25个苜蓿品系在淮河中下游地区的适应性及遗传多样性
2022-05-23何庆元张宇龙徐智明李正鹏詹秋文
何庆元,张宇龙,韩 伟,徐智明,李正鹏,任 义,杨 豪,詹秋文
(1. 安徽科技学院生命与健康科学学院, 安徽 凤阳 233100;2. 秋实草业有限公司, 安徽 五河 233300;3. 安徽科技学院农学院, 安徽 凤阳 233100)
随人们生活水平提高,奶产品需求增大,优质牧草的生产在农区牧草生产中越来越重要[1],将南方农业耕作制度由过去粮经二元结构调整为粮经饲三元结构模式成为农业产业结构调整的重要举措[2]。苜蓿(Medicago sativa)是世界上栽培面积最大的豆科牧草,具有草产量高、粗蛋白质含量在20%左右、富含奶牛所需的各类营养因子和良好的适口性,并且生物固氮能力强等优点,被称为牧草之王[3]。然而,长期以来,由于南方气温高、湿度大,部分地区土壤呈酸性且质地黏重、地下水位相对较高等对苜蓿生长不利的条件,导致南方苜蓿推广中合适的品种较少[4-5],但随着草产业的迅猛发展,筛选出适合中部和南方种植的苜蓿品种十分必要。
淮河中下游地区水热资源丰富,其环境具有发挥苜蓿高产潜力的基础。近年来,虽然有少量关于该地区苜蓿生产相关的部分报道[6-7],但该地区品种选择还比较盲目,在该地区系统开展引种区域适应性试验,并对该地区野生苜蓿资源进行鉴定和比较试验对生产上有一定指导意义,为利用野生苜蓿资源和选育适合该地区种植的苜蓿品种充分发挥苜蓿的产量潜力奠定基础。苜蓿秋眠性与产量和适应性密切相关,是苜蓿引种栽培的重要依据[8],并且不同休眠的苜蓿的遗传多样性存在差异[9-10]。遗传的丰富度是品种产量、品质和适应性的本质基础。本研究在安徽淮河中下游地区选取了25份国内外苜蓿材料分析其表型和基因型遗传多样性,并进行生产适应性及品质比较,旨在为该地区苜蓿生产中选择适宜品种提供指导和育种利用奠定基础。
1 材料与方法
1.1 试验地概况
2017年10月18日将试验材料播种于安徽省蚌埠市五河县朱顶镇秋实草业有限公司试验基地(117°52′ E, 33°18′ N),该地区位于淮河中游下段、淮北平原东南部、半湿润农业气候区,日照时数年均2 306.7 h,无霜期年均212 d,年降水量896.3 mm。年均气温14.7 ℃,1月份最冷,月平均气温0 ℃;7月份最热,月平均气温27.8 ℃。
1.2 试验材料
25个苜蓿品系,9个购自北京正道生态科技有限公司,6个购自百绿国际草业有限公司,6个由北京克劳沃生态科技有限公司馈赠,4个由安徽科技学院安徽省饲草育种与利用重点实验室保存提供。品系名称及原产地如附表1所列。

表1 不同品系对产量相关性状的影响Table 1 Effects of different varieties and lines on yield-related traits
1.3 试验设计与田间管理
采用随机区组试验设计,小区面积9.0 m2(3 m ×3 m),行距0.3 m,每个小区10行,按22.22 kg·hm-2播种量条播,3个重复。常规大田管理,人工锄草,及时防治病虫害。在苜蓿小区约50%开花时刈割,分别于2018年4月28日、6月5日、7月17日、8月23日和10月11日刈割5次,2019年4月25日、5月27日、7月5日、8月6日和10月11日刈割5次。
1.4 产量相关性状测定及方法
株高是地表到植株顶端生长点的绝对高度,主茎节数从子叶节开始计算,株高和主茎节数是每个小区随机抽样10株的平均值。鲜草产量:取中间8行称量小区鲜草重,并换算为每亩(666.67 m2)鲜重。
1.5 品质相关性状测定及方法
每个小区500 g鲜草,经105 ℃杀青30 min,后经80 ℃烘干至恒重,将全部样品粉碎,混合均匀,过0.425 mm孔径筛,用于粗蛋白(crude protein,CP)、粗脂肪(ether extract, EE)、中性洗涤纤维(neutral detergent fiber, NDF)和酸性洗涤纤维(acid detergent fiber, ADF)含量测定。品质性状测定方法:CP按凯氏定氮法(GB/T6432-1994),EE用索氏提取法(GB/T6433-2006),NDF、ADF用笵氏测定法(Van Soest)[11]。相对饲用价值(relative feed value, RFV)利用NDF和ADF计算,RFV = DDM × DMI/1.29;其中可消化干物质(digestible dry matter, DDM) =88.9 - (0.779 × ADF),粗饲料干物质随意采食量(dry matter intake, DMI) = 120/NDF[12]。
1.6 DNA提取及SSR分析方法
从每个品系采集3株苜蓿叶片,混合,经液氮研磨,用改良的CTAB法提取基因组DNA[13],琼脂糖凝胶电泳检测条带清晰,-20 ℃保存备用。
依Sledge等[14]发表的苜蓿公共图谱,选取8对染色体上分布基本均匀的66个SSR标记进行基因分型。SSR引物由上海生工生物技术有限公司合成,10 μL PCR体系中含有DNA 25 ng, Taq酶0.6 U,上下游引物各3 pmol, dNTPs 0.6 mmol,Mg2+20 pmol和10 × buffer 1 μL。扩增程序先94 ℃预变性5 min;随后94 ℃ 40 s、55 ℃ 50 s、72 ℃ 50 s进行30个循环;最后72 ℃延伸8 min。扩增产物经8%聚丙烯酰胺凝胶电泳,银染显色检测。
1.7 数据分析
表型数据用Excel和SPSS 17.0进行方差、新复极差检验和聚类分析获得相似性和进化树。
SSR标记带型通过人工统计,基因型数据用Powemarker软件分析各品系之一的遗传相似性系数[15],用非加权配对法(UPGMA)进行聚类分析,构建发育进化树状图。
2 结果与分析
2.1 产量相关性状影响因素分析
方差分析结果表明,苜蓿鲜重、株高和主茎节数受到年份、品系、茬次3个因素、以及年份与茬次的交互作用的显著影响(P< 0.05),主茎节数还受到品种与茬次交互作用显著影响(P< 0.05),其他的交互作用对3个性状均无显著影响(P> 0.05)。鲜重最高的品种是‘WL363HQ’,其次依次是‘BaRaH416wet’、‘WL298HQ’和‘巨能551’,共23个品种与‘WL363HQ’间无显著差异(P> 0.05) (表1)。黄花苜蓿的主茎节数和株高最高,其中主茎节数显著高于其他品种(P<0.05)。
不同茬次的苜蓿产量之间都有显著性差异,并随茬次推移,产量逐渐降低,第1茬产量最高(表2)。第2年平均产量是第1年的2.24倍,第2年第1茬产量更是第1年第1茬产量的6.74倍。不同茬次株高和主茎节数都是第3茬最高,但产量没有随株高和主茎节数增加而增加,株高和主茎节数对产量没有影响,可能分蘖数才是影响产量的主要因素。
2.2 品质相关性状影响因素分析
经方差分析表明,CP、ADF、NDF含量和RFV都受到品系、年份和茬次的显著影响(P< 0.05),而EE含量受到品系和茬次显著影响(P< 0.05),不受年份的影响。年份与品系交互作用显著影响EE、ADF、NDF含量和RFV (P< 0.05),年份与品种交互作用显著影响CP、EE、ADF、NDF含量和RFV (P<0.05),品种与茬次交互作用显著影响CP和ADF含量(P< 0.05),三者的交互作用显著影响CP、ADF和NDF含量(P< 0.05)。苜蓿CP含量变异幅度较小(表3),变异范围为21.09% ('WL440HQ')~23.54% (‘WL354HQ’),并且‘WL358HQ’显著高于‘WL363HQ’、‘WL712’、‘三德利’和‘WL440HQ’ (P<0.05),与其余品系没有显著差异(P> 0.05)。EE含量最高的是‘四季旺’(6.58%),并显著高于‘WL712’、‘WL366HQ’和‘巨 能995’ (P< 0.05)。RFV以‘WL354HQ’最 高,并 显 著 高 于‘WL656HQ’、‘WL525HQ’、‘巨能601’、‘BaRaH416wet’和黄花苜蓿(P< 0.05),而黄花苜蓿最低,显著低于其余品种(P< 0.05)。
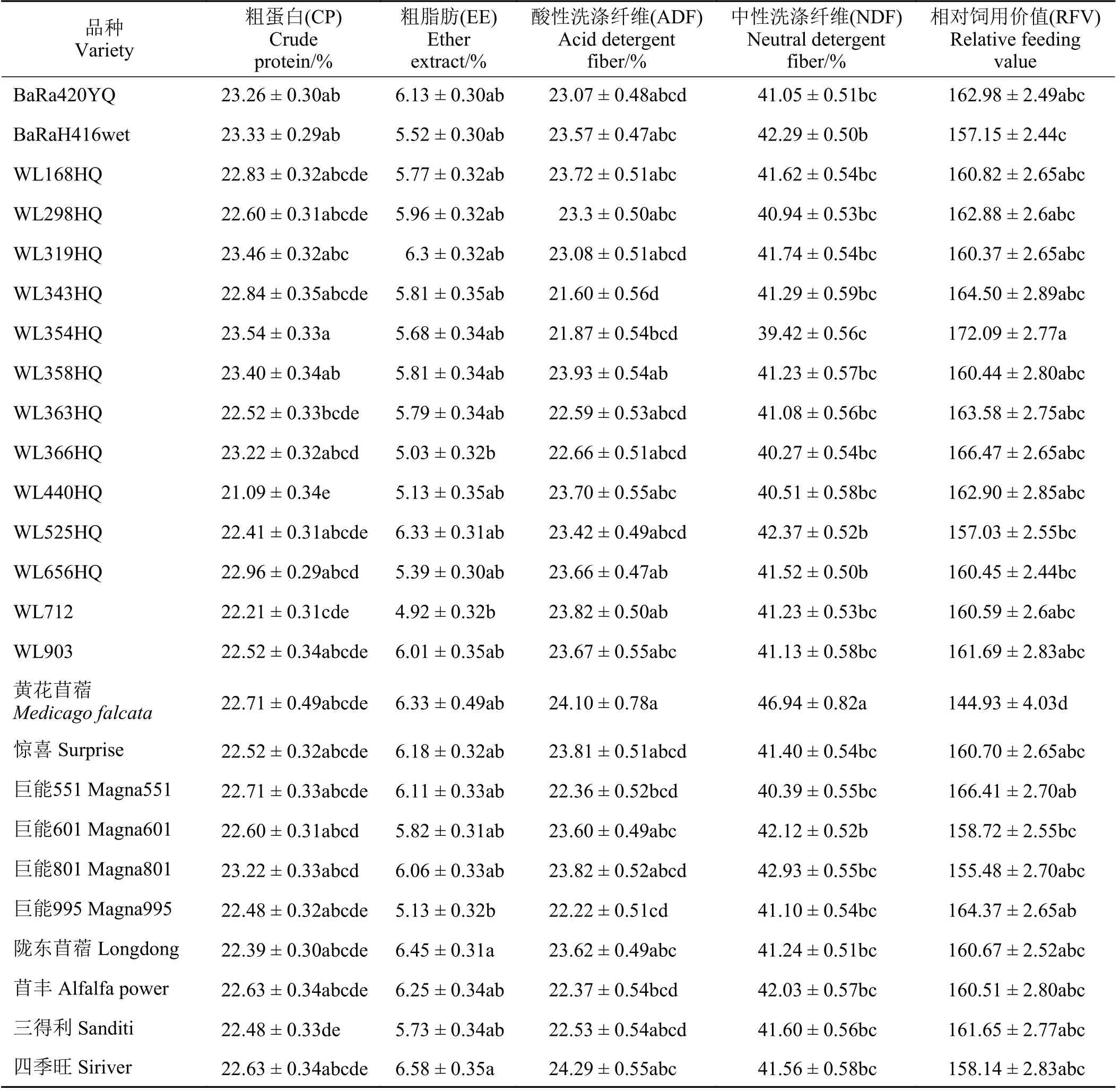
表3 不同品种对苜蓿品质的影响Table 3 Effect of different varieties on quality traits
第1茬的CP和EE含量都最高,并显著高于其余4个茬次(P< 0.05) (表4),第2和4茬的CP和EE含量最低,两茬之间没有显著差异(P> 0.05)。RFV不同茬次之间都存在显著差异(P< 0.05),并且第2茬最高,其次是第1茬,第4茬最低。从品质性状总体来看,第1茬草品质最好。

表4 茬次对产量相关性状的影响Table 4 Quality-related traits of different batches
2.3 苜蓿品系间亲缘关系分析
用平均欧式距离和组间连接法将表型数据间进行聚类分析,结果表明所选25个苜蓿品系被聚为3个群,黄花苜蓿被单独聚为一类,与其他苜蓿的遗传距离较远;BaRaH416wet、WL298HQ、WL343HQ、WL363HQ和WL903这5个品种被聚为一类,其余19个品种被聚为一类(表5)。黄花苜蓿与WL363HQ遗传距离最远,BaRa420YQ与WL319HQ遗传距离最小(图1A、附表2)。
利用66对在苜蓿8个连锁群中分布均匀的SSR标记聚类分析(图1B、附表3),结果表明,25个品系被分为两类,‘BaRa420YQ’、‘BaRaH416wet’、‘WL168HQ’、 ‘WL298HQ’、 ‘WL319HQ’、‘WL343HQ’和‘WL354HQ’共7个品系被聚为一类,其余18个品系聚为另一类(表5)。‘苜丰’和‘WL298HQ’之间得遗传距离最大,‘巨能601’与‘巨能801’之间的遗传距离最小。表型和基因型聚类结果具有一定的差异,说明在育种中亲本选配上既要考虑表型差异,也要从基因型上进行选择。

表5 表型和基因型聚类结果Table 5 Results of phenotypic and genotype clustering
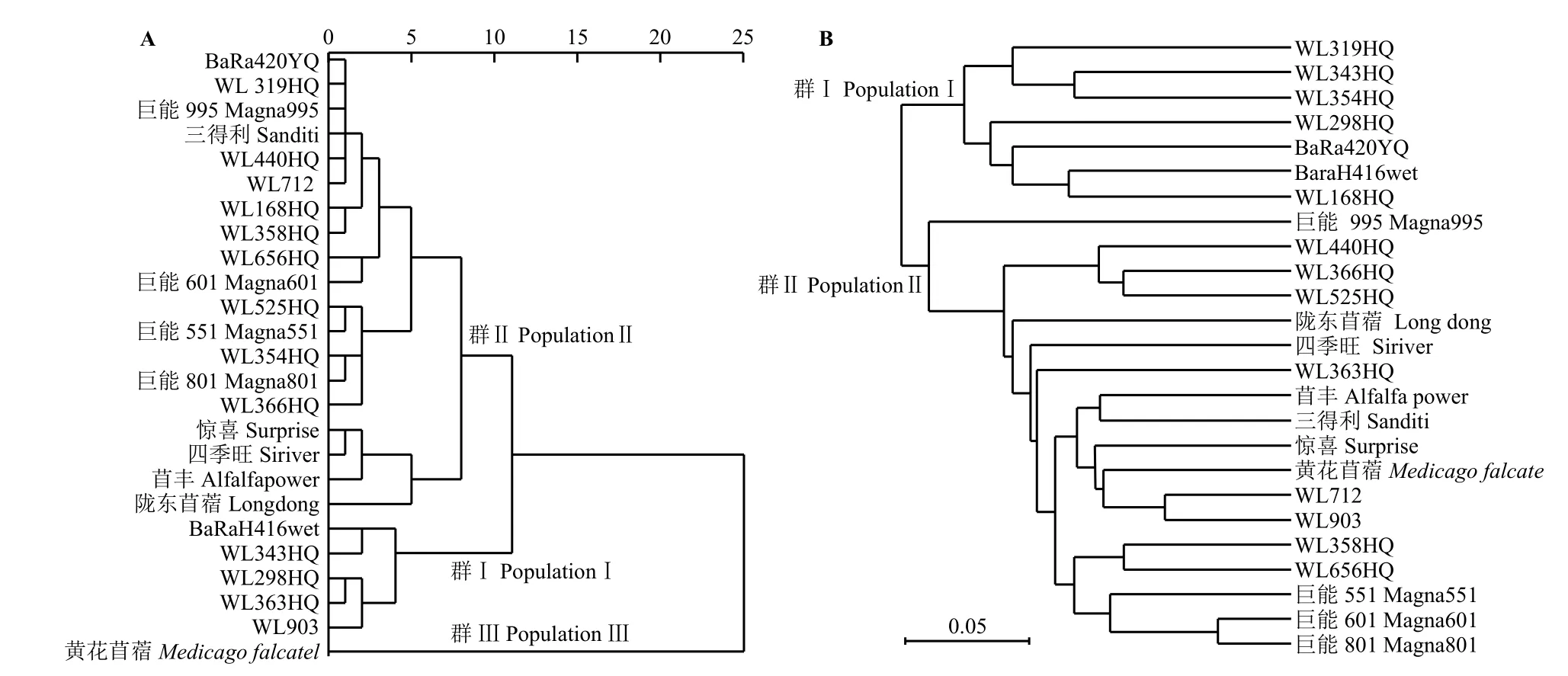
图1 25个苜蓿品系聚类分析图Figure 1 Cluster diagram of 25 alfalfa varieties
3 讨论与结论
3.1 淮河中下游地区苜蓿品种的适应性评价
选择适合淮河中下游地区种植的苜蓿品种,对促进该地区草业的发展具有重要意义。试验结果表明 半 休 眠 的‘WL363HQ’、‘BaRaH416wet’、‘WL298HQ’和‘巨能551’都有较高的产量,并且‘BaRaH416wet’、‘WL298HQ’和‘巨能551’具有较高的蛋白质和粗脂肪含量,与最高蛋白含量的品种‘WL358HQ’之间没有显著差异,它们的相对饲用价值也较高。其余半休眠品种其产量较高,因此淮河中下游地区苜蓿种植应选择半休眠品种。但产量与株高及主茎节数之间没有相关性,可能影响苜蓿的产量主要因素是分蘖数,生产中通过促进苜蓿分蘖达到高产的目标。
淮河中下游地区苜蓿种植第2年产量是第1年的2倍以上,因此苜蓿该地区苜蓿种植应多年利用,否则难以获得较好经济效益。第1年产量第2茬最高,第2年第1茬产量最高,前2茬产量占了全年总产量50%以上,并且其蛋白质、粗脂肪含量和相对饲用价值也是第1和2茬较高,这可能是该地区第3茬以后气温过高,影响了苜蓿生长。
3.2 苜蓿表型和基因型之间的亲缘关系
剖析不同种质资源的表型组和基因组之间的关联程度为育种中的亲本选配提供指导,特别是在育种过程中将地方种质资源中的优异基因导入优异品种达到改良品种的目标,具有重要价值。本研究收集了当地的黄花苜蓿资源,其具有更多的主茎节数和更高的株高等优良特性[16],是优异的地方资源,但产量相对较低,通过杂交回交等将优良基因导入高产优质苜蓿品种中,具有十分重要的意义。基因型和表型数据聚类并不一致,这可能是因为苜蓿是同源异化授粉的四倍体,其表型受基因型控制比二倍体更复杂,不但受到基因位点的显性、上位性和共显性等的影响,同时还受到基因剂量的影响,SSR标记所做的基因型分析并不能体现基因剂量,可能会导致基因型和表型的不一致,但基因型之间能够反映基因组之间的遗传距离关系[17]。因此仅仅通过表型判断苜蓿之间的遗传关系是不准确,亲本选配时,需要表型上选择综合性状优良,差异互补,基因型差异大的品系作为杂交亲本,在后代中更容易选择出优良品种,本研究能够为淮河中下游地区苜蓿品种选育亲本选配提供指导。
