不同浓度外源玉米黄质对低温弱光胁迫下辣椒幼苗的缓解效应
2022-05-24丁东霞颉建明李能慧韩康宁牛天航
丁东霞,李 静,颉建明,李能慧,韩康宁,张 婧,牛天航,杨 滟
(甘肃农业大学园艺学院 ,甘肃 兰州 730070)
辣椒属于喜温不耐寒蔬菜,是北方地区日光温室主栽蔬菜之一。由于全球气候变化异常,秋冬及早春季节日光温室中低温弱光问题频繁出现,严重影响植株正常的生长发育[1],导致产量和品质严重下降,影响经济效益[2]。大量研究发现低温弱光胁迫显著降低了多年生黑麦草的光合效率[3],而且造成黄瓜幼苗叶绿素含量、净光合速率、光合作用碳同化关键酶RuBP羧化酶活性和叶绿素光化学效率降低,抑制植物生长[4]。此外,低温弱光也严重影响辣椒生长、造成过量活性氧积累进而导致膜脂过氧化、叶绿素含量下降[5]和渗透调节物质发生变化[6]。因此,找到缓解低温弱光的有效措施,提高辣椒对低温弱光胁迫的耐受性是增加辣椒品质与产量的关键。
目前,施用外源植物生长调节剂是有效缓解辣椒低温弱光胁迫的重要途径。已有研究发现外源喷施褪黑素[7]、甜菜碱、水杨酸、独角金内酯[8]、油菜素内酯[9]和5-氨基乙酰丙酸[10]等物质能够通过提高抗氧化酶活性、增加渗透调节物质含量等,进而有效缓解低温弱光胁迫对植株生长的抑制。玉米黄质也称为玉米黄素,普遍存在于植物果实、蔬菜、花卉和藻类植物中,属于基本的天然色素,在动植物体内发挥着重要作用。前人研究集中于玉米黄质对光合器官的保护作用[11-12],也有研究表明玉米黄质对类囊体膜脂具有保护作用[13],且明显高于拟南芥叶片中的其他叶黄素[14]。Tang等[15]在10℃/5℃低温条件下,外源喷施玉米黄质预处理‘航椒4号’辣椒幼苗显著上调玉米黄质合成关键酶基因的表达,提高抗氧化酶活性,增强幼苗对低温胁迫的耐受性。植物体内玉米黄质含量与非生物胁迫的耐受性有一定的关系,金超等[16]研究发现将中华枸杞(LyciumChinese, Miller)的β-类胡萝卜素羟化酶基因(LcCHYB)转入‘White’品种洋桔梗中获得转基因株系,其较野生型玉米黄质含量提高了91.1%,同时提高了非生物胁迫的耐受性。Wang等[17]研究也发现,烟草过表达八氢番茄红素脱氢酶基因(PDS)引起烟草叶片中的色素发生了变化,抗氧化酶的活力提高,抗氧化的能力增强。Wu等[18]在洋桔梗中过表达了β-类胡萝卜素羟化酶基因AtchyB,同时给以强光胁迫,发现转基因植株不仅类胡萝卜素含量增加1.046~3.141倍,而且玉米黄质的生物合成速度加快,含量增加(高出3.344倍),转化植株对强光表现出较强的耐受性。此外也有研究表明,辣椒玉米黄质含量与低温弱光的耐受性呈极显著正相关[19]。然而,玉米黄质外源喷施能否缓解低温弱光胁迫对辣椒的伤害则鲜有报道。
因此,本研究旨在探索外源玉米黄质对低温弱光胁迫下辣椒幼苗生长发育、光合特性和抗氧化系统的影响,并且确定最适玉米黄质浓度,以期为设施辣椒栽培管理和培育设施专用辣椒品种(耐低温弱光)提供科学依据及理论基础。
1 材料与方法
1.1 试验材料及培养条件
供试辣椒品种‘航椒2号’为低温敏感型品种,由甘肃省天水神舟绿鹏农业科技有限公司提供;外源物质为玉米黄质(HPLC≥85%)购自上海源叶生物科技有限公司。试验于2021年3—7月在甘肃农业大学园艺学院设施栽培生理实验室进行。挑选优质饱满的‘航椒2号’辣椒种子,55℃温汤浸种30 min后在常温自来水中浸泡6 h,接着将浸泡好的种子放在湿润的毛巾上黑暗条件下催芽3 d,待种子露白后选取发芽一致(长度1 mm)的种子播种于装满基质的营养钵中(9 cm×9 cm),每钵两粒。出苗后转至人工气候箱进行培养(RDN-400E-4,浙江宁波),培养条件为温度28℃/18℃、光照300 μmol·m-2·s-1,每周浇水两次,浇灌1/2 Hogland营养液一次。
1.2 试验处理
待幼苗长至六叶一心,随机选取长势一致的辣椒幼苗进行处理。将挑选的幼苗分为4份,分别喷施0(CK)、20(T1)、50(T2)、100(T3) mg·L-1玉米黄质溶液,每处理重复3次(每个重复50株辣椒幼苗)。于每天20∶00进行叶面喷施处理,喷施以叶片正、反面形成水滴但不流下为标准,每天处理一次,连续喷施处理4 d后转入人工气候箱进行低温弱光处理(温度15℃/5℃,12 h/12 h;光照100 μmol·m-2·s-1/0 μmol·m-2·s-1,12 h/12 h),处理7 d后选择从上至下第3~4片叶进行相关指标测定。
1.3 指标及测定方法
1.3.1 生长指标测定 随机选取5株辣椒幼苗进行生长指标测定。株高:使用卷尺测定辣椒茎基部到生长点的距离;茎粗:使用游标卡尺测定子叶下2 cm处茎秆直径;地上地下干、鲜重:将辣椒幼苗用超纯水清洗干净后,用吸水纸吸去表面多余水分,分为地上、地下两部分称重即为鲜重,之后置于烘箱后105℃杀青1 h,然后80℃烘至恒重即为干重。
1.3.2 生理指标测定 叶绿素含量测定采用丙酮法。选取处理7 d后辣椒功能叶,避开主叶脉打孔(0.5 cm),称取0.1 g叶圆片,加10 mL 80%丙酮浸提取48 h,期间每隔12 h震荡一次,保证充分提取。最后采用分光光度计(UV-1780,岛津仪器(苏州)有限公司)测663 nm、645 nm下浸提液的吸光值,依据公式计算叶绿素含量。
Chla=12.71×OD663-2.59×OD645
(1)
Chlb=22.88×OD645-4.67×OD663
(2)
Chla+b=20.29×OD645+8.04×OD663
(3)
(1)~(3)式中,Chla为叶绿素a;Chlb为叶绿素b;Chla+b为叶绿素a+b。OD663为浸提液在波长663 nm处的吸光值;OD645为浸提液在波长645 nm处的吸光值。
可溶性糖测定采用蒽酮比色法、可溶性蛋白采用考马斯亮蓝G-250染色法、MDA含量测定采用硫代巴比妥酸法、相对电导率用电导率仪(DDSJ-308A,上海仪电科学仪器股份有限公司,上海)测定,以上指标测定均参照高俊凤[20]的方法。超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)和超氧阴离子含量测定采用试剂盒法(苏州科铭生物有限公司)。
1.3.3 光合参数测定 辣椒植株处理7d后,选择植株功能叶,采用CIRAS-2型便携式光合仪测定辣椒幼苗光合气体交换参数,包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)。
1.3.4 叶片组织NBT染色 染色参照Chen等[21]的方法,最后使用EPSON扫描仪扫描叶片。
1.4 数据处理
用SPSS 22.0对试验数据进行统计分析,并运用Duncan’s法进行差异显著性检验(P<0.05),使用Excel软件和Photoshop CS6作图。
2 结果分析
2.1 外源玉米黄质对低温弱光胁迫下辣椒幼苗生长特性的影响
由表1可知,低温弱光胁迫下不同浓度的玉米黄质预处理均可以不同程度地提高辣椒幼苗各生长指标,且随着玉米黄质浓度的增加表现出先增加后降低的趋势,其中,T2处理下辣椒幼苗各指标增长幅度最大。与CK相比,T2处理株高、茎粗、地上鲜重、地下鲜重、地上干重和地下干重分别提高了5.52%、17.09%、29.80%、46.67%、51.83%和59.89%,且差异均达到显著水平(P<0.05,下同)。由图1(见60页)也可看出CK处理辣椒幼苗长势瘦弱,玉米黄质预处理后各处理辣椒长势增强,尤其是T2处理长势最强。说明玉米黄质能促进低温弱光胁迫下辣椒幼苗的形态生长,增强对低温弱光的耐受性。

图1 外源玉米黄质对低温弱光胁迫下辣椒幼苗生长特性的影响

表1 外源玉米黄质对低温弱光胁迫下辣椒幼苗生长特性的影响
2.2 外源玉米黄质对低温弱光胁迫下辣椒幼苗生理特性的影响
2.2.1 外源玉米黄质对低温弱光胁迫下辣椒幼苗光合色素和光合参数的影响 光合色素表征植物光合作用的强弱,表2为玉米黄质预处理下低温弱光胁迫后辣椒幼苗光合色素含量的变化。数据显示,50、100 mg·L-1玉米黄质处理下叶绿素a、b、a+b含量升高,其中T3处理较CK无显著差异;T2较CK则显著增加,叶绿素a、b、a+b含量分别增加29.12%、23.83%和27.71%。
低温弱光胁迫下导致叶片的净光合速率下降,由表2可知,叶面喷施玉米黄质预处理后可以显著提高辣椒幼苗在低温弱光胁迫下的净光合速率,T1、T2、T3分别较CK提高26.32%、78.95%、15.79%。相比于CK,玉米黄质预处理下细胞间隙CO2浓度下降,T1处理与CK不显著,T2、T3处理分别下降18.64%、9.64%。低温弱光胁迫下玉米黄质处理气孔导度和蒸腾速率较CK显著增加,但T1、T2、T3之间无显著差异。

表2 外源玉米黄质对低温弱光胁迫下辣椒光合色素含量和光合参数的影响
2.2.2 外源玉米黄质对低温弱光下辣椒幼苗抗氧化酶活性和MDA含量的影响 抗氧化酶在消除ROS方面起着重要作用。由表3可知,与CK处理相比,在低温弱光胁迫下喷施玉米黄质预处理能够显著提高CAT、SOD酶活性,且随着玉米黄质浓度的增加表现出先增加后下降的趋势,其中T2处理效果最佳。T2处理,CAT、SOD活性较CK分别提高了381.55%和19.23%,使辣椒能更好地适应胁迫环境。但是玉米黄质预处理对于POD酶活性影响较小,各处理间均未达到显著水平。
低温弱光胁迫处理会导致幼苗叶片中MDA含量升高。由表3可知,喷施外源玉米黄质可以抑制低温弱光胁迫下MDA的积累,其中T1与T2处理MDA含量下降达到显著水平。随着玉米黄质浓度的增加,MDA含量变化表现出先降低后升高的趋势,T3处理MDA含量升高且达到对照水平,T1、T2、T3较CK分别下降25.87%、38.07%和8.93%。

表3 外源玉米黄质对低温弱光胁迫下辣椒抗氧化酶活性和MDA含量的影响
2.2.3 外源玉米黄质对低温弱光胁迫下辣椒幼苗相对电导率的影响 图2所示为低温弱光胁迫下辣椒幼苗相对电导率的变化,随着玉米黄质浓度的增加相对电导率表现出与MDA相同的变化趋势,但是均显著低于CK。T1、T2、T3相对电导率较CK分别下降32.25%、52.28%和32.61%。外源玉米黄质浓度为50 mg·L-1时,相对电导率降低至最低,缓解低温弱光胁迫的效果最佳。

图2 外源玉米黄质对低温弱光胁迫下辣椒相对电导率的影响
2.2.4 外源玉米黄质对低温弱光胁迫下辣椒幼苗超氧阴离子组织化学染色及含量的影响 图3A所示为辣椒叶片组织化学染色显示对超氧阴离子进行定性分析。由图3可见,喷施玉米黄质的幼苗叶片颜色较CK变浅,且随着玉米黄质浓度的增加染色先变浅后变深。但整体都较CK变浅,其中以50 mg·L-1玉米黄质处理的叶片染色最浅,说明T2处理叶片超氧阴离子含量最少。
图3B所示为超氧阴离子含量变化,外源喷施玉米黄质后可以显著降低低温弱光胁迫下辣椒幼苗中超氧阴离子含量。结果显示超氧阴离子含量变化与叶片组织化学染色结果一致,随着玉米黄质浓度增加,超氧阴离子含量先下降后上升,但均低于CK,其中T1、T2、T3分别较CK分别下降6.41%、13.81%、7.30%。组织化学染色和含量变化分析均表示玉米黄质能在一定程度上清除低温弱光胁迫下积累的过量活性氧。
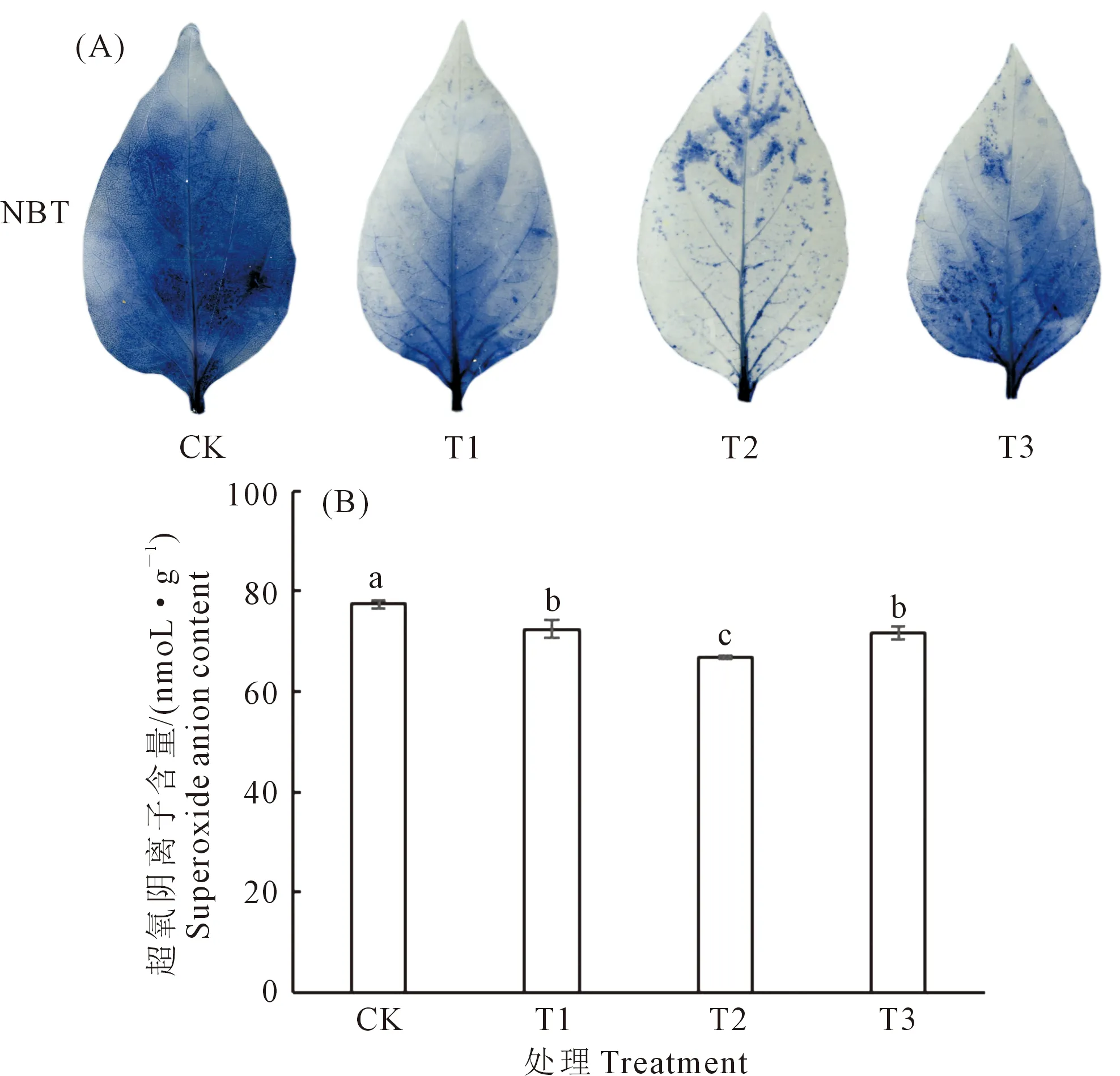
图3 外源玉米黄质对低温弱光胁迫下辣椒超氧阴离子含量的影响
2.2.5 外源玉米黄质对低温弱光胁迫下辣椒幼苗可溶性糖和可溶性蛋白的影响 图4A所示为可溶性糖含量变化,结果显示可溶性糖含量随着玉米黄质浓度的增加呈先增加后降低的变化趋势。T1、T2、T3分别较CK提高了13.83%、22.35%、2.93%,但T3与CK无显著差异。
图4B所示为低温弱光胁迫下可溶性蛋白含量变化,外源玉米黄质处理可以提高辣椒幼苗叶片中可溶性蛋白含量,且随着玉米黄质浓度的增加可溶性蛋白含量呈逐渐增加的趋势。其中T1与CK无显著差异,T2、T3处理与CK相比达到显著水平,分别提高19.58%、22.01%,但是T2与T3之间无显著差异。

图4 外源玉米黄质对低温弱光胁迫下辣椒渗透调节物质的影响
3 讨 论
逆境对植物最直接的伤害是引起形态特征的变化,因此形态指标可以作为判断植株受伤害程度的重要指标[22]。研究表明,低温弱光胁迫可以显著抑制蔬菜作物的生长[23]。在本试验中,胁迫条件下喷施玉米黄质预处理的辣椒幼苗较未喷施的处理长势较强,株高和茎粗分别显著提高了2.09%~5.52%和3.08%~17.09%,能够有效促进生物量的积累,为生长发育提供良好的基础。同时,申磊[24]发现在15℃/5℃、100 μmol·m-2·s-1胁迫下,CoCl2也能提高‘陇椒5号’辣椒植株的形态指标,这可能是外源CoCl2显著提高根系活力,促进植株吸收水分和矿质元素,这与本研究结果相似,即玉米黄质也可能是通过促进根系的生长发育来提高生物量的积累。
叶绿素在光合作用中有吸收、传递和转换光能的作用,其含量高低直接影响植物光合作用,也是衡量植物耐逆境胁迫的重要生理指标之一[25]。光合作用是植物生长发育的基础,为植物提供所需能量和物质,低温胁迫下光系统反应中心对光能利用率降低,对光合作用影响最明显的就是Pn下降,CO2同化受阻[26]。此外,在弱光条件下也会产生过剩的光能,对植物光系统造成伤害,即低温光抑制现象[27]。研究表明,低温弱光或低温处理时,会引起辣椒叶片叶绿素含量[28]、Pn、Tr下降,细胞间CO2浓度升高[29]。本试验发现,在低温弱光胁迫下喷施不同浓度玉米黄质预处理辣椒叶片中叶绿素a、b含量以及Pn、Gs、Tr较CK有不同程度的增加,其中50 mg·L-1处理时最明显,这与28-高芸苔素内酯缓解低温弱光胁迫下辣椒幼苗光合特性的研究结果一致[30]。同时,束胜等[31]研究表明,24-表油菜素内酯可以提高15℃/7℃、150 μmol·m-2·s-1胁迫条件下番茄叶片中1,5-二磷酸核酮糖羧化/加氧酶(Rubisco)的活性和数量,有利于CO2的固定和有机物的积累。外源玉米黄质也可能通过增加气孔导度保证CO2顺利进入叶肉细胞,为光合作用提供原料,提高Rubisco活性,促进碳同化的进行,提高净光合速率,缓解低温弱光胁迫产生的光抑制,且缓解程度随浓度变化。叶绿素含量增加究竟是由于喷施玉米黄质加速其合成还是抑制其降解,其影响机制有待进一步验证。
ROS是植物体内一种重要的信号分子,适宜浓度ROS是植物生长所必需的,但在不适宜的环境条件下会引起植物体内抗氧化酶系统遭到破坏[32],活性氧产生和清除平衡被打破,积累大量活性氧,造成细胞氧化损伤和生物膜损伤[33]。植物体内SOD、POD和CAT是重要的抗氧化酶,能够清除ROS。孟雅宁等[34]研究发现,4叶1心的11个甜椒品种在15℃/5℃条件下处理14 d和21 d后,SOD、POD和CAT 活性显著升高,其中SOD和CAT与植株冷害指数存在显著负相关关系,而POD活性变化与胁迫程度之间无明显关系。本研究中,在低温弱光胁迫下喷施外源玉米黄质预处理后辣椒幼苗SOD和CAT酶活性也显著增加,分别提高了8.98%~19.23%和186.60%~381.55%(表3),同时超氧阴离子含量下降了6.41%~13.81%(图3),说明SOD和CAT酶活性上升能加速清除胁迫条件下积累的过量超氧阴离子。与前人[11,18]研究不同的是,玉米黄质喷施对于POD酶的活性无显著影响,说明低温弱光胁迫下喷施玉米黄质后辣椒体内过氧化物的清除可能存在其他机制,也可能是因为胁迫条件不一致的原因。MDA含量可以反映细胞膜过氧化损伤的程度,相对电导率反映出细胞膜的完整性。低温会导致植物体内MDA含量增加,增加质膜透性对细胞膜造成伤害[35]。低温弱光胁迫下,不同浓度玉米黄质预处理的辣椒幼苗叶片MDA含量和相对电导率与CK相比显著下降,分别下降8.93%~38.07%、32.25%~52.28%,说明喷施玉米黄质能缓解低温弱光造成的损伤,保护细胞膜结构,减少膜质过氧化和细胞内溶质的外渗,维持正常生理代谢。Kang等[36]研究发现过表达番茄红素β-环化酶基因IbLCYB2可显著提高甘薯中玉米黄质含量和SOD酶活性,降低MDA和H2O2含量,提高转基因甘薯抗盐以及干旱胁迫的能力;同样,齐付国等[37]研究也表明-5℃低温胁迫条件下喷施100 μmoL·L-1茉莉酸甲酯显著降低小麦相对电导率,这与本试验结果一致。此外,在本试验条件下,随着外源玉米黄质浓度的升高,相对电导率和MDA含量先下降后升高,这说明外源物质在较适宜浓度下可以缓解低温弱光胁迫,但过高的浓度依然有可能会对植物造成另一种胁迫,这与前人研究褪黑素等生长调节剂的结果相同[38]。
可溶性糖和可溶性蛋白均是植物体内重要的渗透调节物质。在低温弱光胁迫下,辣椒渗透调节物质会发生变化,且可溶性蛋白含量与植株耐低温弱光性呈正相关[6]。本研究表明,外源玉米黄质可以有效促进可溶性糖和可溶性蛋白含量的增加,继而提高细胞液浓度并维持较高的渗透势,防止水分亏缺,提高辣椒幼苗对低温弱光胁迫耐受性,这与Wang等[10]的研究结果一致,即渗透调节物质含量高的植物对逆境表现出更高的耐受性,说明玉米黄质可能提高可溶性糖和蛋白合成关键酶活性使胁迫下辣椒幼苗渗透调节物质含量升高进而增强对逆境耐受性。
4 结 论
50 mg·L-1玉米黄质预处理显著缓解低温弱光胁迫对辣椒幼苗造成的伤害,促进幼苗生长,使低温弱光胁迫下辣椒叶片叶绿素a、b、a+b含量以及净光合速率、可溶性糖、可溶性蛋白显著升高,比CK分别提高29.12%、23.83%、27.71%、78.95%、22.35%、19.58%,抗氧化酶SOD和CAT活性显著增加,比CK分别提高381.55%和19.23%;而辣椒叶片相对电导率、超氧阴离子和MDA含量显著降低,比CK分别降低52.28%、13.81%和38.07%。外源玉米黄质通过提高辣椒幼苗的抗氧化水平,维持保护膜系统的稳定,减少氧化损伤以增强植株的抗性,有效缓解低温弱光胁迫对辣椒幼苗的伤害。
