海南中部山区天然林物种组成与群落特征变化
2022-05-24李晨笛李东海杨小波史建康赵俊福张培春田璐嘉
李晨笛,李东海,杨小波,史建康,赵俊福,李 龙,陈 琳,张培春,田璐嘉
(1.海南大学 生态与环境学院,海口 570228; 2.海南省生态环境监测中心,海口 571126)
热带森林是一个重要的生物多样性中心,虽然其面积仅占地球陆地表面的7%,但拥有世界50%以上的物种[1−2]。海南中部山区的热带天然林物种组成丰富、区系复杂,对海南生态系统稳定具有重要作用。人类活动的干扰使越来越多的热带天然林正在经历着物种组成和群落特征的改变,是影响生态系统功能正常发挥、影响群落稳定性的主要威胁之一。因20世纪50年代到90年代对海南中部山区热带天然林的大面积砍伐[3−5],低地雨林、山地雨林,甚至是云雾林都受到一定程度的影响。自1994年实施天然林保护工程以来,森林才得到一定程度的恢复。因此,研究海南中部山区受砍伐影响的热带天然林在自然恢复过程中的群落动态变化,对维持海南热带天然林生态平衡、预测群落演替趋势以及制定相应的保护政策具有重要意义。物种组成是群落最基本的特征,森林群落的乔木层优势种,很大程度上主导植被演替方向。群落外貌特征有着深刻的生境灵敏指示性,叶片是植物最敏感的器官之一,体现了植物与其生境条件相适应的自身特征[6]。但是,由于森林物种组成和群落特征动态变化的周期较长,越来越多的学者开始强调自然森林群落长期重复研究的重要性[7]。在国际上,以美国热带森林科学研究中心建立的热带森林监测网络最具代表性[8]。近年来,我国学者在古田山、天童国家森林公园、鼎湖山等地开展了大量有关森林动态的研究[9−11]。在海南,虽然杨小波、方精云、龙文兴等[12−14]对海南植物群落的物种组成与结构进行了较全面的研究,但主要基于1次调查的基础上,建立样地跟踪森林动态的研究工作起步相对较晚[15]。本研究以海南中部山区已建成的4个森林动态样地为对象,基于2012—2020年2次植被调查,探讨受砍伐影响的热带天然林在自然恢复过程中物种组成、区系组成和群落外貌特征变化特点,为研究热带森林恢复过程中物种替换规律、预测海南热带天然林发展趋势提供参考。
1 研究区域及研究方法
1.1 研究区域概况研究区域位于海南岛中部山区四大林区——五指山、尖峰岭、吊罗山、霸王岭 内 , 地 理 坐 标 18°20 ′~ 19°12′N , 108°41 ′~109°56′E。属热带季风性气候,年平均气温为22 ℃,年平均相对湿度为 80%~85%,年平均降水量为2 651 mm。年内雨量分布不均匀,80%雨量集中在5—10月,形成明显的干湿季[4]。山地雨林原始林动态样地设于以陆均松(Dacrydium pectinatum)−线枝蒲桃 (Syzygium araiocladum)−灌丛泡花树(Meliosma dumicola)为优势的五指山山地雨林原始林内,海拔1 035 m,土壤为黄色赤红壤,群落保存完好, 群落演替已达顶级阶段。低地雨林次生林动态样地设于以九节(Psychotria asiatica)−毛柿(Diospyros strigosa)−华润楠(Machilus chinensis)为优势的尖峰岭低地雨林内,受人为活动影响属于次生林,地处沟谷附近,海拔484 m,处于演替中期。山地雨林次生林动态样地设于以琼南柿(Diospyros howii)−岭南青冈(Cyclobalanopsis championii)−黄叶树(Xanthophyllum hainanense)为优势的吊罗山山地雨林次生林内,海拔1 035 m,土壤为山地黄壤,处于演替中期。云雾林次生林动态样地设于以蚊母树(Distylium racemosum)−碟斗 青 冈 (Cyclobalanopsis disciformis)−赤 楠 蒲桃(Syzygium buxifolium)为优势的霸王岭云雾林次生林内,海拔1 365 m,山风强烈、气温低、云雾多, 土壤含水量常处于饱和状态,处于演替中期[14]。
1.2 样地设置与调查为监测海南中部山区具有典型代表性的热带天然林生态系统的长期动态,2012年在五指山山地雨林原始林、尖峰岭低地雨林次生林、霸王岭云雾林次生林,2014年在吊罗山山地雨林次生林内各建立1块1 600 m2的生态学监测样地。样地设置见表1,每片样地以西南角作为原点,采用“相邻格子法”将整个样地划分为20 m×20 m的样方,再将20 m×20 m样方划分为10 m×10 m小样方。首次调查以胸径(DBH)≥1 cm为木本植物起测径阶,采用“每木记账调查法”对每木进行编号、挂牌,记录其胸径、树高、冠幅等。通过查阅《海南植物图志》和Flora of China,对样地内木本植物进行物种鉴定[16]。参照《海南植物图志》《中国热带主要经济树木栽培技术》中各树种的生境分布特点以及树种对光的耐受特性[16−17],将样地树种分为阳性、中性、阴性3个类群。2020年调查方法与首次调查相同,并记录新增胸径≥1 cm的补员个体。

表1 海南中部山区热带天然林森林动态样地设置情况
1.3 S ørensen群落相似度计算群落β多样性采用Sørensen群落相似度计算[18],计算公式为 :
式中,a、b是2个样地中各自的物种数,c是2个样地中的共有种数。
1.4 重要值计算依据物种的重要值的大小确定样地优势种。物种重要值的计算公式为:
1.5 群落区系分析依据吴征镒等[19]的种子植物属的分布区类型进行样地群落的植物区系组成分析。
1.6 群落外貌特征分析依据Raunkiaer的叶级划分系统和Paijimans叶质分类系统[20],对样地群落的叶性质进行分析。按叶级将叶划分为鳞叶、微型叶、小型叶、中型叶、大型叶和巨型叶,共6级;按叶型将叶划分为单叶和复叶;按叶质将叶划分为革质叶、纸质叶和膜质叶;按叶缘将叶划分为全缘叶和非全缘叶。
1.7 数据处理统计分析与制图通过Microsoft Excel 2021和Origin 2021软件完成。
2 结果与分析
2.1 物种组成动态2012—2020年海南中部山区热带天然林各群落物种组成变化见表2。山地雨林原始林,2012年共记录木本植物60种1 106株,2020年物种组成较2012年增加5种,无物种退出,群落总个体减少了21株。低地雨林次生林,2012年共记录木本植物94种892株,2020年物种组成年较2012年增加1种,其中退出7种,新增8种,群落总个体增加了18株。山地雨林次生林,2014年共记录木本植物95种1 380株,2020年物种组成较2014年增加17种,其中退出3种,新增20种,群落总个体增加了199株。云雾林次生林,2012年共记录木本植物54种1 382株,2020年物种组成较2012年增加18种,其中退出4种,新增22种,群落总个体增加了207株。4个森林动态样地的Sørensen群落相似度分别为 0.960、0.921、0.889、0.794,山地雨林原始林物种组成表现出最高的稳定性,随海拔升高,人为干扰减少,热带天然林次生林Sørensen群落相似度降低、物种更替明显。随海拔高度差增加,各群落间Sørensen群落相似度有降低的趋势(表3),低地雨林次生林和云雾林次生林海拔差最大,Sørensen 群落相似度最低,为0.068~0.120。低地雨林位于较低海拔,地貌、地形和水热条件都与高海拔的云雾林存在较大差异,从而导致了物种分布的差异性。2012—2020年,各样地间Sørensen群落相似度多有升高趋势,群落间Sørensen群落相似度平均值由首次调查的0.182增长至末次调查的0.217,多年来有更多相同的物种。
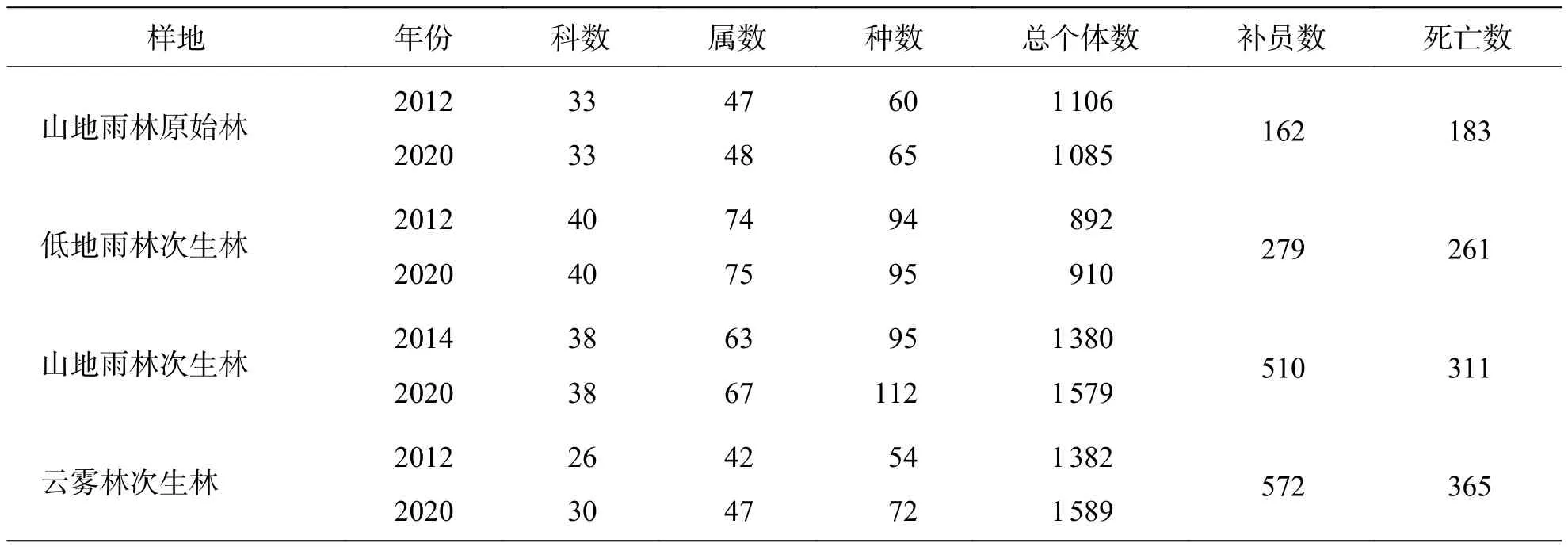
表2 海南中部山区热带天然林各群落物种组成变化

表3 海南中部山区热带天然林各群落间Sørensen群落相似度变化
2.2 优势种动态森林群落优势种对构建群落起着决定性作用,重要值是客观评估群落中物种作用和地位的综合指标,可以有效反映群落优势种在2012—2020年的动态变化。海南中部山区热带天然林各群落优势种重要值变化见图1,山地雨林原始林优势种重要值变动不大,灌丛泡花树由第3升至第1,陆均松、线枝蒲桃下降,乐东拟单性木兰(Parakmeria lotungensis)替代广东山胡椒(Lindera kwangtungensis)进入前10位。低地雨林次生林优势种重要值多有增加趋势,三稔蒟(Alchornea rugosa)、岭南山竹子(Garcinia oblongifolia)替代翻白叶(Pterospermum heterophyllum)、黄椿木姜子(Litsea variabilis)进入前10,九节、翻白叶、假苹婆(Sterculia lanceolata)、黄椿木姜子重要值降低。山地雨林次生林优势种重要值多有不同程度的降低,海南山矾(Symplocos hainanensis)替代陆均松进入前10,四蕊三角瓣花(Prismatomeris tetrandra)、海南山矾重要值增加明显。云雾林次生林优势种重要值多有不同程度的降低,蚊母树重要值由23.32降低至18.10,重要值虽降幅明显,但仍然占绝对优势。五列木(Pentaphylax euryoides)、药用狗牙花(Tabernaemontana bovina)替代厚边木犀(Osmanthus marginatus)、锈毛杜英(Elaeocarpus howii)进入前10,九节、五列木、药用狗牙花重要值增加。4个样地首次调查和末次调查优势种重要值之和占比分别为62.62%、62.16%;44.75%、45.35%;49.55%、49.54%;62.24%、54.39%。
由此可见,2012—2020年海南中部山区热带天然林各群落排名第1的优势种重要值均有不同程度的降低,优势种重要值之和占比较大,为44.75%~62.62%,说明从整体上看,优势种在群落中具有较高的优势地位。云雾林次生林优势种重要值之和占比下降明显,表明在短期演替中,云雾林次生林由蚊母树占绝对优势的单优势种群落向更为混合的多优势种群落发展。其余群落优势种重要值之和占比均保持相对稳定,表明群落优势种未受到强烈干扰,处于稳定演替。随着短期演替进行,喜阳树种重要值多有降低趋势,如广东山胡椒、翻白叶、蚊母树、紫毛野牡丹(Melastoma penicillatum)、厚边木犀等;耐荫树种重要值多有升高趋势,如灌丛泡花树、四蕊三角瓣花等。表明在热带天然林短期演替过程中喜阳树种优势地位降低,耐荫树种优势地位升高。而岭南山竹子等幼树耐荫、壮龄喜阳、繁殖速度快且生长较迅速的树种,在生长的各个时期均处于相对适合的环境,优势地位亦增加明显。
2.3 物种替代规律2012—2020年间,海南中部山区热带天然林山地雨林原始林共记录补员个体32种162株,补员最多的个体为灌丛泡花树、尖峰润 楠 (Machilus monticola)、亨 氏香 楠 (Aidia henryi)、鱼骨木(Canthium dicoccum)、粗叶木(Lasianthus chinensis)。新增物种5种,纤枝蒲桃(Syzygium stenocladum)、变叶榕(Ficus variolosa)、海南鹅掌柴(Schefflera hainanensis)、龙船花(Ixora chinensis)、山龙眼(Helicia formosana)进入群落。死亡个体38种183株,死亡最多的物种为灌丛泡花树、线枝蒲桃、广东山胡椒、拟密花树(Myrsine affinis)、腺叶山矾(Symplocos adenophylla)。无物种退出。
低地雨林次生林共记录补员个体53种279株,补员最多的个体为九节、三稔蒟、毛柿、粗叶木、美脉粗叶木(Lasianthus lancifolius)。新增物种8种,乌材(Diospyros eriantha)、山龙眼、海南大风子(Hydnocarpus hainanensis)、鲫鱼胆(Maesa perlarius)、瓜馥木(Fissistigma oldhamii)、牡荆(Vitex negundovar.cannabifolia)、海南韶子(Nephelium topengii)、 公 孙 锥 (Castanopsis tonkinensis)进入群落。共记录死亡个体61种261株,死亡最多的物种为九节、毛柿、三稔蒟、假苹婆、翻白叶。退出物种7种,箣柊(Scolopia chinensis)、 乌 榄 (Canarium pimela)、 黄 桐(Endospermum chinense)、 罗 伞 (Brassaiopsis glomerulata)、海南蜜茱萸(Melicope chunii)、木荷(Schima superba)、山乌桕(Triadica cochinchinensis)退出群落。
山地雨林次生林共记录补员个体68种510株,补员最多的个体为四蕊三角瓣花、海南山矾、琼南柿、红鳞蒲桃(Syzygium hancei)、蜜茱萸(Melicope patulinervia)。新增物种20种,罗伞、岭 南 柯 (Lithocarpus brevicaudatus)、 三桠苦(Melicope pteleifolia)、铁仔冬青(Ilex chuniana)、刻节润楠(Machilus cicatricosa)、狭叶冬青(Ilex fargesii)、琼中柯(Lithocarpus chiungchungensis)海南狗牙花(Tabernaemontana bufalina)、海桐山矾(Symplocos heishanensis)、两广梭罗(Reevesia thyrsoidea)等树种进入群落。死亡个体60种311株,死亡最多的物种为琼南柿、双叶黄杞(Engelhardia unijuga)、四蕊三角瓣花、红鳞蒲桃、长柄梭罗(Reevesia longipetiolata)。退出物种3种,海南冬青(Ilex hainanensis)、海南猴欢喜(Sloanea hainanensis)、枝花木奶果(Baccaurea ramiflora)退出群落。
云雾林次生林共记录补员个体57种572株,补员最多的个体为海南木犀榄(Olea hainanensis)、紫毛野牡丹、九节、药用狗牙花、变叶榕。新增物种22种,海南木犀榄、黄毛楤木(Aralia chinensis)、广东粗叶木(Lasianthus curtisii)、变叶木(Codiaeum variegatum)、琼岛染木树(Saprosma merrillii)、凸脉冬青(Ilex kobuskiana)、山杜英(Elaeocaepus sylvestris)、冬青(Ilex chinensis)、白背厚壳桂(Cryptocarya maclurei)、红楠(Machilus thunbergii)等树种进入群落。死亡个体41种365株,死亡最多的物种为蚊母树、紫毛野牡丹、厚边木犀、海南鹅掌柴、九节。退出物种4种,石斑木(Rhaphiolepis indica)、山楂(Crataegus pinnatifida)、海南越桔(Vaccinium hainanense)、剑叶冬青(Ilex lancilimba)退出群落。
由此可见,随着短期演替进行,翻白叶、广东山胡椒、厚边木犀、蚊母树等喜阳树种发生过荫死亡,种群死亡个体数量较大;黄桐、木荷、石斑木等喜阳树种退出群落。粗叶木、变叶榕、海南木犀榄等喜阴或幼苗耐荫树种更适应后期生境,种群补员个体数量较大;海南狗牙花、海桐山矾、刻节润楠、两广梭罗、三桠苦、狭叶冬青、变叶榕、白背厚壳桂、红楠、山杜英、冬青等耐荫树种进入群落。随着群落演替,郁闭度增加,群落表现出喜阳的前中期树种逐渐被耐荫的演替中后期种替代的现象。与其他群落相比,云雾林次生林中后期树种更具有耐荫、耐寒、喜湿润的特点,海南木犀榄等喜阴喜湿的树种进入群落后种群快速扩增。调查结果亦显示,随着短期演替进行,纤枝蒲桃、海南韶子、琼中柯、琼岛染木树等分布区窄的海南特有种作为补员树种进入群落,体现出生态位的高度分化。
2.4 区系组成动态对森林动态样地群落木本植物属的分布区类型研究显示(表4),海南中部山区天然林均以热带区分布占绝对优势,为76.2%~94.7%,热带区类型中以热带亚洲(印度−马来西亚)分布、泛热带分布为主;温带区分布较少,中国特有分布极少,仅见于山地雨林原始林样地。表明海南中部山区天然林区系组成具有强烈的热带性质,并具有热带亚洲或印度-马来西亚亲缘,但特有性较弱。随海拔升高,温度降低,呈现出热带成分降低,温带成分增加的特点,低地雨林次生林热带区分布占比最高,云雾林次生林最低。8年间,植物区系地理成分更加复杂,分布区窄的属多有增加趋势。

表4 海南中部山区热带天然林各群落属的分布区类型变化
2.5 群落外貌特征动态叶性质的变化对反映植物适应环境的取向具有重要意义,有着深刻的生理适应基础和生境灵敏指示性。由表5可知,各群落基本呈现出以中型叶、单叶、革质叶、全缘叶为主的热带地区典型外貌特征,沿海拔升高,中型叶比例降低,小型叶比例升高。2012—2020年间,山地雨林原始林,中型叶增加1.9%,单叶增加0.03%,革质叶增加2.06%,全缘叶增加0.27%。低地雨林次生林,中型叶增加0.35%,单叶增加1.11%,革质减少1.35%,全缘叶减少1.44%。山地雨林次生林,中型叶增加5.61%,单叶增加0.02%,革质叶减少4.71%,全缘叶减少1.93%。云雾林次生林,中型叶增加3.41%,单叶减少0.12%,革质叶减少3.48%,全缘叶减少5.03%。8年间,各群落中型叶比例均增加。
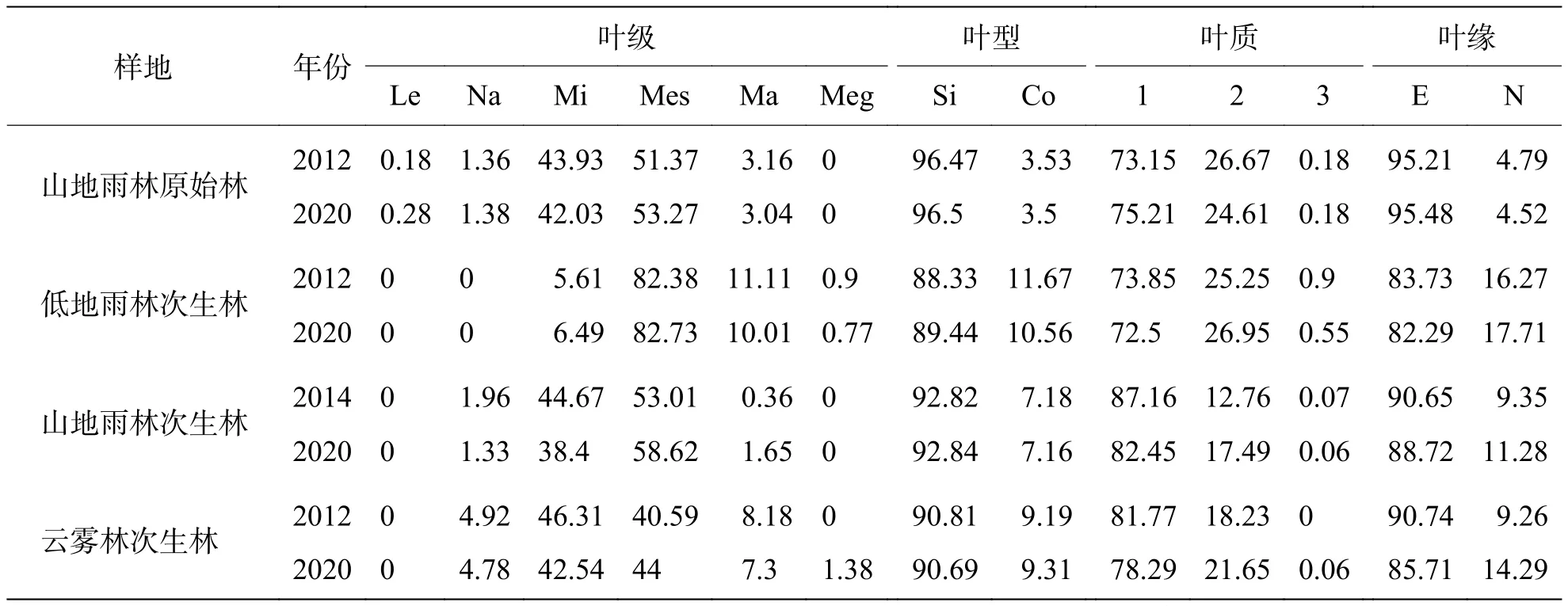
表5 海南中部山区热带天然林各群落叶性质变化%
3 讨 论
3.1 物种组成动态2012—2020年,海南中部山地雨林原始林已演替至顶级阶段,植物彼此间拥有很好的配合且排除新的种类,在组成上并不产生主要的改变,群落最稳定。低地雨林次生林物种退出较多,与海拔低群落易受人为干扰有关,影响了植物繁殖体的定居和生存。山地雨林次生林因海拔高得到较好的恢复,物种组成增加较快。云雾林次生林物种更替最明显,可能是因强烈山风而产生的倒木区域为新树种的进入提供了更多机会[21],林隙、林窗是群落演替中种群自然更新的驱动因子之一,不同的光照、湿度、温度和小气候为小径级植株提供了生存空间[22]。前人研究表明,演替过渡阶段树种具有繁殖速度快、生命周期短、物种周转率高的特点,随着群落郁闭度增加、林中微生境异质性增大,森林结构与功能发生剧烈变化导致物种更替显著[23]。而群落进入成熟稳定阶段时,树种多占有稳定生态位,树木生活史长,物种更替速率较慢[23]。云雾林生境恶劣,物种竞争激烈,群落优势种主要集中于少数物种,但随着短期演替进行,群落排名第1的优势种重要值、排名前10的优势种重要值之和占比均下降明显。这表明云雾林次生林由蚊母树占绝对优势的单优势种群落向更为混合的多优势种群落发展。随着群落径级的增加,优势种优势度降低,群落向着更多物种共存方向发展,物种均匀度更高,种群配置更合理[24]。
β多样性可以有效度量群落演替过程中不同时间物种组成的变化[25]。随着时间推移,各群落间Sørensen群落相似度有升高趋势,多年来有更多相同的物种,说明在气候相似的地区,热带雨林群落演替向着相似趋势发展,与单元顶级学说具有一致性。王泽英[26]发现,上海市3个近自然森林样地在自然恢复过程中,其物种组成相似性增加,初始种植模式相似的2个样地,经过10 a的恢复,物种组成基本趋于一致。
3.2 物种替换规律补员物种和死亡物种对反映群落演替动态和物种替代规律具有重要意义[8,27]。在森林演替初期,群落生境光照优良,土壤湿度较差,有利于阳性树种生长[23]。演替过渡阶段,随着群落郁闭度增加,林内水分充足、但林下光照减弱,使多种阳性树种死亡和种子库种耐荫树种萌发[23]。本研究结果表明,随着郁闭度增加,翻白叶、广东山胡椒、厚边木犀、蚊母树等喜阳树种发生过荫死亡,种群呈衰退趋势甚至退出群落,重要值亦降低。陆均松等上层阳性优势种优势地位虽依旧明显,但更新不良,高大的上层树木可以获取充足阳光,但阴湿的林下生境不利于阳性树种幼苗的继续生长。粗叶木、变叶榕、海南木犀榄等演替中后期种规模扩大,演替中后期种具有喜阴或幼树耐荫的特点,可以在较郁闭的林下自然更新,因而更适应后期生境。群落表现出喜阳的前中期树种逐渐被耐荫的演替中后期种替代的现象。丁圣彦和宋永昌[28]提出,作为我国中亚热带东部地区常绿阔叶林演替前期优势种的马尾松(Pinus massoniana),在演替过程中受木荷等喜光但幼苗耐荫的常绿阔叶树种抑制,使其物质合成能力减弱而分解能力加强,最后逐渐退出群落。汪殷华[29]对古田山常绿阔叶林种群更新动态研究显示,群落阳性树种补员率低,而阴性树种补员率高,柳叶蜡梅(Chimonanthus salicifolius)补员率最高,每年达47.51% ,表现出群落郁闭度高林窗少的生境特征。本研究中纤枝蒲桃、海南韶子、琼中柯、琼岛染木树等分布区窄的海南特有种作为补员树种进入群落,表明随着群落恢复,生态位表现出高度分化,有利于珍稀树种的生存。
3.3 区系组成动态海南中部山区天然林均以热带区分布占绝对优势并以热带亚洲(印度-马来西亚)分布为主,区系组成具有强烈的热带性质,是印度-马来西亚群系的重要组成部分[30]。随海拔升高,温度降低,呈现出热带成分降低,温带成分增加的特点,与苏文苹[31]对高黎贡山、朱华[32]对云南种子植物的区系研究结论具有一致性。2012—2020年,海南中部山区天然林植物区系地理成分更加复杂。过度人为干扰会导致物种区系成分趋于单一,造成狭域分布种、特征种减少,入侵种、广布种增多的现象[33],较少的人为干扰有利于区系组成的丰富[34]。
3.4 群落外貌特征动态叶片是植物对周围生境最敏感的器官之一,群落演替宏观可见标志之一就是植物叶片性质的变化,是构成群落外貌的重要因素,反映出群落的动态与历史。前人研究结果表明,随着纬度降低,温度与湿度增加,落叶树种减少而喜温暖湿润的常绿树种增多,植物叶性质呈现出小型叶、复叶、草质叶、非全缘叶比例降低,中型叶、单叶、革质叶、全缘叶比例升高的现象[35]。赖江山等[36]对世坪常绿阔叶林研究结果显示,群落外貌结构以小型叶(48.97%)、单叶(78.35%)、草质叶(57.22%)为主,全缘叶(51.03%)、非全缘叶(48.97%)比例相差不大,体现了中亚热带北缘常绿阔叶林的特征。本研究中,海南热带天然林外貌结构以中型叶、单叶、革质叶、全缘叶为特征,群落水热条件优良,呈现出热带地区显著特点[20]。与赤道雨林相比,海南热带林中型叶比例减少、小型叶比例增加、大型叶比例很低,这是由于受热带季风气候影响,海南干湿季明显,水湿条件降低,植物为减少水分消耗和降低蒸发量降低了叶面积[30]。张坚强等[37]对珠海淇澳岛次生林研究结果显示,受岛内干季高光、干燥的特殊生长环境影响,淇澳岛次生林呈现出以小型叶(45.3%)、单叶(71.8%)、革质(52.9%)、全缘叶(65.9%)为主的外貌特征。小叶级植物叶片呼吸和蒸腾成本相对更低,同时具有较高的热交换能力,在干燥、炎热、高光的特殊生境下较有优势[38]。体现出不同生境下,植物与其水热条件相适应的特征。从潮湿、温暖到干燥或寒冷地区叶级呈现出由大变小的趋势[33]。随海拔升高,温度降低湿度增加,海南热带林叶面积逐渐缩小。低地雨林次生林地处沟谷附近气候温暖且潮湿,故叶级偏大同赤道雨林相似。云雾林次生林叶级偏小,可能与山风强烈且气温低有关。王希华等[39]提出退化环境将使常绿阔叶林中蕨类植物减少,而有利于落叶树种生存,随着退化程度的加深群落中大型叶比例减少,草质叶比例增加。本研究中各群落中型叶比例均有增加的趋势,表明经过有效的封山育林,热带天然林得到一定程度的恢复。
