确山黑猪HOXA5基因组织表达及生物信息学分析
2022-06-01段栋栋李聪李梦雨李欣王克君李秀领韩雪蕾乔瑞敏李新建
段栋栋, 李聪, 李梦雨, 李欣, 王克君, 李秀领, 韩雪蕾, 乔瑞敏, 李新建
(河南农业大学动物科技学院,河南 郑州 450046)
同源盒(homeobox, HOX)家族是高度保守的调节基因超家族,是与细胞发育相关的重要转录因子,广泛存在于高等真核生物体内,在调控细胞增殖、分化、迁移和凋亡等方面发挥着重要作用[1]。BRIDGES等[2]最早于1923年在果蝇体内发现并报道了HOX基因。在脊椎动物中,根据HOX基因在染色体上的位置将其分为HOXA、HOXB、HOXC和HOXD这4个基因簇,共计39个基因[3-4]。HOX基因不仅与伤口修复等生理功能有关[5],还与急性白血病[6]和癌症[7]等一系列疾病有关,而且在脂肪组织代谢中发挥着重要的调控作用[8]。KALYPSO等[9]研究认为,HOX基因对脂肪组织发育与重塑具有重要的调节作用。GESTA等[10]通过分析人类脂肪组织的特异性差异发现,HOXA5基因的表达水平与肥胖程度和脂肪分布有关。CANTILE等[11]在研究人类脂肪组织中发现,HOXA1、HOXA4和HOXC4基因可能直接参与白色和棕色脂肪组织的分化。PARRILLO等[12]研究发现,在高脂诱导肥胖小鼠的脂肪组织中HOXA5基因发生超甲基化导致其表达水平降低,且在干扰HOXA5基因后脂肪细胞分化被抑制。YAMAMOTO等[13]研究发现,HOXA5基因在小鼠棕脂和附睾白脂表达水平较高,而在皮下白脂中表达量较低,且与脂质分解密切相关。SINGH等[14]发现棕色脂肪细胞的成脂与HOXA5基因紧密相关。脂肪沉积是脂肪组织发生和脂肪细胞增殖分化的一系列复杂的生物学过程,根据脂肪组织生成部位的不同可将其分为皮下脂肪、肌内脂肪和内脏脂肪,而皮下脂肪和背最长肌中的肌内脂肪是影响猪肉品质的重要因素[15]。确山黑猪是河南省优良的地方猪品种,具有繁殖力高、脂肪沉积能力强以及肉质优良等特性,属于脂肪型猪种[16],是研究HOXA5基因表达模式的理想试验动物,但是目前关于HOXA5基因对确山黑猪脂肪代谢影响的研究还未见报道。大白猪是世界上分布最广的瘦肉型猪种之一,与确山黑猪相比在胴体脂肪比例方面有所不同。本研究明确了确山黑猪HOXA5基因在不同组织(心脏、肝脏、脾脏、肺脏、肾脏、皮下脂肪、腹部脂肪、背最长肌)中的表达差异,对比了HOXA5基因在确山黑猪与大白猪的皮下脂肪和背最长肌表达效果,并对确山黑猪HOXA5蛋白生物学进行了分析,以期探究HOXA5基因对猪脂肪沉积的影响,为今后研究HOXA5基因调控猪脂肪沉积的分子机制提供理论基础。
1 材料与方法
1.1 样品与试剂
试验选用在相同饲养条件下约110 kg健康的确山黑猪和大白猪各3头。屠宰后,采集确山黑猪的心脏、肝脏、脾脏、肺脏、肾脏、皮下脂肪、腹部脂肪和背最长肌组织,以及大白猪的皮下脂肪和背最长肌组织,并迅速置于-196 ℃液氮中保存备用。操作过程均在河南农业大学机构动物护理和使用委员会(institutional animal care and use committee,IACUC)(编号11-0085)的批准下进行。
试剂:Trizol、PrimerScriptTMRT reagent Kit等常规试剂均购自TaKaRa公司;琼脂糖凝胶 DNA 回收试剂盒购自天根生化科技有限公司;大肠杆菌TOP10感受态细胞购自TransGen Biotech公司。
1.2 引物设计与合成
根据GenBank中猪HOXA5基因(NM_001195232.1)的mRNA序列信息,使用Primer Premier 5.0 软件设计HOXA5基因克隆引物及实时荧光定量PCR(real-time quantitative PCR,qRT-PCR)相关引物。HOXA5基因克隆引物序列:HOXA5-F1(5’-CGAGCCACAAATCAAGCA-3’)和HOXA5-R1(5’-ATTAGGGCAACGAGAACA-3’),预期产物长度为971 bp,其中包含HOXA5基因813 bp 编码区(coding sequence,CDS)序列。qRT-PCR 特异性引物序列:HOXA5-F2(5’-CAACATAGGCGGCCCTGAA-3’),HOXA5-R2(5’-CGGAGAGGCAAAGAGCATGT-3’),预期产物长度为143 bp;β-actin内参基因引物序列:β-actin-F(5’-AGAGCAAGAGAGGCATCCTG-3’),β-actin-R(5’-CACGCAGCTCGTTGTAGAAG-3’),预期产物长度为111 bp。以上引物均由生工生物工程(上海)股份有限公司合成。
1.3 HOXA5基因组织表达差异分析
采用Trizol法提取确山黑猪各组织和大白猪皮下脂肪和背最长肌的总RNA,使用质量分数1%的琼脂糖凝胶电泳检测总RNA的质量,NanoDrop One分光光度计检测总RNA浓度。使用反转录试剂盒Prime Script RT Reagent Kit对总RNA进行反转录。以确山黑猪8种组织(心脏、肝脏、脾脏、肺脏、肾脏、皮下脂肪、腹部脂肪和背最长肌)的cDNA为模板,β-actin为内参基因,设置3个生物学重复,每个生物学重复设置3个技术重复,检测HOXA5基因在确山黑猪不同组织的相对表达量;以确山黑猪和大白猪的皮下脂肪和背最长肌的cDNA为模板,检测HOXA5基因在确山黑猪和大白猪皮下脂肪和背最长肌的表达情况。qRT-PCR反应体系为10 μL:cDNA 1 μL,上、下游引物各0.3 μL,2×SYBR Premix Ex Taq Mix 5 μL,ddH2O补足10 μL。qRT-PCR反应程序:95 ℃,30 s;95 ℃,10 s;60 ℃,30 s;共40个循环;95 ℃,15 s;68 ℃,5 s;95 ℃ ,5 s。采用2-ΔΔCt法计算其相对表达量,然后采用SPSS 25.0软件进行单因素方差分析。
1.4 确山黑猪HOXA5基因克隆及测序
以确山黑猪皮下脂肪组织cDNA为模板进行PCR扩增。PCR反应体系为50 μL:2×Phanta Max Buffer 25 μL,dNTP Mix 1 μL,Phanta Max Super-Fidlity DNA 1 μL,HOXA5-F1和HOXA5-R1各2 μL,cDNA 2.5 μL,ddH2O 16.5 μL。PCR反应程序为:95 ℃预变性3 min;95 ℃变性15 s,55 ℃退火15 s,72 ℃延伸1 min,共35个循环;72 ℃延伸5 min。PCR扩增产物经质量分数1%的琼脂糖凝胶电泳,对目的条带进行纯化回收,并与pMD19-T载体连接、转化。挑取单克隆菌落,经菌液PCR检测鉴定正确后进行测序。
1.5 确山黑猪HOXA5基因生物信息学分析
运用NCBI数据库和DNA Star Meg Align 7.1.0软件将确山黑猪HOXA5基因CDS区序列与牛(NM_001077098.1)、绵羊(XM_015095103.2)、马(XM_001499578.5)、鸡(NM_001318419.1)、人(NM_019102.4)、小鼠(NM_010453.5)和黑猩猩(XM_519011.6)的CDS区序列进行同源性对比并采用Mega 7.0软件构建系统进化树;使用Expasy在线软件中的ProtParam和ProtScale分别对HOXA5蛋白的理化性质和亲/疏水性进行分析。利用DTU Health Tech在线软件中的TMHMM和SignaIP分别预测HOXA5蛋白的跨膜结构以及信号肽。使用InterPro在线软件分析HOXA5蛋白的结构域。使用PSPRT Ⅱ Prediction在线软件预测HOXA5蛋白亚细胞定位。使用SOPMA和SWISS-MODEL在线软件分别预测HOXA5蛋白的二级结构和三级结构。
2 结果与分析
2.1 HOXA5基因在确山黑猪各组织表达分析
利用qRT-PCR技术检测HOXA5基因在确山黑猪不同组织中的相对表达量。结果显示,HOXA5基因在确山黑猪各种组织中均有表达。其中,在肺脏中表达量最高,其次是肝脏和皮下脂肪,且显著高于其他组织(P<0.05)(图1)。

注:不同字母表达差异显著(P<0.05)。
2.2 HOXA5基因在不同猪种的皮下脂肪和背最长肌中表达分析
为了进一步探究HOXA5基因对脂肪沉积的影响,利用qRT-PCR技术检测并对比HOXA5基因在确山黑猪和大白猪的皮下脂肪和背最长肌中的表达情况。结果显示,在皮下脂肪中确山黑猪HOXA5基因表达水平显著高于大白猪(P<0.05),而在背最长肌中2个猪种的HOXA5基因表达水平差异不显著(P>0.05)(图2)。

注:*,差异显著(P<0.05);ns,差异不显著(P>0.05)。
2.3 确山黑猪HOXA5基因克隆及序列分析
以确山黑猪皮下脂肪组织cDNA为模板,经PCR扩增后,使用质量分数1%的琼脂糖凝胶电泳检测,得到了1条片段大小为971 bp的特异性条带(图3),与目的条带结果一致。经序列分析后发现,确山黑猪HOXA5基因的CDS区没有发生碱基突变,与NCBI中猪HOXA5基因的CDS区序列完全一致。该CDS区全长813 bp,编码270个氨基酸,起始密码子为ATG,终止密码子为TGA,A、T、G和C分别占22.51%、13.90%、29.77%和33.82%(图4)。

注:M,DL2000 DNA Marker;1,HOXA5基因PCR产物;2,阴性对照。

图4 确山黑猪HOXA5基因CDS区和编码氨基酸序列
2.4 确山黑猪HOXA5基因生物信息学分析
2.4.1 确山黑猪HOXA5基因序列同源性对比及进化树分析 通过DNA Star Meg Align进行同源性对比分析,结果显示确山黑猪HOXA5基因与人、鸡、牛、马、绵羊、小鼠和黑猩猩的同源性分别为97.4%、81.3%、99.1%、98.8%、99.0%、95.6%和97.8%,变异度分别为2.6%、17.4%、0.9%、1.2%、1.0%、4.6%和2.3%(图5)。系统进化树分析显示,确山黑猪HOXA5基因与牛、绵羊的亲缘关系最近,其次为马、人和黑猩猩,与鸡亲缘关系最远(图6),这表明HOXA5基因核苷酸序列在不同物种间的保守性较高。

图5 确山黑猪与其他已知物种的HOXA5基因核苷酸序列比对

注:比例尺代表该单位长度下序列间差异数值为0.020;节点上数字表示bootstrape值。
2.4.2 确山黑猪HOXA5蛋白理化性质分析 经Prot Param预测分析可知,确山黑猪HOXA5蛋白总原子质量为3 991,分子质量为29.28 kD,分子式为C1262H1929N391O399S10,理论等电点(pI)为9.42。HOXA5蛋白共编码20种氨基酸(表1)。其中,丝氨酸(Ser)含量最高,为14.4%;半胱氨酸(Cys)和色氨酸(Trp)最低,均为1.1%。HOXA5蛋白含强酸性氨基酸(Asp+Glu)23个,强碱性氨基酸(Arg+Lys)31个,总体偏碱性。经预测HOXA5蛋白不稳定系数为65.30,为不稳定蛋白。
表1 确山黑猪HOXA5蛋白的氨基酸组成

续表
2.4.3 确山黑猪HOXA5蛋白亲/疏水性分析 使用ProtScale软件对确山黑猪HOXA5蛋白进行亲/疏水性分析,结果如图7所示。多肽链的第230位氨基酸的疏水分值最大,为1.778;第249位和250位氨基酸的疏水分值最小,为-2.967。综合分析显示,确山黑猪HOXA5蛋白共有34个疏水性氨基酸,228个亲水性氨基酸,表明该蛋白属于亲水性蛋白。

图7 确山黑猪HOXA5蛋白亲/疏水性预测
2.4.4 确山黑猪HOXA5蛋白跨膜结构预测及蛋白亚细胞定位预测 通过TMHMM软件对确山黑猪HOXA5蛋白跨膜结构进行预测分析,结果显示HOXA5蛋白不包含跨膜结构(图8);通过Signa IP软件预测信号肽,结果显示HOXA5蛋白没有信号肽(图9)。上述结果表明确山黑猪HOXA5蛋白不属于跨膜蛋白和分泌蛋白。亚细胞定位结果显示,HOXA5蛋白定位在细胞核、细胞质和细胞骨架中,其比例分别为91.3%、4.3%和4.3%,表明该蛋白主要位于细胞核内。

图8 确山黑猪HOXA5蛋白跨膜结构预测
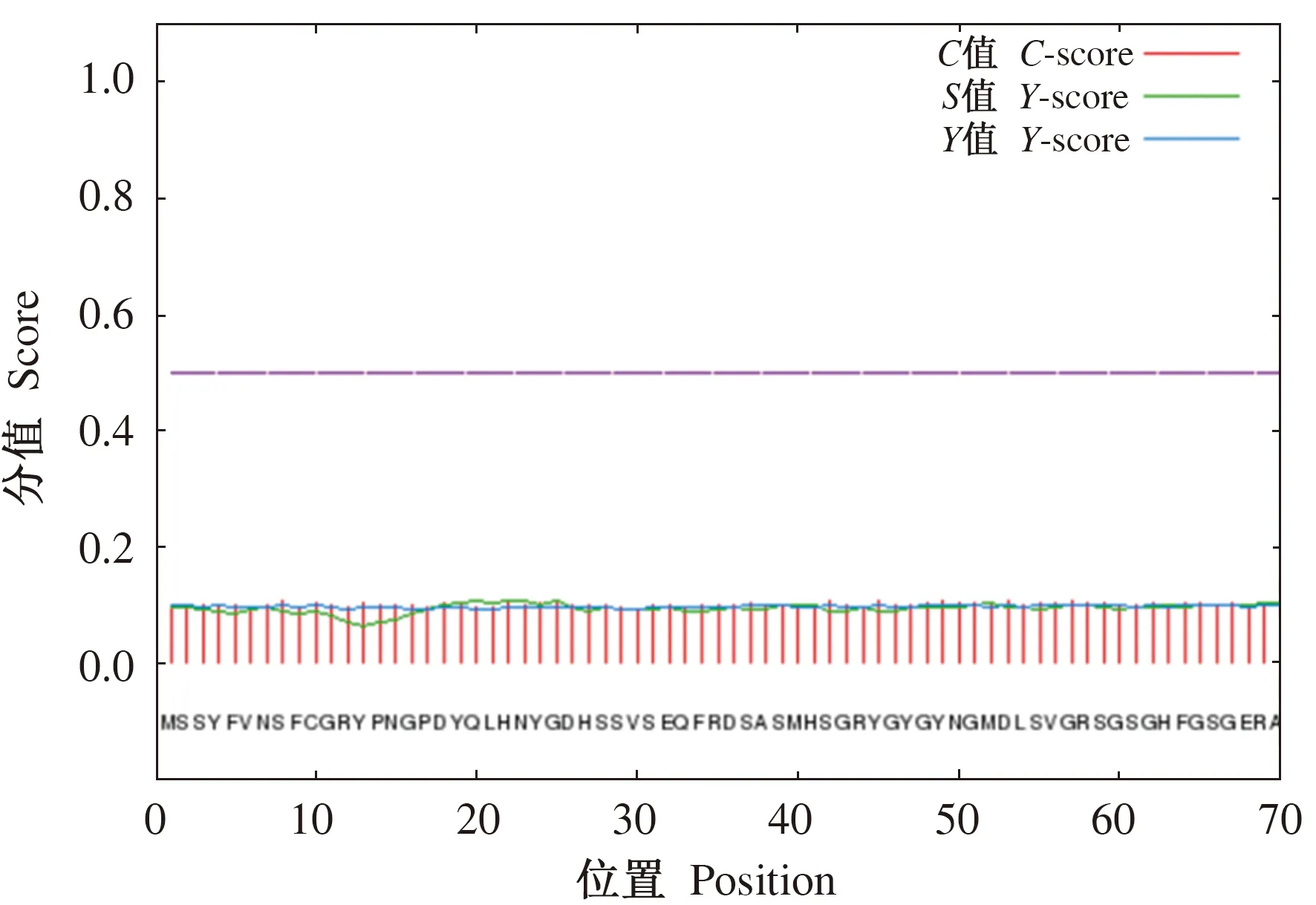
图9 确山黑猪HOXA5蛋白信号肽预测
2.4.5 确山黑猪HOXA5蛋白二级结构和三级结构预测分析 通过SOPMA软件对猪HOXA5蛋白二级结构预测,确山黑猪HOXA5蛋白含有4种二级结构,分别为α-螺旋(22.96%)、延伸链(11.48%)、β-转角(4.81%)和无规则卷曲(60.74%)(图10)。通过SWISS-MODEL软件对HOXA5蛋白三级结构进行预测,并将确山黑猪HOXA5蛋白的三级结构与牛、鸡、鼠和人的三级结构比较发现,在不同物种中HOXA5蛋白的三级结构无明显差异,且主要以无规则卷曲为主(图11)。
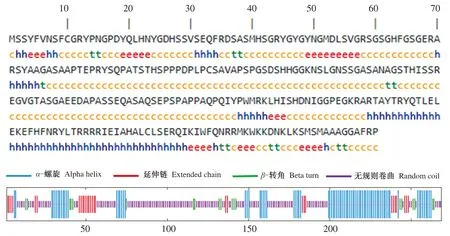
图10 确山黑猪HOXA5蛋白二级结构

图11 不同物种间HOXA5蛋白三级结构
3 结论与讨论
HOXA5基因在胚胎发育、器官形成和细胞命运等生物过程中起着重要的调节作用[17]。本研究结果显示,HOXA5基因在确山黑猪心脏、肝脏、脾脏、肺脏、肾脏、皮下脂肪、腹部脂肪和背最长肌8种组织中均有不同程度的表达,表明该基因在机体内广泛表达。其中,HOXA5基因在肺脏中表达量最高,且显著高于其他组织。HOXA5基因对肺脏的发育和成熟发挥重要的作用。MANDEVILLE等[18]研究表明,HOXA5基因的缺失对肺部后天发育产生严重的影响。KRUMLAUF等[19]研究表明,在小鼠出生后HOXA5基因在肺脏中保持高表达水平,并且HOXA5基因的缺失会导致小鼠呈现肺脏异常,肺泡分隔不足和肺不扩张等症状。此外有研究表明,HOXA5基因间接控制脂肪细胞中的信号因子调节脂肪代谢。GESTA等[10]研究表明,HOXA5基因的表达量与身体指数和腰臀比成正相关。CAO等[20]研究表明,HOXA5基因通过抑制 PKA/HSL通路促进脂肪细胞分化。FEI等[21]研究表明,HOXA5基因通过抑制小鼠白色脂肪细胞中的Akt/mTORC1信号通路来增加细胞凋亡。本研究中,HOXA5基因在肝脏、皮下脂肪及腹部脂肪等与脂肪代谢相关的组织中表达量较高。同时,为了进一步研究HOXA5基因在确山黑猪脂肪沉积中的调控功能,本研究检测了HOXA5基因在确山黑猪和大白猪皮下脂肪组织和背最长肌中的表达情况。结果显示,HOXA5基因在脂肪沉积量高的确山黑猪皮下脂肪组织中的表达量显著高于大白猪,推测HOXA5基因的高表达提高了确山黑猪脂肪沉积的能力。因此,HOXA5基因可作为调控猪脂肪沉积的重要候选基因,其具体作用及机制仍需进一步研究。
本研究通过分子克隆技术对确山黑猪HOXA5基因进行克隆测序,得到了确山黑猪HOXA5基因完整的CDS区序列,对其CDS区序列进行生物信息学分析。结果显示,确山黑猪HOXA5基因CDS区序列全长813 bp,编码270个氨基酸。研究发现,确山黑猪HOXA5基因CDS区序列与Gen Bank中公布的猪HOXA5基因(NM_001195232.1)序列完全一致。王艳[22]研究表明,梅山猪和大白猪HOXA5基因CDS区序列与Gen Bank中猪的序列也完全一致,由此推测HOXA5基因在不同猪品种中高度保守。本研究进一步对不同物种的HOXA5基因核苷酸序列进行同源性对比,发现确山黑猪HOXA5基因与牛和羊同源性最高,与该基因的系统进化树分析结果一致,这说明该基因在不同物种间具有高度保守性,在稳定物种的遗传性状方面起到重要的调控作用。
确山黑猪HOXA5蛋白生物信息学分析显示,确山黑猪HOXA5蛋白没有信号肽序列同时不含跨膜区,表明该蛋白属于不分泌蛋白且不参与信号传导。本研究还发现在HOXA5蛋白中有许多亲水性氨基酸,因而该蛋白也具有亲水性,为不稳定蛋白。蛋白质的功能与其空间结构有着密切的关系,HOXA5蛋白二级结构和三级结构预测分析,发现确山黑猪HOXA5蛋白主要由α-螺旋(22.96%)、延伸链(11.48%)、β-转角(4.81%)和无规则卷曲(60.74%)组成,为混合型蛋白,且该蛋白以无规则卷曲为主,与其不稳定蛋白的理化性质相符。本研究通过生物信息学分析,深入研究了HOXA5基因的结构和功能,不仅有利于揭示HOXA5基因在猪脂肪沉积方面的调控机制,同时也为确山黑猪品种的基因改良提供了理论基础。
