黑龙江省马铃薯疮痂病菌种群结构及PAI致病基因分析
2022-05-30张铉哲陈苏慧徐浩然任雪琦朱二龙
张铉哲 赵 雪 陈苏慧 徐浩然 任雪琦 朱二龙
(东北农业大学农学院,黑龙江哈尔滨 150030)
马铃薯疮痂病(potato common scab,PCS)是一种由致病链霉菌(spp)引起的土传病害,在各地马铃薯生产区几乎均有发生。该病害的特点是马铃薯块茎表皮层上有凸起、凹陷、网状或其他类型木栓状坏死病斑,颜色由褐色至黑色不等,其中病斑最深可造成约7 mm 的组织凹陷(刘淑娜,2019)。致病链霉菌除了能侵染马铃薯外,还能感染其他根菜类蔬菜,如胡萝卜、萝卜、根甜菜等,在肉质根表面形成疮痂病斑,降低蔬菜的品质和市销性,造成重大的经济损失(Wanner,2004;Hill &Lazarovits,2005;Qu et al.,2008)。近几年,马铃薯疮痂病的发生越来越严重,很大程度上影响了马铃薯的市销性。疮痂病是由1 种或几种链霉菌复合侵染引起的病害。链霉菌属放线菌,种类繁多,几乎所有的链霉菌都是土壤腐生菌,具有较强的抗逆性,产生的孢子在干燥条件下能长期存活。常见的致病链霉菌主要有3 种,即、以及(Loria et al.,1997)。是第1 个被报道的致病链霉菌,是全球引起疮痂病的主要病原菌(Lambert &Loria,1989;Corrêa et al.,2015),且被认为是分布最广的一种,其次是和(Lerat et al.,2009)。国内外对马铃薯疮痂病病原菌的分类进行了大量研究,不断有致病性链霉菌新菌株被发现,致病菌的组成趋于复杂化。
引起马铃薯疮痂病的致病链霉菌具有多样性,但致病性的基因和机制似乎在引起疮痂病的链霉菌中共享。致病菌的致病基因聚集在染色体一个大的区域中,且该区域具有在链霉菌种间水平移动的特点,可以水平转移到新的链霉菌物种中以产生致病性物种或菌株,从而起到致病岛(pathogenicity island,PAI)作用。该致病岛上参与毒素合成及调控的基因有、、、、、等(Healy et al.,2002;Kers et al.,2004;Joshi et al.,2010)。目前研究最多的基因是、和基因。
为了解黑龙江省马铃薯疮痂病菌的种类和链霉菌PAI 致病基因的特点,本试验从黑龙江省不同马铃薯种植地区采集具有典型马铃薯疮痂病症状的薯块,进行疮痂病菌的分离,通过病原菌的形态特征、生理生化特性结合链霉菌16S rDNA 序列对病原菌进行分类鉴定,并检测、和3种PAI 致病基因的组成,以期为追踪马铃薯疮痂病相关链霉菌的进化历史和传播资源,也为有效防治马铃薯疮痂病提供参考依据。
1 材料与方法
1.1 马铃薯疮痂病菌的采集与分离
2018—2020 年马铃薯收获期间,在黑龙江省牡丹江市、鸡西市、绥化市、鹤岗市、佳木斯市及哈尔滨市等马铃薯主产区采集疮痂病薯块。参考赵伟靖(2018)稀释分离法进行疮痂病菌的分离。在无菌条件下,选择白色粉状单菌落,划线纯化3 次,将纯化后的菌株编号保存,备用。
1.2 链霉菌的致病性测定
参考赵伟靖(2018)的方法利用萝卜幼苗法测定分离菌株的致病性。挑选已发芽长势一致的萝卜种子放入含有1%水琼脂的试管内,每支试管3 粒,接种10CFU·mL的马铃薯疮痂病菌菌悬液200 μL,以接种等量无菌水的处理作为空白对照。于光照培养箱中每天16 h 光照、8 h 黑暗培养7 d 后,统计萝卜幼苗的长度并计算病原菌对萝卜幼苗的抑制率。
抑制率=(1 -处理幼苗平均长度/对照幼苗平均长度)× 100%
1.3 致病链霉菌的鉴定
1.3.1 形态观察 挑取长势良好的致病链霉菌单一菌落,划线接种于ISP2 培养基(酵母提取物4 g,麦芽糖提取物10 g,葡萄糖4 g,琼脂粉20 g,蒸馏水1 000 mL)平板上,28 ℃黑暗条件下培养7 d,观察各测试菌株的色素产生情况。将菌株接种于高氏1 号培养基(可溶性淀粉20 g,NaCl 0.5 g,KHPO0.5 g,KNO1.0 g,MgSO·7HO 0.5 g,FeSO·7HO 0.01 g,琼脂粉20 g,蒸馏水1 000 mL)平板上,置于28 ℃培养箱中培养,14 d 后测试菌株充分生长,观察菌落形态及孢子链形态等特征。孢子链形态观察采用印片法(殷修鲁,2018)。
1.3.2 生物学特性测定 参考徐丽华等(2007)的方法对马铃薯疮痂病致病链霉菌的黑色素产生、硫化氢产生、纤维素利用、明胶液化、淀粉酶产生、碳源利用、氮源利用、敏感性等进行测定。
1.3.3 分子生物学鉴定 链霉菌总DNA 的提取采用细菌基因组DNA 提取试剂盒(北京天根生化科技有限公司)进行,用链霉菌16S rDNA 通用引物PA/PH 进行检测(李驰 等,2019)。PCR 产物送至生工生物工程(上海)股份有限公司进行双向测序,将序列拼接处理后的基因序列通过NCBI 进行Blast比对,构建系统发育树,对菌株进行同源性分析。
1.4 致病链霉菌的PAI 致病基因检测
通过PCR 的方法检测致病链霉菌的致病基因类型,检测菌株中是否含有、和致病基因,引物序列、PCR 扩增反应条件和体系参考李驰等(2019)方法,PCR 扩增结果用1%琼脂糖凝胶电泳检测,统计致病菌株的致病基因组成情况。
2 结果与分析
2.1 马铃薯疮痂病菌的分离纯化
2018—2020 年每年8—9 月马铃薯收获时节,采用5 点取样法从黑龙江省共6 个市8 个采样点的马铃薯主产区采集疮痂病薯块,得到154 份马铃薯疮痂病样品,进行菌株分离,挑选具有链霉菌形态特征的单菌落,纯化菌株,共分离得到链霉菌菌株312 株(表1)。
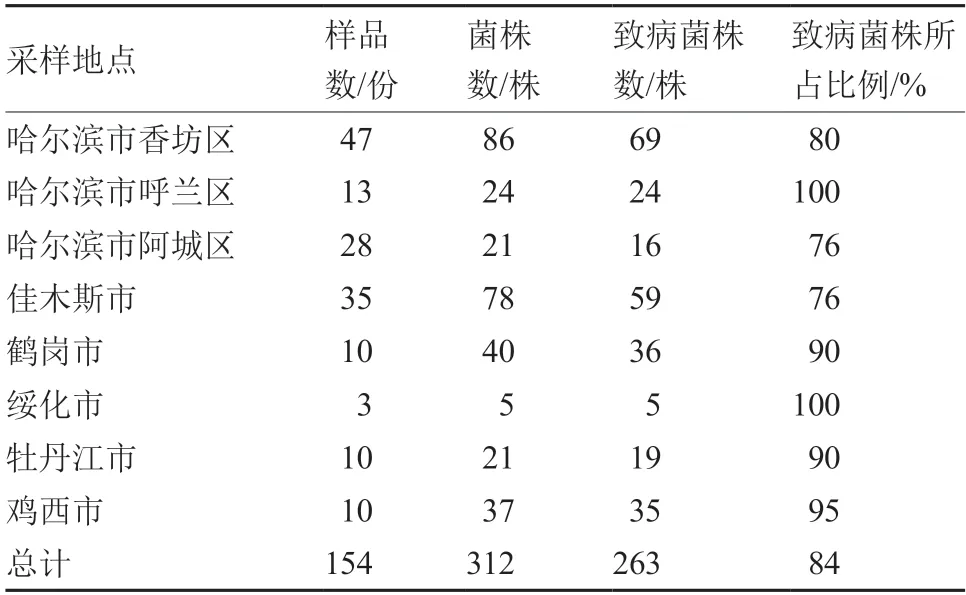
表1 2018—2020 年黑龙江省各马铃薯主产区样品采集与菌株分离统计
2.2 链霉菌菌株的致病性测定
利用萝卜幼苗法测定纯化的链霉菌菌株的致病性,根据链霉菌菌株是否会对萝卜幼苗的生长产生抑制判断其致病能力。结果表明,与空白对照相比,致病菌株使萝卜幼苗的生长受到明显抑制,抑制率均在45%以上,且使根须的数量减少(图1)。通过萝卜幼苗法从分离纯化的312 株链霉菌菌株中筛选出263 株致病链霉菌,致病菌株所占比例达到84%。各地区致病菌株数及所占比例有差异,在哈尔滨市香坊区、呼兰区和阿城区3 个地点共分离链霉菌菌株131 株,致病菌株为109 株,所占比例约为83%,其中香坊区致病链霉菌菌株所占比例为80%,呼兰区所得链霉菌菌株全为致病菌株,即所占比例为100%,阿城区获得21 株链霉菌菌株,致病菌株占比达到76%;佳木斯市共分离78 株链霉菌菌株,致病菌株为59 株,所占比例为76%;鹤岗市致病链霉菌菌株为36 株,所占比例为90%;绥化市5 个分离菌株中致病菌株所占比例为100%;牡丹江市致病菌株所占比例为90%;鸡西市为95%(表1)。

图1 萝卜幼苗法测定菌株的致病性
2.3 致病链霉菌的鉴定
2.3.1 形态观察 将263 株链霉菌致病菌株在ISP2 培养基平板上黑暗培养7 d,MDJ-1、AC-1、SH-4、JMS-17 和HL-23 等5 个菌株对萝卜幼苗的抑制效果明显,且形态各异,因此选择这5 个菌株进行观察鉴定。MDJ-1 和AC-1 菌株能在ISP2 培养基上产生褐色色素,其余菌株均无色素产生(图2-A)。菌株菌落表面粗糙,形状有圆形和边缘不规则两种,多数菌落菌丝平铺于培养基上,但AC-1菌株的菌落中央凸起;大部分菌株菌落表面能产生无色小液滴,仅SH-4 菌株无液滴产生(图2-B)。MDJ-1 和AC-1 的孢子链为螺旋状,SH-4、JMS-17 和HL-23 的孢子链形态为直柔状(图2-C)。

图2 致病链霉菌色素的产生、菌落形态及孢子链显微形态(400 倍)
2.3.2 生物学特性测定 对5 株具有代表性的菌株进行黑色素产生、硫化氢产生、纤维素利用、明胶液化、淀粉酶产生、碳源利用、氮源利用、敏感性等生理生化特性测定。详细结果见表2。
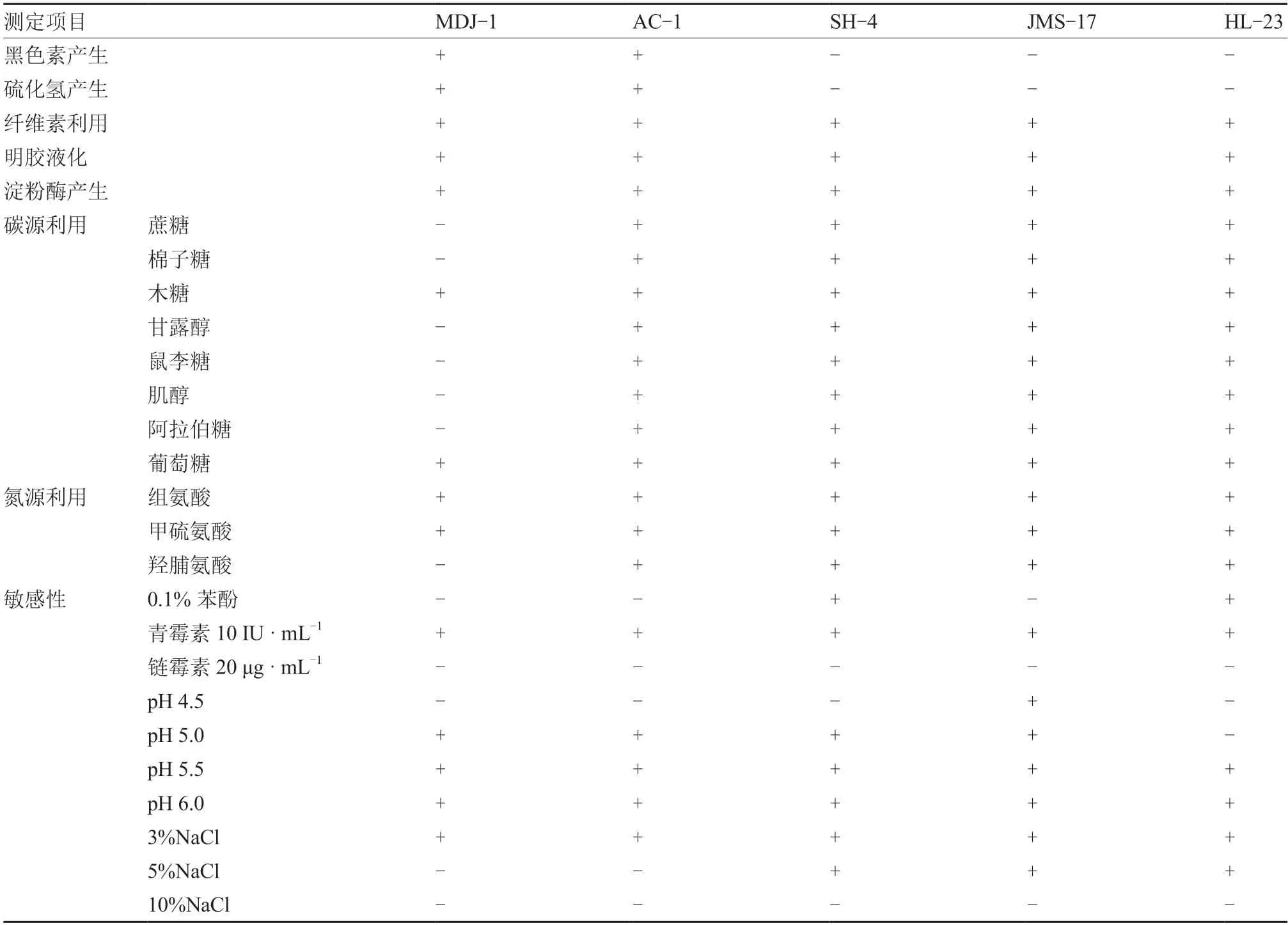
表2 致病链霉菌菌株生理生化特性
2.3.3 分子生物学鉴定 用链霉菌通用引物PA/PH对5 个代表性菌株DNA 进行PCR 扩增,PCR 产物测序后,将所得序列于NCBI 进行序列Blast 比对,并通过Mega 7.0 软件对序列进行多重比对,构建系统发育树,以确定测试菌株的种类。从图3 可以看出,代表性菌株MDJ-1 与亲缘关系最近,序列在NCBI 中Blast 比对同源性达到99%。哈尔滨市阿城区的代表性菌株AC-1 与亲缘关系相近,同源性为99%。菌株JMS-17 与亲缘关系最近,Blast 比对同源性为99%。SH-4 与的同源性为99%,HL-23 与的同源性为99%。

图3 根据16S rDNA 基因序列构建的系统发育树
2.4 黑龙江省马铃薯疮痂病菌种类分析
从黑龙江省6 个市8 个采样点共分离得到、、、、等5 种致病性链霉菌。263 株致病菌株中,127 株为,占比48.3%;14 株为,占比5.3%;46 株为,占比17.5%;24 株为,占比9.1%;52 株为,占比19.8%。各地区病原菌种类结果如下(表3):哈尔滨市有3 种,其中香坊区为(39 株)和(30 株),呼兰区为(24 株),阿城区为(16 株);佳木斯市有2 种,分别为(45株)和(14 株);鹤岗市有(24 株)和(12 株)2 种;绥化市为(5 株);牡丹江市为(19 株);鸡西市为(35 株)。其中,分布最广泛,在哈尔滨市、佳木斯市、鹤岗市和牡丹江市均有分布。
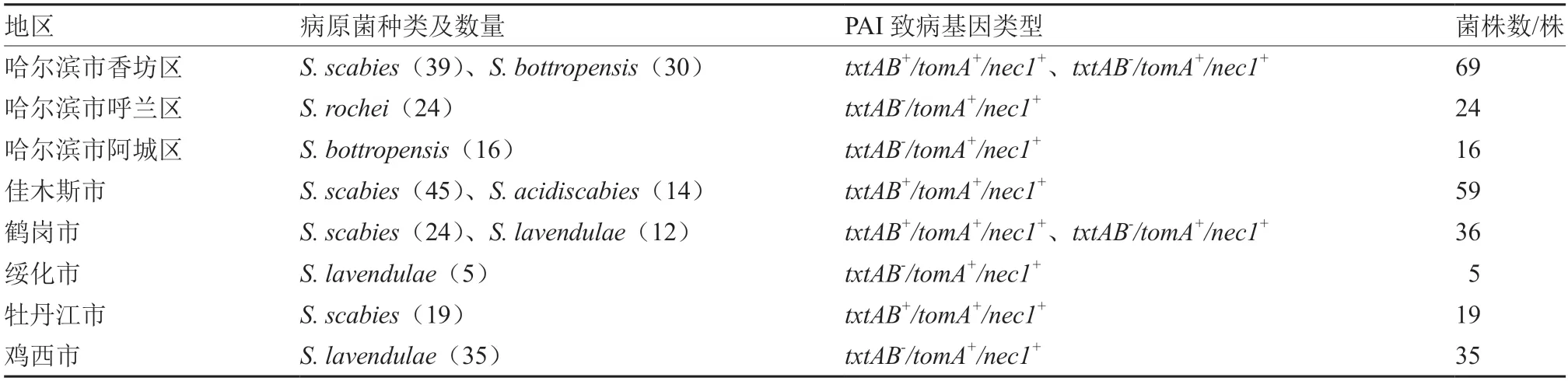
表3 致病链霉菌种属的地区分布
2.5 致病链霉菌PAI 致病基因的检测
对分离的263 株致病菌株进行PAI 致病基因(、和)的检测,用3 种致病基因引物进行PCR 扩增,经凝胶电泳检测,共检测出2 种致病基因类型,分别为和。其 中和的致病基因类型均为。、和致病基因类型均为(表3)。哈尔滨市和鹤岗市均具有2 种致病基因型,其中哈尔滨市香坊区致病基因型和的分离比率为56.5%和43.5%,鹤岗市和的分离比率分别为66.7%和33.3%。
3 讨论与结论
由致病链霉菌(spp.)引起的马铃薯疮痂病是一种严重的土传病害,已报道的病原菌种类较多。目前黑龙江省已报道的链霉菌有、、和4 种病原菌(邢莹莹 等,2016;杨德洁等,2018)。本试验从黑龙江省6 个市采集了154份马铃薯样品,分离出312 株链霉菌菌株,利用萝卜幼苗法测定菌株致病性,共获得263 株致病菌株,结合致病菌株的形态学、生理生化特性和分子生物学特征进行分类鉴定。结果表明,各地区致病菌种类有差异,6 个市共分离到5 种致病链霉菌、、、、,说明黑龙江省马铃薯疮痂病病原菌具有多样性。是分布范围最广的种类,分离率达到全部致病菌的48.3%,、和是黑龙江省首次报道的马铃薯疮痂病菌。
基因的水平转移在细菌进化中发挥着重要作用,使细菌快速获得新的性状以提高对特殊环境(包括与致病相关的真核寄主)的适应能力。细菌菌株可能含有1 个或多个PAI 致病基因,亲缘关系相同的菌株可能含有不同的PAI(增加或完全缺失PAIs),该PAI 是使农业系统中出现新的植物致病链霉菌种的原因(Kers et al.,2005;Lapaz et al.,2017)。致病基因在病原菌致病性中起着重要作用,PAI 致病基因存在种内和种间水平转移的特性,可以从致病链霉菌中转移至非致病链霉菌中,改变菌株的致病基因组成,使致病基因类型多样化、复杂化,最终导致新致病菌的产生(Armijos-Jaramillo et al.,2017)。具有遗传多样性的链霉菌分离株中高度保守的致病基因是测定致病性的理想分子标记。例如,基因已被用作育种计划中潜在致病性的遗传标记,该基因编码一种分泌的坏死蛋白(Joshi et al.,2007),的异位表达足以将非致病性链霉菌转化为致病菌(Bukhalid &Loria,1997;Bukhalid et al.,1998)。有报道认为基因在非亲缘病原链霉菌中结构和功能高度保守,而在非致病性菌株中不存在(Bukhalid et al.,1998;Kreuze et al.,1999)。致病性、和菌株可能同时含有基因和毒素thaxtomin A,但thaxtomin A 的产生不需要(Bukhalid &Loria,1997;Bukhalid et al.,1998)。Joshi 等(2007)研究表明对于致病性是必要的,但也有人认为它在致病性中起辅助作用(Bukhalid et al.,1998),因为Wanner(2004)发现其在链霉菌的其他一些致病菌株中缺失。本试验将分离鉴定的具有代表性的菌株进行PAI 致病基因(、和)的PCR 扩增,通过琼脂糖凝胶电泳检测,紫外凝胶成像系统观察。结果表明,致病基因的组成类型有2 种,即和,大多数菌株均含有全部3种致病基因,、和含有和,但缺乏,是非典型的致病基因组成类型,有些地区缺少含有全部3 种致病基因的分离株,因此,在这些地区对致病链霉菌PAI 基因的组成还需进一步调查。
