鸟喙尖头溞的生态分布和培养利用研究进展
2022-05-23周雪妮赵文尹东鹏
周雪妮,赵文,尹东鹏
(大连海洋大学 水产与生命学院,辽宁省水生生物学重点实验室,辽宁 大连 116023)
目前,海水鱼虾育苗所用的动物性活饵料主要有褶皱臂尾轮虫Brachinusplicatilis、卤虫、蒙古裸腹溞Moinamongolica、西藏拟溞Daphniopsistibetana和桡足类等,这些活饵料大多来自内陆盐水或近岸水体,多为广盐性种类,均存在规模化培养要求高或体型小等缺点,不能最大化地发挥其海水养殖业应有的经济效益[1]。随着海水养殖业的迅速发展,在不断提高原有生物活饵料规模化培养技术工艺的基础上,也急需开发新的更适合海水规模化培养且营养丰富的海洋土著种活饵料动物。
鸟喙尖头溞隶属于节肢动物门Arthropoda甲壳亚门Crustacea鳃足纲Branchiopoda枝角类Cladocera栉足目Ctenopoda仙达溞科Sididae尖头溞属Penilia。鸟喙尖头溞属于枝角类中广温广盐性的海洋暖水种[2-4],在全球范围内分布广泛,其具有发育时间短、繁殖量大、易于培养等优势[5]。过去的几十年里,国内外学者对其在全球范围内的季节分布情况进行了系统调查,并探讨了影响其分布及种群动态的因素,如温度[6-11]、盐度[12-16]、叶绿素含量、饵料浓度[17-21]及其他生物因素[22-26]等,但关于其培养利用方面的研究较少。本文概述了鸟喙尖头溞在国内外的生态分布和种群动态,以及大规模培养利用的研究进展,以期为鸟喙尖头溞作为新一代优质海水鱼类稚幼鱼饵料及作为监测生态环境变化的指示生物提供参考。
1 鸟喙尖头溞的生物学特征
鸟喙尖头溞为雌雄异体,雌性体大(0.70~1.09 mm),而雄性体略小,成体雌溞最显著的形态特征是吻呈鸟喙状,壳瓣有腹缘壳刺,第二触角刚毛式为2-6/1-4,尾爪有2根基刺;该溞体短,左右侧扁且分节不明显[2]。鸟喙尖头溞一般在较温暖的季节以浮游生物的形式存在,在较寒冷的季节以底栖休眠卵的形式存在[27]。鸟喙尖头溞的休眠卵为椭圆形或扁形,呈暗黑色,长、宽、高分别为250、180、100 μm,分布在海底从表层到2 cm深的海泥内,休眠卵数量占总量的60%~100%,而6 cm以下的海泥中未发现休眠卵存在。休眠卵保存温度越高,孵化率越低,在18 ℃时孵化率最高。
2 国内鸟喙尖头溞的生态分布和种群动态
2.1 生态分布
鸟喙尖头溞是一种沿岸低盐性种类,具有较广的适温范围,其地理分布主要受食物、温度、盐度和海流等影响,其中受温度影响最大,是上层鱼类(如鲱、鲐、沙丁鱼、蓝圆鲹等)特别是其幼鱼的饵料[2]。鸟喙尖头溞在中国近海分布较广,在河口也有少量分布。夏季鸟喙尖头溞在渤海、黄海和东海沿岸均有分布,春季因温度不高,只在渤海出现,在黄海、东海沿岸均未出现[1]。在中国香港北部近海水温为16~32 ℃、盐度为7.3~37.2范围内,发现有鸟喙尖头溞分布[6]。徐兆礼等[4]调查发现,鸟喙尖头溞春季在水温较高的浙江、福建附近海域,以及海南附近的雷州半岛近岸水域中存在,夏季在东海北部近海和长江口外水域有较多数量,丰度较高时的水温为25 ℃、盐度为14,且从雷州半岛到海南岛东南部的沿岸水域也有大量分布,其密度最高时达1 121.5 ind./m3[7]。阙江龙等[8]于2012年在属南亚热带海洋季风气候的广西近海,经春、夏、秋、冬4个季节的调查发现,鸟喙尖头溞在4个季节均有出现,且为冬、春、夏3季的浮游动物优势种,尤其在冬季,为绝对优势种,在北部湾西北部广西近海的沿岸及湾内水域有较大数量出现,而近海水域平均丰度明显降低,优势度为0.65。高倩等[12]对春季中国从南到北的灌河口、长江口、瓯江口、椒江口、九龙江口和北仑河口6个典型河口调查发现,鸟喙尖头溞仅存在于最南端的北仑河口[12]。
根据近年来的调查发现,在中国,鸟喙尖头溞在温度范围为16~32 ℃时可以存活,且水温为25 ℃时生长状态最为活跃;盐度范围在7.3~37.2时可以存活,且水体盐度为14时生长状态最为活跃。
2.2 种群动态
温度、盐度、叶绿素含量等环境因子及环境污染是影响鸟喙尖头溞种群动态变化的主要因素。调查发现,广东省大亚湾近海海域主要的暖水性枝角类是鸟喙尖头溞,因海水温度上升,鸟喙尖头溞出现的频率也有明显变化,1987年8—12月该海域鸟喙尖头溞的数量从最大值3 225.3 ind./m3急速下降至3 ind./m3,12月更是下降至0.5 ind./m3,且有些区域已绝迹;2007—2008年,在温度较低的5月该溞出现高峰,9月温度较高时该溞数量急剧下降,12月又有所增加[9];2001年对广东省水产自然保护区大亚湾大鹏澳和澳头海域中的两种海洋枝角类数量的变动进行了短期和长期的调查,结果显示,采获的枝角类主要是鸟喙尖头溞,无论是长期还是短期,两个海域中鸟喙尖头溞的密度均高于其他浮游动物,同时夏春两季的数量高于冬季[10]。这表明,温度是影响大亚湾海洋枝角类数量的重要因素之一。此外,人类活动及网箱养殖对环境产生的污染对海洋枝角类数量变动也有一定影响。吴玲玲等[13]研究发现,肥胖三角溞个体数量的增加与鸟喙尖头溞个体数量的减少呈显著正相关,表明二者的生态位接近。同时,黄渤海水域的鸟喙尖头溞数量出现爆发性的激增,可能与该海域的叶绿素含量升高有关,也可能由于环境条件较优越,导致鸟喙尖头溞通过孤雌生殖造成个体数量的爆发性激增[14]。杜飞雁等[11]于2008年调查发现,鸟喙尖头溞在大亚湾近岸海域出现的高峰期不再是夏季,而是有所推迟或提前,且由于季节变化鸟喙尖头溞出现的数量变动也没有以往明显。
综上所述,鸟喙尖头溞种群的动态变化大多是由人为或自然环境变化使海水温度上升所导致。鸟喙尖头溞若与其他溞共同处于适宜的环境条件时,鸟喙尖头溞的种群数量会大幅度激增,乃至超过其他溞类,成为当地的优势种。此外,叶绿素含量也是影响鸟喙尖头溞种群动态的主要环境因子,若浮游植物爆发性激增,可能会引起鸟喙尖头溞数量大幅度上升,对现有浮游植物种群动态产生影响,未来可以尝试开展利用鸟喙尖头溞防治赤潮的研究。
3 国外鸟喙尖头溞的生态分布和种群动态
3.1 生态分布
在全球范围内的调查显示,大多数海洋枝角类都有特定的地理分布,而鸟喙尖头溞没有显示出明显的地理分布[28]。比斯开湾表层水域中的鸟喙尖头溞主要出现在夏、秋两季[20];加泰罗尼亚海陆架鸟喙尖头溞在6月首先出现在南部地区,那里的气候较为温暖,在夏季种群逐渐向北移动[26]。可见,影响鸟喙尖头溞在全球分布的最主要的因素是温度,无论是国内还是国外,鸟喙尖头溞出现频率较高的季节均是夏季,其次是春、秋两季。1999年秋季,大西洋东北部的北海出现了大量的鸟喙尖头溞,该物种的迅速出现与异常温暖的海洋表面温度相一致,自1999年以来,这一物种的存在已经成为北海秋季浮游动物群落的特征,而且由于海水温度升高,有利于鸟喙尖头溞种群的繁殖,在2002和2003年,该物种的数量比以往记录的还要多[29]。鸟喙尖头溞在世界范围内的分布主要集中在亚洲、欧洲、非洲三洲处于热带的交界处,另外,在北美洲及南美洲也有少量分布,且主要集中在夏、秋两季。
3.2 种群动态
3.2.1 水环境因素 在全球变化条件下,呈现出诸多影响鸟喙尖头溞种群动态的因素。达达尼尔海峡的所有枝角类中,鸟喙尖头溞占比高达49%,具有非常可观的种群数量[15]。加蒂斯湾夏季时鸟喙尖头溞的种群密度最高,且与温度呈正相关,与盐度和水深呈负相关[16]。阿拉伯海东南部热带沿岸上升流生态系统中,冬季风和春季风季节桡足类是优势类群,而在夏季风季节,则以鸟喙尖头溞为主的枝角类为优势类群[22],这可能与叶绿素a含量、优先食物供应和良好的环境条件(温度和盐度)有关。9月在土耳其黑海南部锡诺普海的哈姆斯洛斯湾(Hamsilos Bay)出现大量鸟喙尖头溞,导致其丰度和生物量明显升高[23]。2005年11月—2006年8月,对伊斯肯德伦湾4个监测站表层海洋枝角类的调查发现,鸟喙尖头溞最大密度出现在2006年5月,最小密度出现在2005年11月[24]。
3.2.2 生物因素 鸟喙尖头溞的数量不仅会因为一系列原因剧增,与此同时,越来越多的研究报告显示,全球水母数量的增加可能会对海洋浮游生物的食物网动态产生影响。马尔拉马地区原夏秋优势种鸟喙尖头溞于2006年秋季可能因此消失[18]。可见,有些地区鸟喙尖头溞的数量会受到某些因素的影响,浮游动物数量的减少以及影响其减少的因素对远洋生态系统和社会经济的破坏性值得关注。分析原因,这可能与该区域内饵料浓度的增加,以及鸟喙尖头溞出现区域附近潜在捕食者捕食压力的降低有关。Kang等[17]研究发现,夏季夜光藻与鸟喙尖头溞在同一地点出现时,鸟喙尖头溞呈突然增长趋势。在大洋洲金海湾附近、大西洋中南部、大西洋西南部的阿根廷海及地中海西南部均在夏季出现鸟喙尖头溞,其中,在地中海西南部该溞的种群密度最高,可达2 500~3 000 ind./m3,在夏末秋初该种群数量急剧下降,并开始产生休眠卵,以确保种群在下一个繁殖季节的连续性[19-21]。除产生休眠卵外,世代周期短、孤雌生殖和持续的体细胞生长也是鸟喙尖头溞在季节性出现期间也可成功繁殖的3个主要原因[27,30]。
3.2.3 物理因素 在巴西乌巴岛内陆架上鸟喙尖头溞全年出现,温度范围较宽(14.8~28.2 ℃)。在温暖季节,密度通常较高(>2 000 ind./m3),这是由于寒冷和营养丰富的南大西洋中部水与底部水的水层交换而形成的水柱分层所导致[31]。牙买加金斯敦港属亚热带水域,鸟喙尖头溞全年均有出现,平均密度为1 821 ind./m3,生物量(不包括卵、胚)为2.87 mg/m3,生殖力随体型增大而增大,丰度、生物量及繁殖力无明显的季节变化[32]。在瓜纳巴拉湾鸟喙尖头溞的数量在3月达到最高,在5—8月种群几乎消失[33],调查发现,潮汐和夜晚时间是影响其密度的主要因素,由于水温及盐度在当地较为恒定,所以其对溞密度几乎没有影响[34]。近年来研究发现,海洋枝角类包括鸟喙尖头溞的现存量和种类组成的时空变化在较大程度上同时受温度、天敌及海洋环流与锋面流变化的影响[35-36]。Valentin等[37]建立了一个模拟巴西热带瓜纳巴拉湾鸟喙尖头溞种群动态的生态模型,研究发现,捕食是影响当地鸟喙尖头溞死亡的主要因素,但观察到的种群数量急剧减少,不能仅用生殖方式从孤雌生殖转为有性生殖来解释,也可能包含其他因素,如物理因素中潮汐和风驱动下的水团交换均会影响其种群数量。Marazzo等[38]研究发现,在巴西瓜纳巴拉湾只有夜间捕获的鸟喙尖头溞中有成熟胚,而在白日捕获的却未有,胚胎成熟在黑暗期间的一个优势是减少对生殖期雌性的捕食,增加出生率,进而提高种群数量。
综上所述,在当今全球环境下,温度、叶绿素a、优先食物供应及浮游植物在一定范围内与鸟喙尖头溞的种群数量呈正相关;盐度、分布深度、日照时间及海洋环境中的天敌在一定范围内与鸟喙尖头溞的种群数量呈负相关。季风、潮汐及海洋环流在不同程度上对鸟喙尖头溞的种群数量也存在影响。由此可见,鸟喙尖头溞可以尝试作为气候变化的预报生物,预知季风、潮汐及海洋环流的走向和起止时间,还可以作为海洋环境监测的指示生物,监测海洋环境中叶绿素及浮游动植物的种群动态,或者进一步作为海洋污染的指示生物。
4 鸟喙尖头溞的规模化培养
4.1 培养饵料
枝角类富含多种维生素和蛋白质。国内外学者对鸟喙尖头溞所摄食藻的种类、大小及规模化培养等方面进行了诸多试验[39-40]。鸟喙尖头溞适宜在水温19~20 ℃、盐度24~28的条件下进行大规模培养。日本学者在鸟喙尖头溞培养试验中发现,在前3周种群数量无明显变动,30~35 d时种群数量快速增长,44~63 d后达到初始种群数量的100倍[41]。
鸟喙尖头溞规模化培养的重要条件之一是饵料的选择。研究发现,鸟喙尖头溞培养中,使用小球藻做饵料较面包酵母培养的效果更好[5],且对硅藻的摄食率更高[42]。采用高效液相色谱法(HPLC)对鸟喙尖头溞的食物选择性进行分析,通过对浮游植物色素组成的比较,发现鸟喙尖头溞优先选择硅藻、隐藻,而未摄食甲藻,只有在浮游植物种群以甲藻为主的情况下,才能在其体内发现甲藻的标记色素,与此同时,鸟喙尖头溞所摄食浮游植物的密度也会对其生长产生影响[42]。随着食物密度的增加,鸟喙尖头溞的滤水率呈倒“U”曲线型,且食物粒径在15~70 μm时效果最好,最利于种群繁殖[43]。对比3种生物饵料大规模培养情况(表1),西藏拟溞选用蛋白核小球藻Chlorellapyrenoidosa、牟氏角毛藻Chaetocerosmuelleri、盐生杜氏藻Dunaliellasalina和湛江等鞭金藻Isochrysiszhanjiangensis混合投喂,第33天时种群密度达到最高,为(9 500.00±550.73)ind./L[44];蒙古裸腹溞第36天时密度最高,为4 631 ind./L[45-46];鸟喙尖头溞在第44~63天期间达到最高密度,为20 000 ind./L[5]。虽然鸟喙尖头溞培养时种群密度到达最大值的时间长于蒙古裸腹溞和西藏拟溞,但最适放养密度大于蒙古裸腹溞和西藏拟溞,且其生长发育阶段所摄食的饵料较为单一便于培养,因此,鸟喙尖头溞作为海水生物饵料的补充可进一步规模化培养。
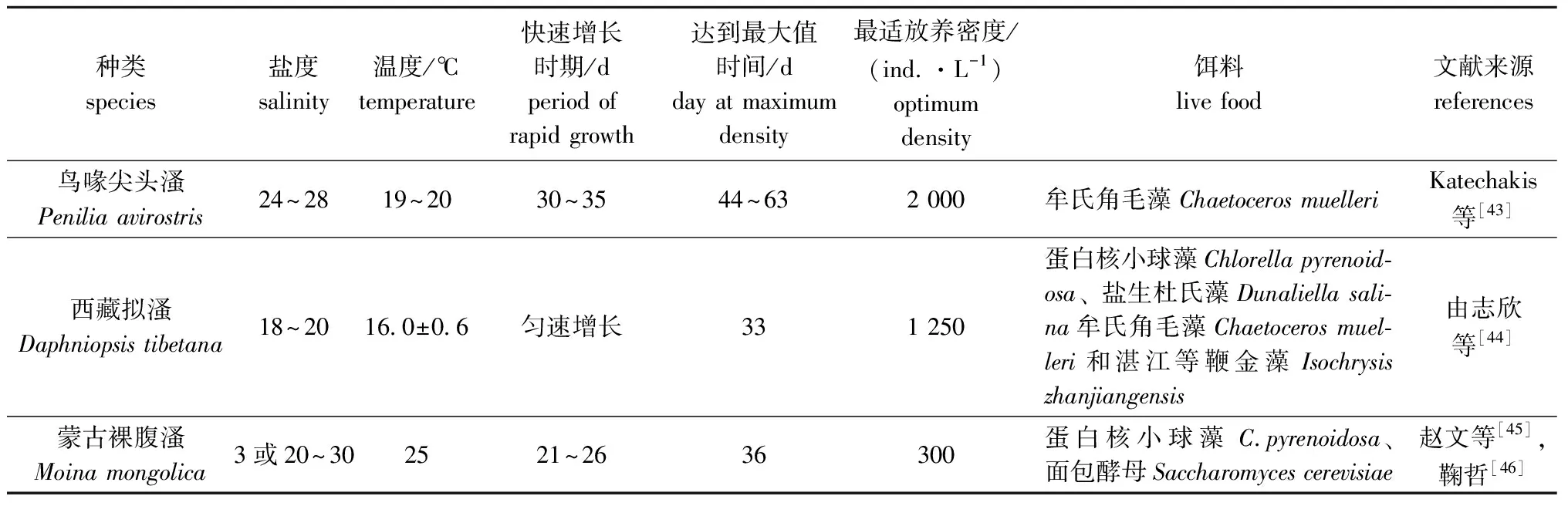
表1 鸟喙尖头溞、西藏拟溞和蒙古裸腹溞大规模培养情况
4.2 培养密度
适宜的种群密度也是培养鸟喙尖头溞的关键。当鸟喙尖头溞种群密度增殖到20 000 ind./L时开始收获,每周收获一次,且每次收获现存量的50%,其密度波动最小[5]。Marazzo等[38]研究发现,当种群密度过高时,会出现两性生殖现象,产生休眠卵,于是该研究者提出了恒定种群性别比的假设,即两性生殖的出现是一个函数,这种现象并不是由于不利的繁殖环境造成,而是由于在种群中雄性的出现率较低且恒定所造成。与此同时,种群密度过高会引起各种细菌及原生动物大量繁殖,虽然这些细菌和原生动物可以作为鸟喙尖头溞的食物,但如果密度过高也会造成水质污染,影响枝角类种群增长,必要时可以加入抗生素以改善培养水环境。此外,培养的优劣程度可用体色的深浅作为参考,体色越深则溞活力越好[2]。鸟喙尖头溞的体色深浅是否可以作为判断其营养价值高低和营养物含量的参考指标,还有待进一步研究。
5 存在问题及展望
5.1 鸟喙尖头溞培养利用中存在的问题
尽管关于鸟喙尖头溞的形态、分布和室内培养方面已有一些研究,但还存在如下问题亟待解决。
1)在全球环境变化背景下,鸟喙尖头溞的生态分布需要跟踪监测。
2)在规模化培养过程中,鸟喙尖头溞相对于西藏拟溞、蒙古裸腹溞等盐水枝角类,其种群密度达到最大值的时间较长,其营养成分和营养强化方面均未见报道。这些因素均严重影响了该溞的进一步培养利用。
3)在海洋污染监测和评价方面尚缺少海洋土著枝角类测试生物,而鸟喙尖头溞具有作为海洋污染测试生物的潜力,目前国内外关于该溞生态毒理学的研究均未见报道。
5.2 未来重点研究方向
众所周知,相对淡水和内陆盐水枝角类而言,典型海洋枝角类种类较少。鸟喙尖头溞是典型的海洋枝角类,作为海洋浮游动物群落的重要成员已显示出其重点保护和开发利用价值。关于鸟喙尖头溞的生态分布情况国内外学者报道较多,并对其大规模培养展开了一系列初步试验,取得了显著成效,但相对于其他盐水枝角类如蒙古裸腹溞、西藏拟溞、轮虫等,其规模化培养工艺优化、营养成分及营养强化方面的研究仍需要进一步开展试验,对鸟喙尖头溞毒理学研究也刻不容缓。未来关于鸟喙尖头溞的研究与利用应从以下几方面重点开展工作。
1)深入开展鸟喙尖头溞生物学研究。随着全球环境变化对海洋环境的不良影响加剧,未来关于鸟喙尖头溞对气候变暖和水体富营养化的响应机制,以及该溞在全球环境变化背景下的海洋生态系统中的种群动态、功能作用及其与其他海洋生物种间的关系尚需深入研究,同时对该溞的遗传多样性及作为模式生物的可行性研究也非常重要。也可从微藻-鸟喙尖头溞-鱼虾三级食物链角度,探讨海洋生态系统能量流动、物质循环和信息联系等生态学理论和应用问题。
2)加强规模化培养工艺优化研究。在生产应用方面,鸟喙尖头溞的规模化培养在最适温度、盐度及饵料方面均已有较为完善的研究,但是对于其驯化方面尚未见报道。可通过改变鸟喙尖头溞的饵料组合,或者通过调整其生活水体的盐度和温度,以缩短其到达最大值的时间,缩短快速增长期,为鸟喙尖头溞的规模化培养奠定基础,为其作为新型优质育苗活饵料开拓前景。鸟喙尖头溞在不同发育阶段体长规格有所不同,可以作为活饵料规模化培养,在更多水产经济动物育苗和养成中加以应用。另一方面,也应尽早开展该溞作为水产经济动物优质饲料蛋白源或人类功能食品的可行性研究。
3)加强营养强化机制和技术手段研究。利用生物学及分子生物学等多种技术手段,对营养强化后鸟喙尖头溞的脂类及脂肪酸组成的改变进行检测,以及对生理代谢有关酶的活性进行研究。主要方法可通过改变浮游动物的饵料,如在饵料中添加酵母、鱼油、维生素、叶酸等营养成分,从而改变鸟喙尖头溞的营养成分组成,或者通过改变其生活条件,如水温、盐度、pH及光照等,研究最适的环境条件组合方式,以提高育苗成效,降低育苗成本。在培养过程中,鸟喙尖头溞的体色与其营养成分间有着密不可分的关系,今后的试验过程中应不断完善判断鸟喙尖头溞培养的标准,进一步开展其体色与营养价值间的关系研究。
4)开展生态毒理学相关研究。由于全球诸多环境监管部门对水体环境问题持续高度关注,需要选择一种易培养且低成本的水生生物作为检测水质的指示生物。鸟喙尖头溞具有生活周期短、易培养、培养过程中不污染水质、全球分布范围广且适应高盐度生活环境等特点,这些优势使鸟喙尖头溞在海洋水体环境监测的实际应用中较盐水枝角类更具有优势,这对于海洋污染风险评估十分必要。应加强该溞在温度、盐度、pH、海流等环境因素的改变,以及在常见海洋污染物(石油、有机污染物、紫外线、化工废液、农药和重金属等)的影响下,其生理或行为(摄食能力、呼吸能力、游泳能力、感光能力和繁殖能力等)反应机制的研究,并开展相关免疫组织化学研究,加强相关基因分析,为其作为环境监测的指示生物开拓前景。
综上所述,鸟喙尖头溞在世界范围内分布广泛,其种群丰度存在地理和季节差异,其实验种群的生态学及其培养利用尚处于初始阶段。随着鸟喙尖头溞生物学、规模化培养技术工艺、营养强化机制、生态毒理学研究的不断深入,鸟喙尖头溞将成为更优质的育苗活饵料及环境监测指示生物的研究必将受到高度重视,并取得更加蓬勃的发展。
