 构建CRISPR/Cas9载体所用到的双元载体pYLCRISPR/Cas9P35S-H(潮霉素抗性)和CRISPR/sgRNA载体pYLsgRNA-AtU3d/LacZ、pYLsgRNA-AtU3b、pYLsgRNA-AtU6-1 和pYLsgRNA-AtU6b-29(氨苄抗性)由华南农业大学刘耀光课题组提供;用于油菜转化的农杆菌菌株GV3101由本实验室保存;大肠杆菌菌株DH5α购自北京全式金生物技术有限公司。
1.2 方法
1.2.1 编辑载体构建与转化 靶点设计采用CRISPR-P 2.0(http://crispr.hzau.edu.cn/CRISPR2/)在线网站设计,靶点引物列于附表1。具体的构建流程参照Ma等[17]所描述的方法构建CRISPR/Cas9编辑载体,得到的编辑载体经过测序验证后转入农杆菌感受态,再通过油菜下胚轴遗传转化方法转化油菜受体材料J9707。具体操作方法参考杨阳[18]的方法。
1.2.2 突变体的转基因阳性鉴定和编辑鉴定 对获得再生苗采用CTAB法提取植物基因组DNA,采用载体上的引物PB-L/R(表1)进行转基因的阳性鉴定。对得到的阳性单株按照Liu等[19]的方法进行高通量测序,测序数据使用在线网站Hi-Tom(http://www.hi-tom.net/hi-tom/)分析每个单株的具体突变形式。
2 结果
2.1 甘蓝型油菜BnaBRC1的基因克隆及序列分析
根据拟南芥ATBRC1(AT3G18550)序列信息,在甘蓝型油菜基因组Darmor中blast比对到5个同源基因拷贝,分别为BnaC01g34090D、BnaA01g26700D、BnaC05g51920D、BnaCnng23770D和BnaA03g34820。设计引物(表1)对这5个拷贝进行基因克隆,得到这5个拷贝的基因序列。进一步构建BnaBRC1同源基因系统进化树发现BnaC01g34090D和BnaA01g26700D相似性最高,在核苷酸和蛋白序列上相似性分别为97.9%和96.7%(图1)。在受体材料J9707中进行同源基因DNA克隆和cDNA克隆,结果发现这5个同源基因都具有4个外显子。通过MEME进行保守结构域分析,结果发现BnaBRC1中存在2个保守的motif,属于glutamic acid-cysteine-glutamic acid motif(ECE), 功能未知,位于保守TCP domain中(图2-A),TCP domain位于第8-164个氨基酸之间(图2-B)。
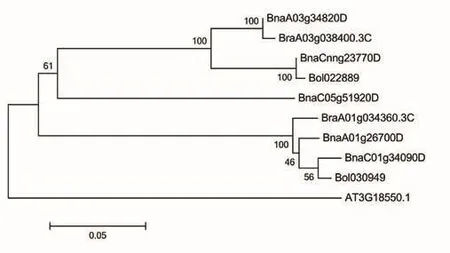
图1 BnaBRC1蛋白系统进化分析Fig.1 Phylogenetic analysis of BnaBRC1 protein

图2 BnaBRC1基因的Motif分析及保守结构域分析Fig.2 Motif analysis and conserved domain structures of BnaBRC1 gene
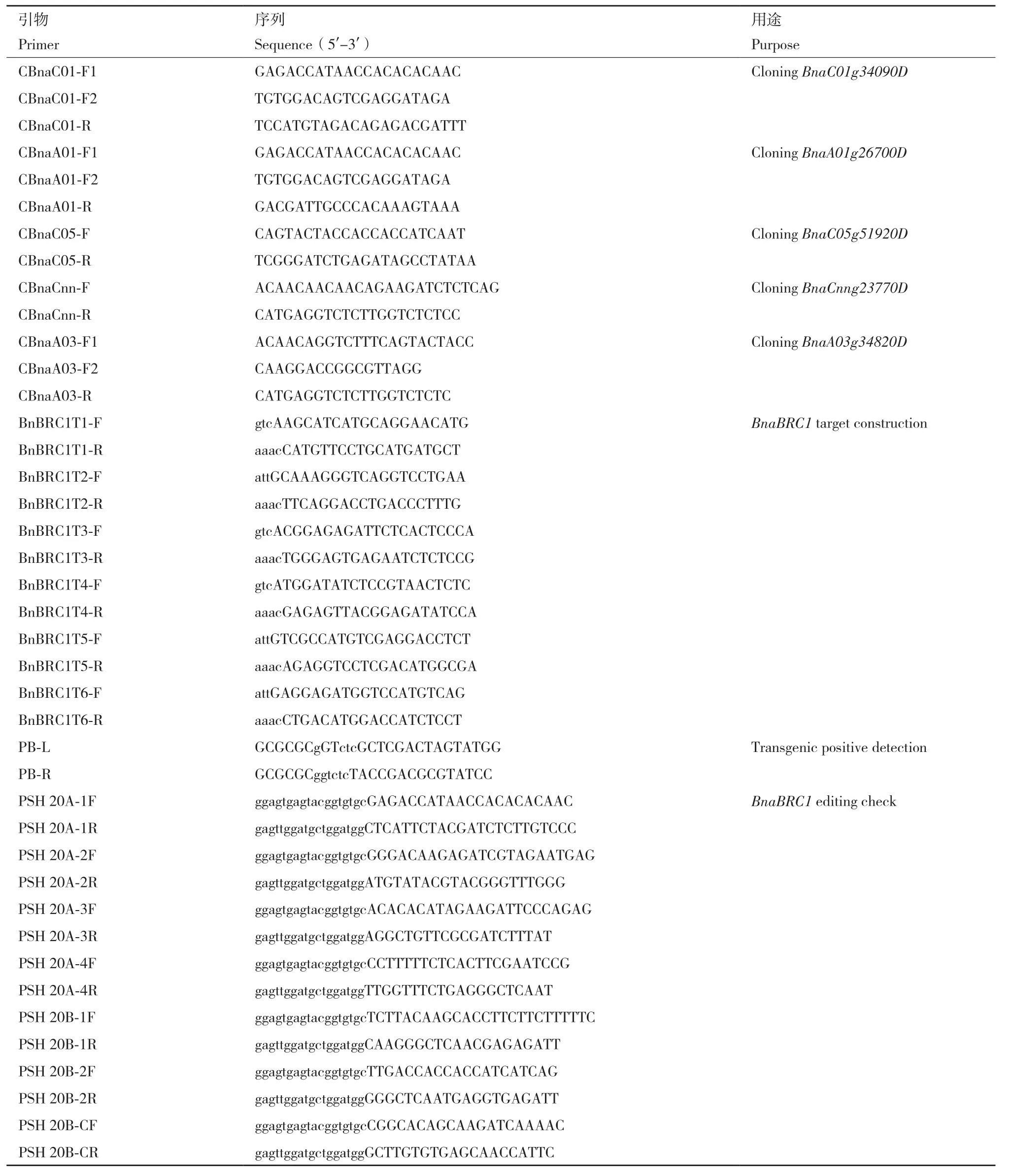
表1 本研究中使用的引物Table 1 Primers used in this study
2.2 甘蓝型油菜BnaBRC1的组织表达特异性分析
利用网上公开的甘蓝型油菜中双11的转录组数据库(http://yanglab.hzau.edu.cn/BnTIR/expression_show)对BnaBRC1基因在不同组织中表达水平进行分析,发现BnaA01g26700D和BnaC01g34090D的表达水平相对其他拷贝较高;且BnaBRC1基因的不同拷贝主要在花粉、花蕾和16 d-26 d的种子中有相对较高的表达,在其他组织中表达量很低或者不表达(图3)。

图3 BnaBRC1基因5个同源拷贝在油菜的不同组织中的转录丰度Fig.3 Transcript abundances of five BnaBRC1 copies in different tissues of B.napus
2.3 甘蓝型油菜BnaBRC1多靶点的CRISPR/Cas9载体构建
在设计sgRNA靶点时,根据BnaBRC1基因的5个同源拷贝之间序列相似性分成两组,设计两个CRISPR/Cas9载体,分别靶向不同的同源拷贝(图4-a)。第一组设计 3个 sgRNA 盒(S1-S3),分别靶 向BnaC01g34090D、BnaA01g26700D和BnaC-05g51920D,其中BnaC01g34090D命名为BnaC01.BRC1a(CC1),BnaA01g26700D 命 名 为 BnaA01.BRC1a(AA1),BnaC05g51920D 命 名 为 BnaC05.BRC1a(CC2)。该载体命名为P35s:Cas9-BnBRC1a(SBRC1a)( 图4-b)。 第 二 组 也 设 计 3个 sgRNA盒(S4-S6), 分 别靶向 BnaC05g51920D、BnaCnng23770D和BnaA03g34820,其中BnaCnng23770D命 名 为 BnaCnn.BRC1b(CC3),BnaA03g34820命名为BnaA03.BRC1b(AA2),该载体命名为P35s:Cas9-BnBRC1b(SBRC1b)(图4-b)。其中 sgRNA 靶向基因的位置信息和与基因序列的匹配程度如图4-a所示。这两个载体的sgRNA分别用拟南芥启动子pU3d、pU3b和pU6-1启动表达(图4-b)。

图4 BnaBRC1基因结构与靶点序列及双元质粒载体示意图Fig.4 BnaBRC1 gene structure with target sequences and schematics of binary plasmid vectors
2.4 BnaBRC1突变体的创建
将构建好的CRISPR/Cas9编辑载体通过农杆菌介导的下胚轴遗传转化法转化甘蓝型油菜受体J9707。其中SBRC1a载体共获得100株再生苗,SBRC1b载体共获得90株再生苗。利用载体引物PB-L/R(表1)对所有再生苗进行转基因阳性检查,结果SBRC1a和SBRC1b载体分别获得80和70株转基因阳性单株,转基因阳性效率分别为80%和78%。
为进一步确定转基因阳性单株的基因型,采用Hi-TOM两轮PCR产物高通量建库测序的方法[19],分析每个单株的具体突变形式。结果SBRC1a和SBRC1b转化载体分别检测到11株和12株编辑单株,编辑效率分为13.8%和17.1%,其中SgRNA1(S1)和SgRNA5(S5)处没有发生编辑。SBRC1a中BnaC01g34090D和BnaA01g26700D同时发生编辑的单株有11株,而BnaC01g34090D、BnaA01g26700D和BnaC05g51920D三个拷贝都发生编辑的单株仅有2株;SBRC1b中BnaCnng23770D和BnaA03g34820同时发生编辑的单株有11株,BnaCnng23770D单独发生编辑的单株有1株,而没有BnaCnng23770D、BnaA03g34820和BnaC05g51920D三个拷贝都发生编辑的单株。本研究中个别的靶点靶向不同同源拷贝的编辑效率有较大的差异,其中S2、S3靶点针对BnaC01g34090D和BnaA01g26700D的编辑效率明显高于BnaC05g51920D,其余的靶点则没有明显的拷贝偏好性。
根据测序结果,发现CRISPR/Cas9介导的基因编辑在甘蓝型油菜中引入的突变类型大多数为单个碱基的插入或缺失(图5),少数情况为大片段缺失。

图5 BnaBRC1突变体株系在T0代和T1代中S2和S3靶位点突变基因型Fig.5 Mutated alleles at the S2 and S3 target sites of BnaBRC1 mutants from T0 to T1 generation
2.5 BnaBRC1突变体表现为多主轴表型
由于疫情原因,只有部分T0代突变体材料得到自交种,其中在SBRC1a转化载体中只获得了BnaBRC1基因C01和A01同源拷贝同时突变的株系。而SBRC1b编辑转化材料没有收到种子,由于组织培养出来的再生苗表型受到很多因素的影响,故没有收集T0代突变体的表型。对获得的BnaC01g34090D和BnaA01g26700D拷贝突变体进行基因型鉴定,结果表明突变基因型可以从T0代稳定遗传给T1代。在T1代对aa1AA2cc1CC2CC3突变体进行表型考察,发现相较于野生型材料,突变体平均株高只有93.7cm,为野生型材料的53.5%(图6-A);苗期叶片数为野生型材料的3倍(突变体平均叶片数为33,野生型材料平均叶片数为11)(图6-A);相对于野生型材料只有一个主轴,突变体有2-4个主轴(图6-C),突变体的多分枝表型在植株成熟期依旧保持和苗期一样的数目。以上结果说明,BnaBRC1同源基因中BnaC01g34090D和BnaA01g26700D拷贝对分枝数目的调控具有比较重要的作用。

图6 BnaBRC1突变体的表型观察Fig.6 Phenotypes of BnaBRC1 mutants
2.6 BnaBRC1编辑材料的脱靶检测
为了检测本研究中6个靶位点的脱靶情况,对S1-S6所有靶点进行脱靶检测。利用CRISPR-P在线软件预测每个靶点可能存在的小于5个碱基错配的潜在脱靶位点。S1-S6分别对应有9、14、11、18、18个脱靶位点,我们对这70个潜在脱靶位点设计PCR扩增引物(附表1),分别对72、72、72、54、54株编辑单株进行PCR扩增,用PCR产物通过高通量测序。分析测序结果发现这70个潜在脱靶位点都没有基因编辑发生(表2),说明这些靶点特异性较高,均没有脱靶。

表2 在T0代突变体中检测每个sgRNA的脱靶活性Table 2 Detection of potential off-target effects for each sgRNA target site in T0 mutated plants
3 讨论
获得特定基因的突变体并对其进行表型分析是鉴定基因功能非常有效的一种方式。而采用物理和化学诱变等传统的方法创建突变体,不仅耗时费力、缺乏目标性,而且在突变体基因组中还存在大量的随机突变[20-21]。油菜为异源四倍体油料作物,大多数基因都具有功能相似的多个同源拷贝。因此,要获得具有表型变异的突变体,往往需要将这些功能冗余的拷贝同时突变才可能实现,进一步增加了利用传统诱变方法鉴定油菜基因功能的难度。近年来发展起来的CRISPR/Cas9基因编辑技术能靶向特定的基因进行定点突变,很好的克服了传统诱变方法的缺点,而且该技术易于掌握和操作,为油菜等多倍体作物的突变体创建和基因功能研究提供了强有力的工具[22-23]。
在本研究中,共设计了6个sgRNA表达盒靶向BnaBRC1基因,在T0代共获得23株突变体,表明该方法的有效性。进一步分析发现各个靶点的编辑效率存在较大差异。这可能与Cas9和sgRNA的表达水平、靶点GC含量、靶向背景和靶向sgRNA的二级结构等因素相关[22-24]。
在创建的BnaBRC1突变体中,我们观察到BnaC01g34090D和BnaA01g26700D双基因纯合突变体具有多分枝的表型,说明BnaBRC1基因的BnaC01g34090D和BnaA01g26700D同源拷贝对分枝数目的调控具有比较重要的作用。由于本研究中未能获得该基因其它拷贝的纯合突变体,因此这些拷贝是否具有调控分支的功能仍然未知。在后续研究中需要通过分离或聚合得到更多同源拷贝的单基因和多基因突变体,并对其进行表型考察以明确该基因各个同源拷贝的功能。此外,需要对获得的BnaBRC1多主轴突变体进行综合农艺性状考察,明确该多主轴表型是否具有增产效益。
本研究验证了BRC1的基因功能在不同物种中具有保守性,都参与腋芽的发育,从而调控分枝形成。研究表明BRC1的表达受到多种激素包括细胞分裂素、独角金内酯及赤霉素的调控[25-27]。而油菜中BRC1的表达主要由哪些激素调控还未知,对BnaBRC1的调控机理仍需进一步的研究。
4 结论
本研究利用CRISPR/Cas9技术对甘蓝型油菜中的BnaBRC1进行了定点突变。其中,BnaC-01g34090D和BnaA01g26700D双拷贝纯合突变体表现出明显的多分枝表型,表明该基因的这两个拷贝参与油菜中的分枝和株型调控。本研究为后续研究油菜的株型调控分子机理提供了重要研究材料。
文章所有附表数据请到本刊网站下载(http://biotech.aiijournal.com)。
