1个花生EMS突变体的表型评价及荚果大小相关性状分析
2022-06-04吕正昊史晓龙周东英刘喜波康树立赵新华王传堂于海秋
吕正昊,史晓龙,周东英,钟 超,刘喜波,张 鹤,康树立,赵新华,王传堂,于海秋
(1.沈阳农业大学农学院,沈阳 110161;2.山东省花生研究所,青岛 266000)
花生(Arachis hypogaea
L.)是一种重要的油料作物和经济作物,能为人类提供蛋白质、必需脂肪酸等营养物质,在我国以及世界范围内被广泛种植。近年来,我国对食用油的需求量不断增加,花生已成为我国植物油的第二大来源。辽宁是我国新兴的花生产区,常年种植30 万hm以上,但多种植于土壤贫瘠的地块,提高花生的产量潜力对培育高产、优质的花生新品种及保障食用油供给具有重要意义。产量是花生最重要的育种目标之一,花生的产量与单株荚果数、荚果重和种子重成正比。花生荚果由果壳和种子两部分组成,在多数情况下,花生荚果大小和种子大小为正相关,种子的生长受到果壳大小的制约,较大的果壳能给种子提供更充分的发育空间。果壳由子房壁发育而来,分为外果皮、中果皮和内果皮。在果针入土后,果壳率先发育,并作为保护和感知器官使种子免受外界胁迫,确保其正常发育。其中花生的内果皮是供应种子发育的光合产物临时贮存场所,而种子重及饱满与否,取决于光合产物的可用性以及向种子中的转运。总而言之,花生的果壳对种子的形成及产量有直接影响。目前已有许多关于果针、荚果和种子发育的研究,但对于花生果壳形成及发育的研究还相对较少。
利用诱变技术创制新种质,是加速花生育种的重要手段。EMS 诱变具有操作方便、突变率较高、损伤较小等优点,被广泛应用到花生突变体创制中。课题组前期从Tifrunner的诱变后代中筛选出一个大果突变体,命名为ps
。本研究对突变体及其野生型亲本进行调查研究,以期探明突变体的表型变异及花生荚果大小与种子大小的关系,为进一步研究花生荚果大小的调控机制奠定基础。1 材料与方法
1.1 材料
供试材料的野生型为美国栽培花生Tifrunner(花生四倍体参考基因组),为匍匐型花生。突变体为山东省花生研究所分子育种课题组于2018年诱变而来,经M单株选择后,M和M被种植于2020年和2021年,突变体命名为ps
。1.2 方法
称取EMS 45g,用少量70%酒精溶解,再加pH 值7.0 磷酸缓冲液(0.1mol·L)5L,配成浓度为0.9%的试剂。按每粒种子2mL用量,种子经EMS预浸泡6h,摇动浸泡4h。处理完成后,用流水冲洗种子约4h。
突变体和野生型材料在2020、2021 年均种植于山东省花生研究所莱西试验站。2020 年4 月30 日播种,一垄双行,行长1m,株距20cm;2021年5月1日播种,一垄单行,行长4m,株距30cm。田间管理同常规。田间种植均采用完全随机设计,每个材料3 次重复。盆栽试验于2021 年4 月8 日在室内播种,出苗后待温度适宜,移到室外。盆深21cm,直径33cm,每个材料播种10 盆,每盆2 粒,即每个材料20 个单株,用于调查开花特性。单株开花数=总开花数/单株数。果针标记于2021年大田进行,在盛花期用彩线进行标记,标记后覆土。
突变体的表型鉴定,植株性状测量参考《花生种质资源描述规范和数据标准》,在花生成熟期每个材料随机选取5 株有代表性植株,测定主茎高、第一对侧枝长、总分枝数、结果枝数、结实范围、双仁果数,收获晾干后各材料分别取5个有代表性的荚果,3次重复,测定荚果长、荚果宽、荚果重、种子长、种子宽、种子重。开花数量统计是从植株第1朵花出现开始,每天记录所有花生每株当天的开花量,连续观测60d。
石蜡切片及甲苯胺蓝染色试验:纵切面是将花生果壳沿腹缝线切开,横切面是在此基础上在果腰位置水平切开。随后将样品立即固定于FAA 中,无水乙醇梯度脱水(75%、85%、90%、95%和2×100%),100%二甲苯透明,石蜡包埋。采用RM2016显微切割仪切割6μm蜡带,贴于载玻片上。依次将切片放入二甲苯Ⅰ(20min)、二甲苯Ⅱ(20min)、无水乙醇Ⅰ(5min)、无水乙醇Ⅱ(5min)和75%酒精(5min),自来水洗;甲苯胺蓝染色:组织切片入染液约2~5min 后水洗,镜检,根据组织着色深浅进行适当的分化或者不分化,自来水洗后,将切片置于烤箱内烤干;透明封片:切片入干净的二甲苯透明10min,中性树胶封片;显微镜镜检,图像采集分析。
正置光学显微镜及成像系统分别使用日本尼康公司NIKON ECLIPSE E100和NIKON DS−U3。图像分析使用CaseViewer及ImageJ软件。细胞数量按公式计算,细胞面积=单位面积内细胞总面积/单位面积内细胞个数。试验数据采用Microsoft Excel 2013进行统计,采用GraphPad Prism8及OriginPro 2021b 软件进行方差分析、相关分析及图片绘制。
2 结果与分析
2.1 突变体的表型鉴定
通过2020~2021 连续两年的种植,发现与Tifrunner(WT)相比,突变体(ps
)仍表现出匍匐型的特点,叶型、叶色无明显变化(图1)。同时,与WT相比,ps
的主茎高、第一对侧枝长、结果枝数及结实范围均无显著差异(表1),而ps
的总分枝数显著减少(p
<0.05),两年分别减少47.3%和44.4%。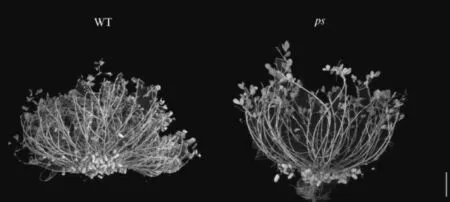
图1 ps和WT的表型特征Figure1 Phenotypic characterization of pw and WT lines
表1 和WT的株型相关性状
Table1 Plant type related traits of and WT lines
注:检验,*表示<0.05。
Note:−test;*<0.05.
ps
表现出明显的表型变异。ps
的荚果长、荚果宽和荚果重均极显著大于WT(p
<0.01)(图2A,B,C),其中荚果长在两年分别增加11.1%和18.2%;荚果宽分别增加15.4%和21.4%。在种子大小方面,ps
在2020年的种子长、种子宽和种子重均显著大于WT(p
<0.05),这一差异在2021 年达极显著(p
<0.01)(图2D,E,F)。种子长在两年分别增加30.8%和20.0%;种子宽分别增加42.9%和11.1%。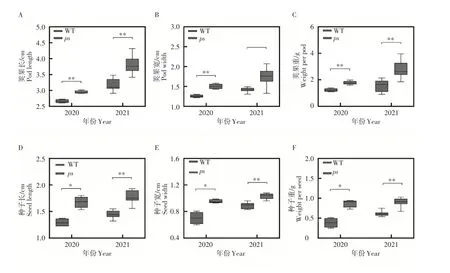
图2 ps和WT的荚果及种子的表型特征Figure2 Phenotypic characterization of pod and seed in the ps and WT lines
2.2 单株开花动态
由图3 可知,ps
在5 月31 日出现第一朵花,于6 月3日进入始花期。WT 在6 月16 日出现第1 朵花,于6 月17 日进入始花期。ps
于6 月14 日开始进入盛花期,而WT 于7 月4 日开始进入盛花期。相比于WT,ps
的始花期提前15d,而盛花期提前19d。同时,在调查期内,ps
的单日最高开花量高于WT。这些结果表明ps
的花期较WT提前,且植株活力略有提高。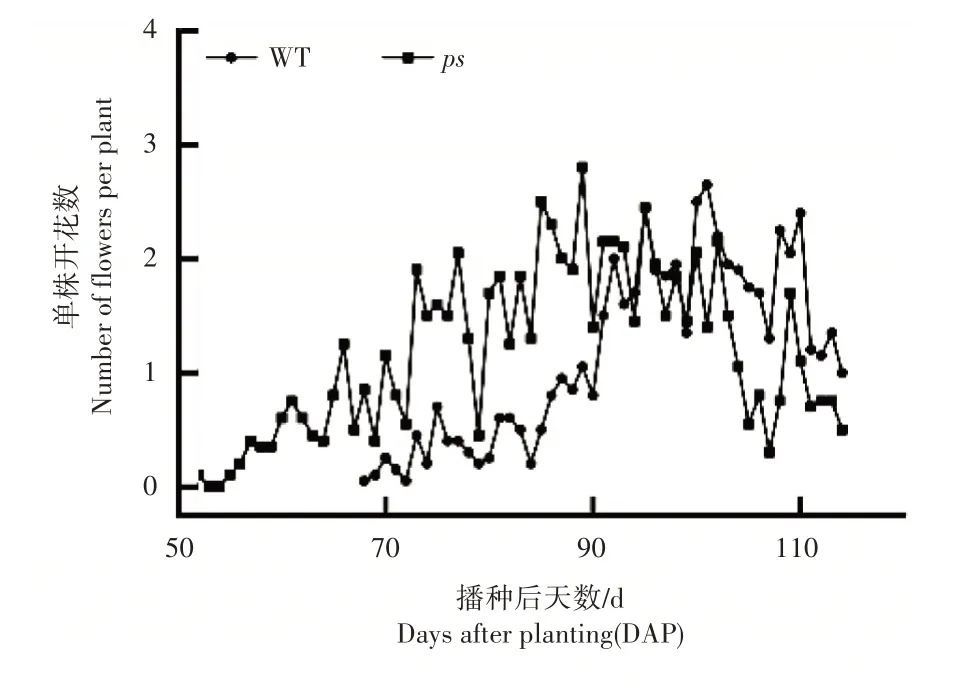
图3 ps和WT的单株单日开花量Figure3 The number of ps and WT flowering per day
2.3 主要农艺性状的相关性分析
通过对ps
及WT 的主要农艺性状进行相关性分析(表2),发现主茎高与总分枝数(r
=−0.73,p
<0.01)极显著负相关,与双仁果数(r
=−0.67,p
<0.05)存在显著负相关关系;第一对侧枝长与百果重(r
=0.77,p
<0.01)极显著正相关,与百仁重(r
=0.68,p
<0.05)显著正相关;总分枝数与结果枝数(r
=0.72,p
<0.01)及双仁果数极显著正相关(r
=0.81,p
<0.01);结果枝数与双仁果数间存在显著正相关关系(r
=0.61,p
<0.05);百果重与百仁重极显著正相关(r
=0.87,p
<0.01)。表2 主要农艺性状的相关性分析
Table2 Correlation analysis of main agronomic traits
2.4 荚果大小与种子大小的相关性分析
由图4可知,花生的荚果长与种子长存在极显著相关性(p
<0.0001),相关系数为0.9162(图4A),由此可见,随着荚果长度增加,种子长度也随之增加。荚果宽与种子宽也存在极显著的线性关系(p
<0.0001),相关系数为0.9154(图4B)。花生荚果的长宽即为果壳的长宽,这一结果表明种子的大小与果壳大小存在正相关关系。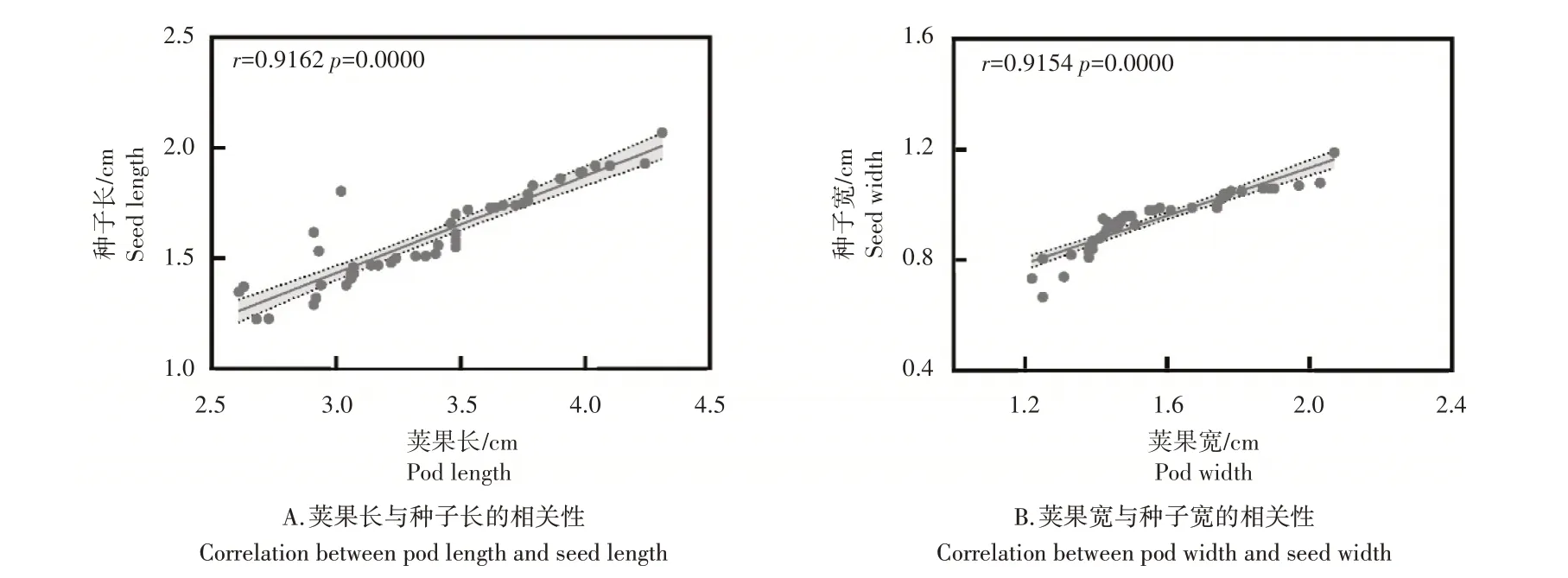
图4 荚果长、宽与种子长、宽的相关性Figure4 The correlation between pod length and seed length and the correlation between pod width and seed width
2.5 ps和WT的果型与粒型分析
荚果、种子的长宽比直接决定了果型和粒型,对产量有直接影响。由图5 可知,相比于WT,ps
的长宽比并无显著变化,而种子的长宽比显著增加(p
<0.01)。这说明ps
的果型与WT 一致,在突变后并未发生变异,但粒型显著变长。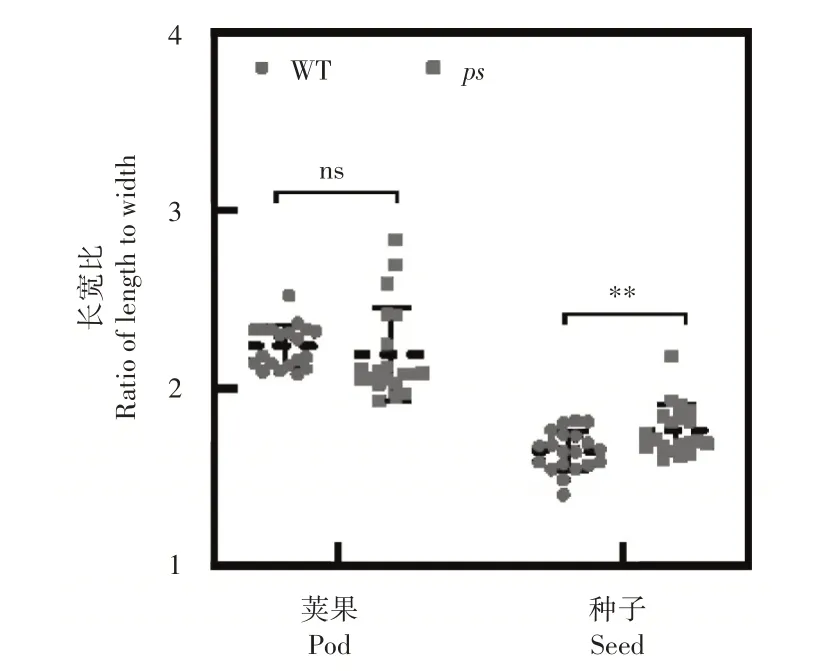
图5 ps和WT的荚果、种子长宽比Figure5 Comparative analysis of PL/SL and PW/SW in ps and WT
2.6 ps和WT的细胞学分析
为了进一步分析ps
荚果变大的原因,在果针入土后20d 对ps
和WT 的果壳进行了细胞学分析。细胞数量及细胞面积的统计方法图6A 和图6D)。结果表明,在纵切面上,与WT 相比,ps
的细胞数量极显著增加(p
<0.01),增幅达到18.42%(图6B),细胞面积与WT无显著差异(图6C);在横切面上,ps
的细胞数量相较于WT 显著增加了(p
<0.05)10.32%,(图6E),而细胞面积与WT无显著差异(图6F)。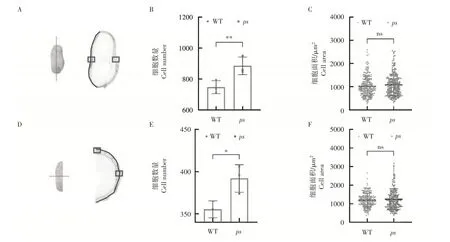
图6 ps和WT的细胞学分析Figure6 Cytological analysis of ps and WT
3 讨论与结论
在作物育种中,新品种选育的基础是遗传变异。而栽培花生的遗传基础狭窄,遗传变异率较低,制约了花生新品种的选育。花生品种改良应通过创制优异花生新种质,进而获得理想的性状。诱变是拓宽作物遗传变异以克服遗传基础狭窄限制的重要途径之一,通过诱变获得优异种质,是花生新品种选育的有效途径。EMS诱变被广泛应用于花生并成功获得株型、品质、种皮颜色及荚果发育突变体。在本试验中,通过EMS 诱变获得了一个分枝数减少、花期提前且荚果变大的突变系ps
,为选育适宜本地区机械化种植的花生新品种提供了优异种质资源。相关性分析表明,总分枝数与结果枝数及双仁果数存在显著正相关关系(表2)。ps
的分枝数相比于WT显著减少(表1),然而结果枝数却无显著变化,这说明ps
所减少的为无效分枝,促进有效分枝的养分积累。花生的株型能间接影响产量,而分枝数是花生株型构成的重要因素之一,减少无效分枝,有利于有机物向有效分枝的转运从而提高产量。此外,ps
进入始花期及盛花期的时间要比WT提前,下针相对较早。研究表明,花生的开花期越早,形成果针及结荚率就越高。荚果大小直接影响花生的产量,增加荚果大小是提高花生产量潜力的有效途径。在本试验中,ps
的荚果及种子大小均显著大于WT(图2),且相关性分析表明,荚果长、宽与种子长、宽存在着极显著的线性关系(图4),说明ps
的种子大小随着荚果的变大而增大。然而,更大的器官不一定产生更大的种子,种子和器官的大小具有不同的调控途径。在本试验中,ps
的果型并未改变,但粒型显著变长(图5),说明其果壳和种子的发育具有不同的调控途径。正因如此,针对ps
荚果大小研究时应重点关注果壳的发育。植物果实的大小主要由细胞数量和大小决定,细胞学分析表明,造成ps
荚果长和宽增加的主要原因是细胞数量的增加(图6)。综上,本试验为研究花生荚果大小及构建定位群体提供了优异的突变体材料,并阐明了针对突变体ps
的果型研究应从果壳发育入手,为进一步研究其分子机制奠定基础。