槭叶铁线莲(Clematis acerifolia Maxim.)群落物种生态位研究
2022-05-20庞久帅原阳晨周苗苗任俊杰成克武
庞久帅,原阳晨,2,周苗苗,任俊杰,高 鹏,成克武
1 河北省洪崖山国有林场,保定 071000 2 河北农业大学,保定 071000
生态位是指生物个体或群体所能利用的各种资源的总和及其与其他生物之间的功能关系[1—2],生态位宽度、生态位重叠和生态位相似性是描述物种生态位特征的主要指标,可以量化物种间的竞争关系和物种对环境中资源的利用能力,因此,生态位分析已成为评价种间关系和物种在群落中功能地位的重要方法[3],对于揭示群落内种间关系、群落结构功能、群落动态演替、生物多样性和种群进化等方面具有重要意义[4]。自1917年Grinnell首次提出生态位概念[5]以来,国内外学者对生态位的相关理论及应用方面进行了深入探究,使其成为生态学研究的热点领域之一。目前,国内外对生态位的研究主要集中在生态位大小测度[6]、物种共存[7]、生态位与生境关系[8—9]、物种适应性策略[10]方面,对珍稀濒危物种在群落中的生态位及其生存安全性方面的研究相对不足。
槭叶铁线莲(Clematisacerifolia)是毛茛科铁线莲属多年生灌木植物,花大美丽,花期早,是早春极为珍贵的观赏花卉[11]。已知的槭叶铁线莲分布范围仅限于太行山北部,包括北京门头沟6处(担礼村担礼隧道旁登山古道、妙峰山、担礼村前山、担礼村担礼遂道旁、百花山柳林水附近、丁家滩),房山区9处(上房山接待奄、六渡、霞云岭水库、霞云岭公社四合村、十渡镇王老铺村、十渡镇平峪村、四合村、贾峪口村、河北省镇),河北涞水县2处(野三坡、杨家坪老坡岭),在昌平区也发现该植物,均生长于石灰岩山地的悬崖峭壁上,多为零星分布,种群数量通常在几株到数十株之间,同时由于人为因素和美国地锦(Parthenocissusquinquefolia(L.)Planch)的威胁,至今发现的槭叶铁线莲均处于濒危状态[11—13]。槭叶铁线莲花期极早、数量稀少、生境独特,已被列入为《北京市一级保护野生植物名录》[14]。目前对槭叶铁线莲的研究还很少,穆林[15]等对其属内分类地位进行了探讨,认为该植物系统位置孤立,为铁线莲属在北温带古老类群的孑遗,其他学者对槭叶铁线莲遗传多样性和遗传结构[16]、个体与种群水平上杂合性和适应性[17]进行研究,关于槭叶铁线莲生态位方面的介绍还处于空白。本研究在野外群落学调查的基础上,以物种重要值作为生态位计量指标,对新发现的分布于河北省易县葫芦峪沟谷两侧崖壁上的槭叶铁线莲群落中物种的生态位进行研究,以期为槭叶铁线莲的有效保护和合理利用提供理论依据。
1 研究区概况
研究区位于河北省保定市易县流井乡建新村林区,地理坐标39°28′23.47″— 39°32′21.34″N,115°27′19.98″—115°26′34.57″E,属太行山北端东麓低山区域。该区属于暖温带大陆性季风气候类型,四季分明,春季多风少雨,夏季高温多雨,秋季干燥气爽,冬季低温少雪。平原地区多年平均气温为13.5℃,山区多年平均气温为9.4℃。全年7月份气温最高,1月份气温最低,极端情况下,气温最高为41.6℃,最低为-21.4℃。全年降水分配不匀,7月、8月最多,年均降水为544.8 mm。研究区基岩为石灰岩,地貌主要由葫芦峪主沟及其两侧支沟构成,主沟由南向北弯曲延伸,长约5 km,呈峡谷状,两侧为山脊,坡度陡峭,中下部多为垂直崖壁,崖壁呈不连续状分布,是槭叶铁线莲的主要生长环境。从主沟入口端至沟谷末端沟底海拔范围为120—530 m。
2 研究方法
2.1 样地选择及调查
槭叶铁线莲在葫芦峪沟谷悬崖峭壁上呈斑块状分布,从2020年3月下旬槭叶铁线莲开花开始,对其在葫芦峪分布范围及生境状况进行初步调查,2020年6月上旬采用典型取样方法,选择7块样地对槭叶铁线莲群落进行调查。样方设置采用1 m×1 m网格法,调查各网格(小样方)中出现的植物物种及个体数、频度和盖度,同时记录样地内的海拔、坡向、坡度、坡位和植物总盖度等因子。各样地取样面积分别为(Ⅰ)33 m2、(Ⅱ)100 m2、(Ⅲ)50 m2、(Ⅳ)25 m2、(Ⅴ)50 m2、(Ⅵ)25 m2、(Ⅶ)9 m2。样地基本概况见表1:
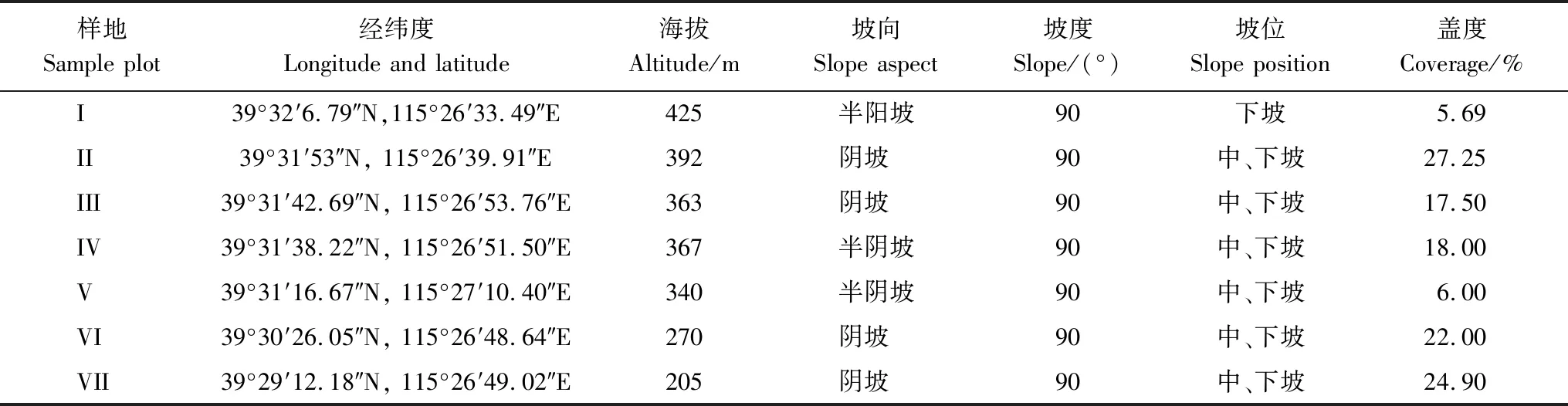
表1 样地基本概况Table 1 Basic conditions of sample zones
2.2 生态位指标及研究方法
各个样地作为不同的生态位资源状态,通过各植物种的重要值,对生态位宽度、生态位重叠和生态位相似性比例进行计测和分析[18],相关计算方法如下:
2.2.1重要值测定
重要值能够反映出群落中的优势物种,可以客观的表达出各物种在群落中的地位和作用,其计算方法[19—23]为:
重要值=(相对密度+相对盖度+相对频度)/3
相对密度=某物种株数/所有物种株数和×100%
相对盖度=某物种盖度/所有物种盖度和×100%
相对频度=某物种频度/所有物种频度和×100%
2.2.2生态位宽度测定
采用 Levins(1968)[24—25]指数测定各物种的生态为宽度,计算公式为:
式中,i表示物种,j表示第几个资源位(样地),r表示资源位数(样地数)。Bi为物种i的生态位宽度,Pij为物种i在第j个资源位中的重要值与所有资源位中物种i重要值之和的比例,Pij=nij/Ni,nij为物种i在第j资源位中的重要值,Ni=∑nij,∑nij为物种i在所有资源位中重要值之和。
2.2.3生态位重叠
采用Pianka[26—31]指数测定物种间的生态位重叠,计算公式为:
式中,Qik为种i与种k之间的生态位重叠程度,值域为[0,1];i与k分别表示两个不同的物种,j表示第几个资源位(样地),r为资源位数(样地数)。Pij与Pkj分别为物种i与物种k在资源位j的重要值与所有资源位中物种i与物种k重要值之和的比例,Pij与Pkj计算方法相同。Pkj与Pij计算方式同生态位宽度。
2.2.4生态位相似性比例
生态位相似性比例指2个物种利用资源的相似性程度,其计算公式[32]为:
式中,Cik表示物种i与物种k的生态位相似性比例,具有数值范围[0,1];公式中其他字母含义以及Pij与Pkj的计算方式同生态位重叠。
2.3 数据分析
在Excel 2013中整理调查数据,并计算槭叶铁线莲群落中物种的重要值、生态位宽度值、生态位重叠值和生态位相似性比例值。
3 结果与分析
3.1 物种组成及重要值
重要值是表征物种在群落中的地位和作用的一个综合性数量指标,也能够反映物种在群落中的优势程度[33]。根据调查统计(表2),槭叶铁线莲群落7个样地中共包含17个植物物种,分属于14科15属,包括12种
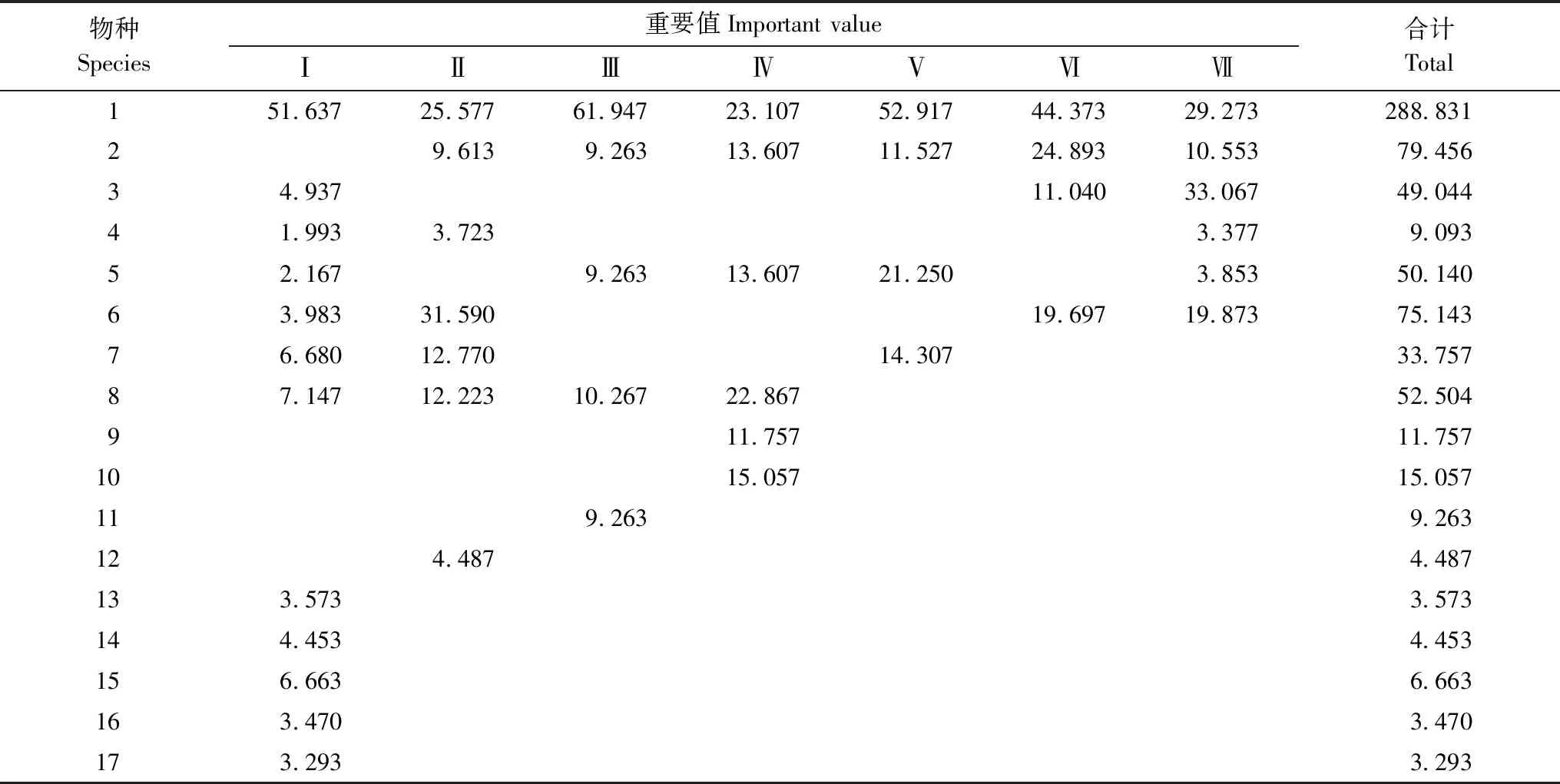
表2 槭叶铁线莲群落各物种重要值Table 2 Improtant value of each species in Clematis acerifolia community
以下各表物种和样地编号不变species and sample plot number sequence not change in thereinafter;1.槭叶铁线莲Clematisacerifolia;2.绒毛绣线菊Spiraeadasyantha;3.太行藨草Scirpusschansiensis;4.薄皮木Leptodermisoblonga;5.荆条Vitexnegundovar.hetrophyla;6.独根草Oresitropherupifraga;7.侧柏Platycladusorientalis;8.黄栌Cotinuscoggygriavar.cinerea;9.本氏木兰Indigoferabungeana;10.小叶鼠李Rhamnusparvifolia;11.雀儿舌头Leptopuschinensis;12.小叶白蜡Fraxinussogdiana;13.鼠李RhamnusdavuricaPall.;14.锐齿鼠李RhamnusargutaMaxim;15.抱茎苦荬菜Ixelrissonchifolia;16.紫花地丁Violayedoensis;17.斑种草Bothriospermumchinens。Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ和Ⅶ分别为槭叶铁线莲群落7个样地编号
乔灌木植物和5种草本植物,群落物种组成较为简单。槭叶铁线莲重要值在5个样地中位居首位,在其他2个样地中居第二位,在7个样地中重要值总和为288.831,占所有物种重要值的41.26%,7个样地中其他物种各自的重要值之和均低于槭叶铁线莲,且各样地中重要值排序也不一致,反映出槭叶铁线莲群落不同样地在物种组成和数量分布上存在着差异,槭叶铁线莲在群落中总体上占绝对优势地位。
3.2 生态位宽度
生态位宽度是指植物群落中物种利用生境资源的总和,可以度量各物种对环境资源的利用情况,物种生态位宽度大,对环境具有较强的适应能力,可以充分利用环境资源[34]。槭叶铁线莲群落中各物种的生态位宽度(Bi)见表3,群落内17个物种中,槭叶铁线莲生态位宽度最大,为6.264;其次是绒毛绣线菊,生态位宽度值为5.144;荆条(Vitexnegundovar.hetrophyla)、黄栌(Cotinuscoggygriavar.cinerea)、独根草(Oresitropherupifraga)、薄皮木(Leptodermisoblonga)和侧柏(Platycladusorientalis)的生态位宽度处于中等水平,分别为3.388、3.326、3.143、2.828和2.763;太行藨草(Scirpusschansiensis)生态位宽度较窄,为1.940;其他物种生态位宽度值均为1,宽度值最小,他们在所有调查的样地中仅出现1次。
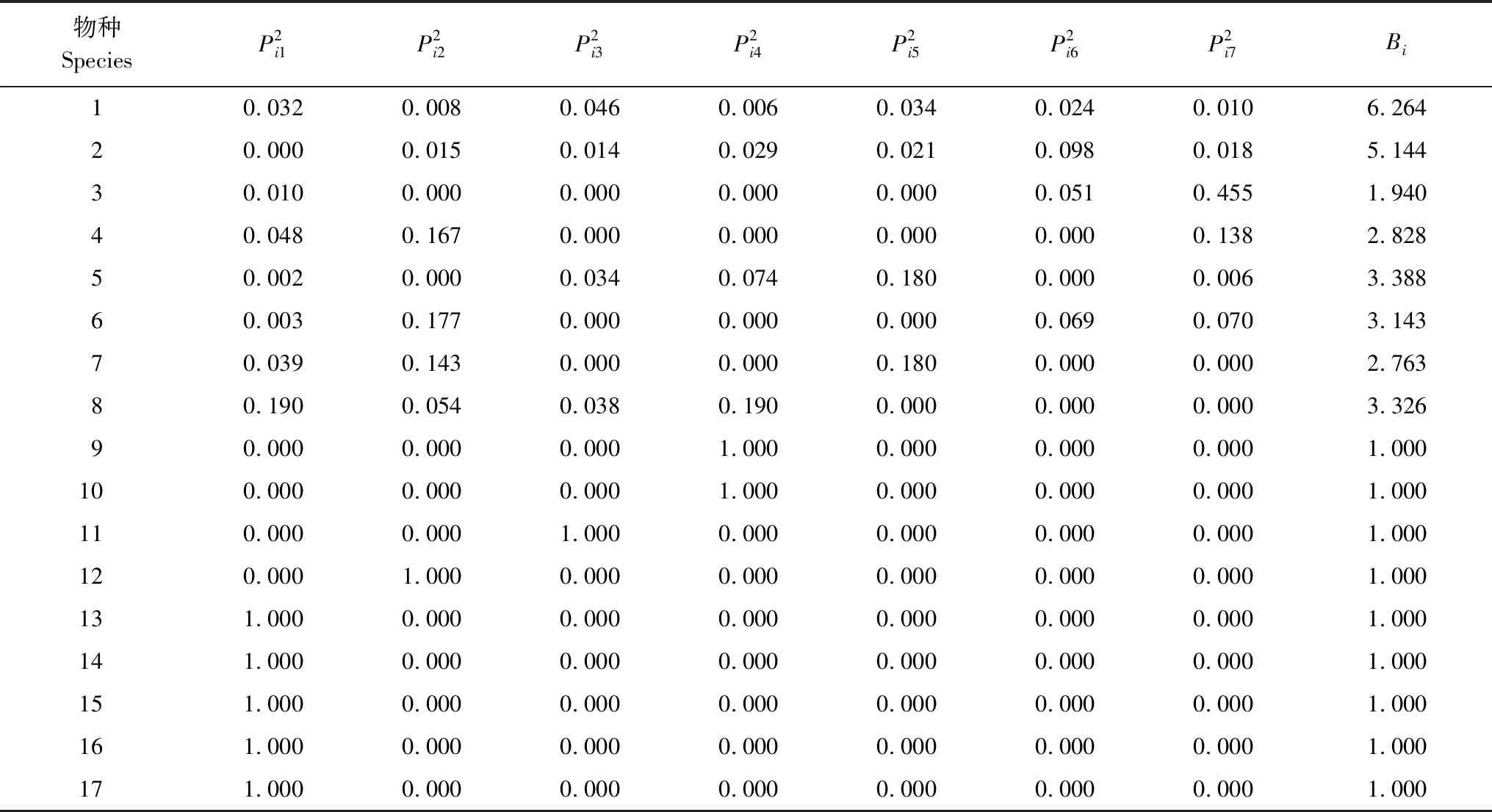
表3 槭叶铁线莲群落各物种生态位宽度Table 3 Niche breadth of each species in Clematis acerifolia community
物种的生态位宽度越大,其特化的程度就越小,越趋于泛化种,反之为特化种[35]。泛化种在资源有限的情况下,具有较强的竞争能力,而特化种竞争能力较弱,生态位较窄[36]。从生态位宽度计算结果可以反映出群落中槭叶铁线莲竞争能力和适应性强,本氏木兰(Indigoferabungeana)、小叶鼠李(Rhamnusparvifolia)等多数物种生态位宽度小,在此环境下趋于特化种。
3.3 生态位重叠程度
槭叶铁线莲群落各物种的生态位重叠情况见表4,由群落内17个物种组成的136个种对中,完全重叠(Qik=1)的种对有11个,占总对数的8.09%;生态位重叠程度较高(0.5≤Qik<1)的种对有23个,占总对数的16.91%;生态位重叠程度较低(0 表4 槭叶铁线莲群落各物种生态位重叠Table 4 Niche overlap value of each species in Clematis acerifolia community 群落中槭叶铁线莲与其他所有物种之间生态位都存在不同程度的重叠,与其他物种形成的16个种对中10个种对的生态位重叠程度较低(0 生态位相似性可以反映两个物种利用资源的相似性程度。槭叶铁线莲群落各物种生态位相似性比例情况见表5,17个物种组成的136个种对中,生态位完全相似(Cik=1)的种对有11个,占总对数的8.09%;生态位相似性比例较高(0.5≤Cik<1)的种对有8个,占总对数的5.88%;生态位相似性比例较低(0 表5 槭叶铁线莲群落各物种生态位相似性比列Table 5 Niche proportional similarity of each species in Clematis acerifolia community 群落中槭叶铁线莲与其他所有物种之间生态位相似性不同,与其他物种形成的16个种对中13个种对的生态位相似性程度较低(0 重要值和生态位宽度都可以反映物种在植物群落中的地位和作用,但两者具有不同的生态学意义,重要值体现物种在植物群落中的相对重要性和优势程度,而生态位宽度反映物种对生境资源的利用能力和对环境适应性等[4]。槭叶铁线莲重要值与生态位宽度在群落中均居首位,分别为288.831和6.264,反映出在葫芦峪沟谷崖壁环境中其生态适应幅度相对较广,利用资源能力也较强,处于群落中的优势地位,在群落演替中处于较为安全的地位。其他物种在不同样地中重要值排序各不相同,生态位宽度低于槭叶铁线莲。 槭叶铁线莲群落中大部分物种重要值与生态位宽度呈正相关关系,但部分物种例外,如太行藨草重要值大于薄皮木,而生态位宽度却比薄皮木小,小叶鼠李的重要值也大于薄皮木,生态位宽度却也小于薄皮木,反映出重要值不是决定生态位宽度的唯一因素,相关研究表明,这一现象与物种的分布频度[4]或所处的异质性生境[24]有关。本研究中,物种生态位宽度除与重要值密切相关外,与其分布的资源位数量具有紧密关联,生态位宽度较大的槭叶铁线莲和绒毛绣线菊分布资源位数量多,生态位宽度最小的本氏木兰、小叶鼠李和雀儿舌头仅出现于一个资源位,这与郑晓阳、钱逸凡等[37—38]研究结论相符。 生态位重叠是反映同一空间中物种对生境资源利用差异以及物种间竞争共同资源的重要指标,生态位重叠程度越高说明物种间对生境资源利用越相似,可能发生激烈竞争[39]。通常物种生态位越宽,对资源利用能力就越强,分布范围也越广,与群落中其他植物的重叠程度也较大[40],本研究中生态位宽度较大的槭叶铁线莲、绒毛绣线菊、独根草,他们之间所构成的种对间的生态位重叠值也较大,符合此规律。同时,本研究中也发现个别生态位宽度大(小)的物种间会出现生态位重叠幅度较低(高)的情况,如荆条和独根草各自生态位宽度较大,但种对生态位重叠程度很小,太行藨草和薄皮木生态位宽度较小,但种对生态位重叠程度却很大,这种情况首先与槭叶铁线莲群落的环境分异有关。槭叶铁线莲虽分布于垂直崖壁上,但不同坡向崖壁的水分条件和光照条件存在差异,导致生态习性不同的物种出现在不同样地中,尽管各自在分布样地中占据较大生态位,但其他物种间共同出现的频率较低,生态位重叠程度低;即便是在同一样地中,岩壁上也会因裂隙变化不同、雨水补给差异或岩石松动掉落形成微环境和资源的空间异质性,使不同物种呈现斑块状分布,降低生态位重叠程度[41];其次与其他物种的侵入和定居存在一定的随机性有关,由于槭叶铁线莲分布的崖壁生境严峻,植物生存所需资源匮乏,入侵和定居较为困难,根据竞争抽彩理论[42],物种的入侵存在随机性,先入侵物种可限制其他物种侵入,导致部分物种出现频率较高,部分种类则很低,甚至仅出现1次或没有机会出现。以上原因都会影响个别物种的分布不完全遵循生态位重叠的基本理论。 生态位相似性是衡量物种间资源利用相似程度的重要指标,可以利用其判断物种间是否存在竞争以及群落是否稳定[39]。整体上槭叶铁线莲群落中各物种间的生态位相似性程度较低,表明物种间竞争不激烈,群落较为稳定,其原因一方面与生境的分异和物种对生境的需求差异有关,另一方面槭叶铁线莲生长的崖壁土壤养分和水分缺乏,环境极为严酷,难以满足更多物种生存所需的资源,物种的生存更趋向于Grime的CSR理论中的胁迫-忍耐对策(S选择)[43]。槭叶铁线莲与群落中绒毛绣线菊的生态位相似性程度较高,它们的生活习性相近,对环境资源需求相似,因而共同出现的频率较高,但与其他种类相似性较低,竞争较低。研究表明,生态位相似性与其生态位宽度呈正相关[44],本研究中大部分种对间均成正相关,如生态位宽度大的槭叶铁线莲-绒毛绣线菊种对的生态位相似性达0.685,生态位宽度窄的本氏木兰-雀儿舌头种对生态位相似性为0。但本研究也出现生态位宽度窄而生态位相似性大的种对,如本氏木兰-小叶鼠李种对和抱茎苦荬菜(Ixelrissonchifolia)-紫花地丁(Violayedoensis)种对,主要原因也与各样地的环境差异性以及物种生态特性不同有关,槭叶铁线莲群落分布于不同坡向的垂直崖壁上,崖壁缝隙提供的水分、养分较为缺乏且不均匀,导致物种分布受生境和生态习性影响,同时个别物种的分布也存在一定的偶然性,如在调查的样地中,个别阳坡样方中的崖壁岩石碎裂脱落后形成积土小平台,导致喜光、适应性强的一年生草本植物(抱茎苦荬菜、紫花地丁和斑种草Bothriospermumchinens)会偶然出现,而通常情况下它们仅分布于土层较厚、水分条件较好的环境,在崖壁上难以生存,这种个别物种出现在某一特定小环境中的情况,造成其物种生态位宽度窄,但种对间的生态位相似性较高的情况出现。 生态位分析能揭示群落中不同物种利用资源和空间的能力以及物种间的共存与竞争关系,可以为珍稀濒危物种如槭叶铁线莲的生存状态分析和未来的安全风险评价提供理论依据,但目前在依据此理论进行有关珍稀濒危物种安全风险分析的研究还较为缺乏,本研究通过对槭叶铁线莲群落物种生态位分析,为濒危物种槭叶铁线莲的生存安全状况评价提供了一种方法途径,有助于其保护和管理策略的制订。 槭叶铁线莲群落中各物种之间的生态位重叠程度较小,生态位相似性较低,物种间在资源利用上存在一定差异、相互竞争不是很激烈,群落结构较为稳定,槭叶铁线莲在群落中的的重要值和生态位宽度最大,占绝对优势,所受到的竞争作用相对较小,处于较为安全的状况。槭叶铁线莲群落生境特殊,在葫芦峪沟谷主要分布于悬崖峭壁上,绝大部分生境处于人为无法触及的地方,人为干扰小,基于严酷的环境条件,物种间自然竞争强度较小,槭叶铁线莲种群整体上能够安全生存,仅沟谷底部局部地段的槭叶铁线莲个体会受到游人活动或放牧影响,需加强就地保护[16],维持现有的栖息地环境[17],来保证其安全生存,可在受游人或放牧影响的区域,通过修建围栏、挂立警示标牌等管理措施,以实现槭叶铁线莲的保护与管理。通过对槭叶铁线莲在葫芦峪沟谷的分布数量和年龄结构方面的调查发现,葫芦峪沟谷有35个槭叶铁线莲集群,个体数量达3166丛,阴坡和阳坡均有其分布,阴坡幼年个体和成熟阶段前期个体比例占优,集群处于增长阶段,阳坡各阶段组成比例接近,集群处于较为稳定状态,年龄结构也表明葫芦峪沟谷槭叶铁线莲种群较为安全[45]。鉴于目前对槭叶铁线莲生理、生态习性方面研究还较为缺乏,今后应加深对其生物学、生理生态学和遗传学等方面的研究。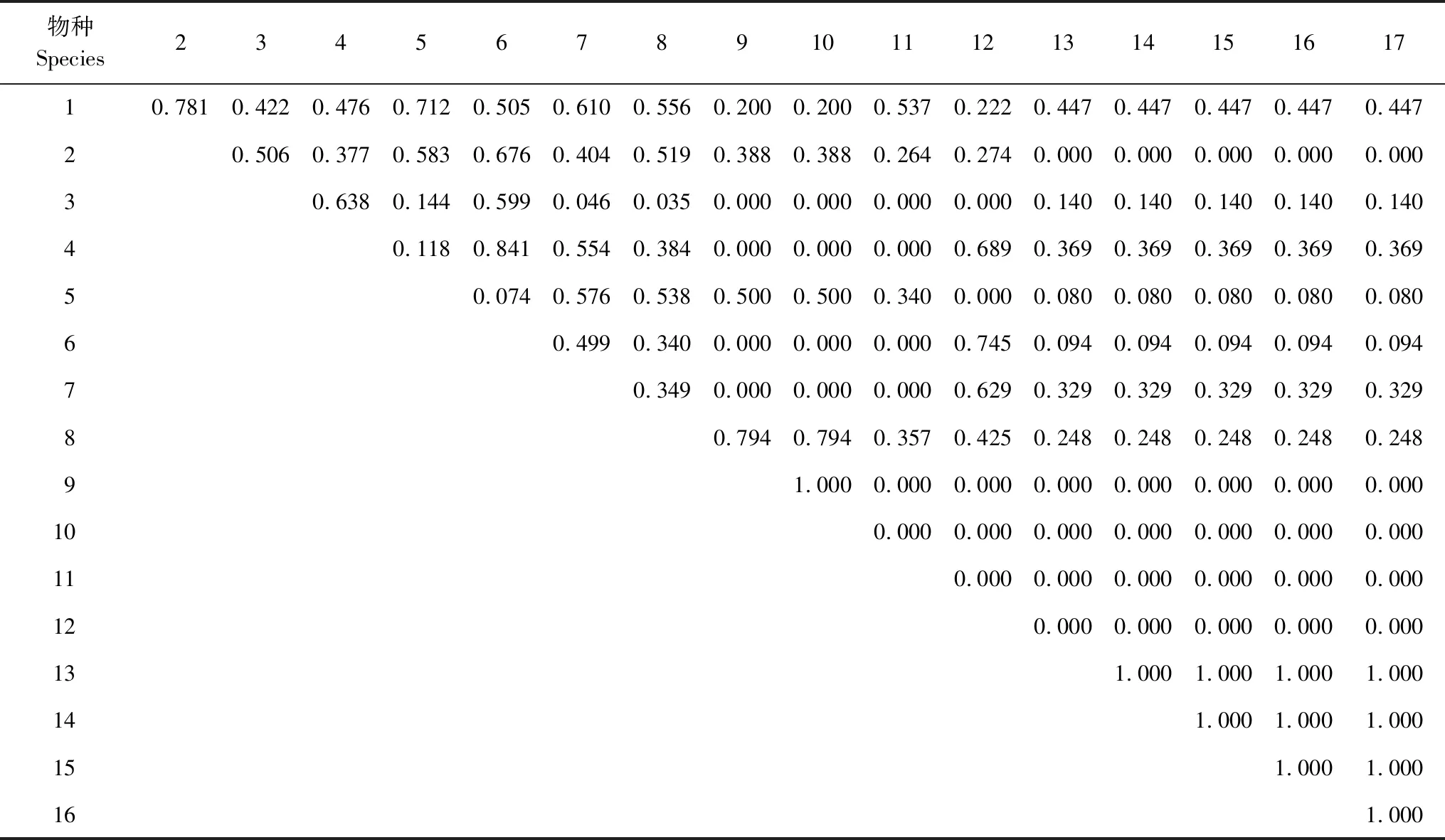
3.4 生态位相似性比例
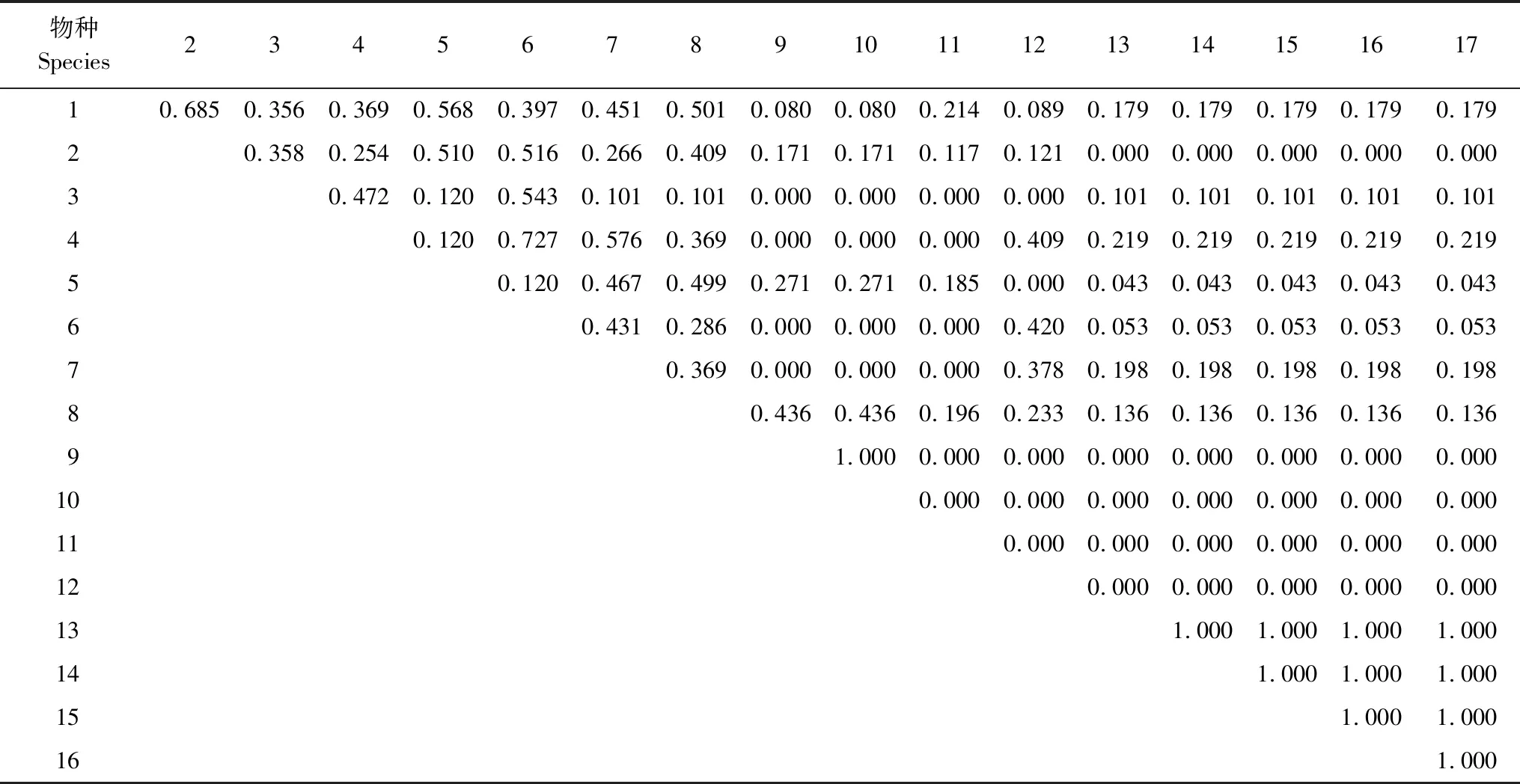
4 讨论
4.1 槭叶铁线莲群落生态位宽度特征
4.2 槭叶铁线莲群落生态位重叠特征
4.3 槭叶铁线莲群落相似性特征
5 结论
