芝麻细胞色素P450超家族的成员鉴定及SiCYP703A2的功能验证
2022-05-13李田雨杨远霄刘红艳汪魏周芳周婷赵应忠
李田雨,杨远霄,刘红艳,汪魏,周芳,周婷,赵应忠
(中国农业科学院油料作物研究所,农业 农村部油料作物生物学与遗传育种重点实验室,湖北 武汉, 430062)
芝麻是我国的传统特色油料作物,具有极高的营养价值和经济价值。我国虽是世界上芝麻的主产国之一,但也是最大的芝麻进口国,芝麻原料供给严重不足[1]。利用杂种优势可以显著提高作物产量,增强植株抗逆性和抗病性,是可持续发展条件下增加作物产量的有效途径。目前,在水稻、油菜、小麦等多种作物中已建立杂优育种体系,显著提高了作物产量[2~5]。植物雄性不育作为杂种优势利用的最优途径而受到广泛研究,而挖掘雄性不育相关基因,对于通过基因工程创制雄性不育材料、选配优势杂交组合、提高芝麻产量具有重要的意义。
细胞色素P450(cytochrome P450,简称CYP450)是植物中最大的代谢酶家族,具有广泛的底物谱和催化活性,催化大量初生和次生代谢物的生物合成及外源毒性化学物质的降解[6]。基于系统发育研究,Nelson[7]将陆地植物P450 超家族归类为10 个独立的家族簇(clan),即源于相同祖先的不同基因家族的集合,同时对双子叶植物和单子叶植物中各个家族簇中家族分布情况做出阐述。每个家族簇(clan)以簇中最小蛋白家族的编号作为簇名,包括71 Clan、72 Clan、85 Clan、86 Clan、51 Clan、74 Clan、97 Clan、710 Clan、711 Clan 和727 Clan,其中727 Clan为单子叶植物特有[8,9]。家族和亚家族基因的分类与命名主要依据基因编码的氨基酸序列之间的相似度[10]。目前在植物基因组中共鉴定出74 个P450 家族(family),包括127 个不同的亚家族(subfamily)[11]。
随着分离和鉴定突变体技术的发展,不断有新的P450 基因功能被发掘。目前有多个家族,近40个P450基因被克隆并鉴定参与了脂肪酸的羟基化、环 氧 化 等,包 括 家 族CYP703、CYP704、CYP92、CYP81、CYP77、CYP78、CYP96、CYP709、CYP726、CYP86 和CYP94[12]。其中拟南芥CYP704B1是一种长链脂肪酸v-羟化酶基因,对花粉中孢粉素的合成至关重要[13]。有研究表明牵牛花CYP703A1在花药发育早期阶段表达,并催化月桂酸的羟基化[14]。拟南芥CYP703A2在花药小孢子和绒毡层细胞表达,催化中链饱和脂肪酸转化为相应的单羟基脂肪酸,被鉴定为是一个特异参与花粉发育的P450 家族[15]。另外,水稻CYP703A3作用于花药角质层和花粉外壁的发育,参与雄性器官发育所需的月桂酸链内羟化途径[16,17]。除了脂肪酸外,CYP450 还主要参与生物体内萜类化合物的合成,菊科植物黄花蒿的CYP71AV1作为多功能倍半萜类氧化酶基因在青蒿素的生物合成途径中起关键作用[18]。在唇形科植物丹参中,发现CYP71BE52氧化铁氨酚,产生了二萜类物质丹参酚[19]。
本研究鉴定了芝麻P450超基因家族的成员,并克隆了芝麻CYP703 家族成员SiCYP703A2基因,初步揭示了其在花药发育中的功能。SiCYP703A2在芝麻花蕾中特异表达,且在花药发育的四分体时期高量表达。在拟南芥中超量表达SiCYP703A2导致转基因拟南芥雄性不育,花粉壁发育异常。研究的结果期待为芝麻花药发育和杂种优势利用提供有效的基因资源。
1 材料和方法
1.1 材料
芝麻材料为隐性细胞核雄性不育两用系95ms-5AB,种植于中国农业科学院油料作物研究所试验田,于自然条件下正常生长。于盛花期,分别取芝麻根、茎、叶、花蕾和果荚组织,以及可育和不育花药的四分体时期、小孢子发育时期和成熟时期的花药样品,迅速冷冻于液氮中,用于后续试验。哥伦比亚生态型拟南芥(Col-0)种植于培养室中,温度为22℃,光照强度为130 µmol·m-2·s-1,光周期为光照培养16 h,黑暗培养8 h,湿度控制在60%~70%。在苗期时取转基因植株和野生型的幼嫩叶片,在花期时取不同花药发育时期(stage7-8、9-10、11-12、13-14)的花蕾,迅速冷冻于液氮中,所有样品保存在-80℃备用。
1.2 芝麻CYP450超家族的鉴定
芝麻全基因组氨基酸序列下载自Sinbase 数据库(http://www. ocri-genomics. org/Sinbase/)[20],使 用TBtools 软件将其建立一个本地数据库,以拟南芥P450基因的氨基酸序列作为查询(query),在本地数据库中进行BlastP(E-value = 0.001),初步筛选出芝麻P450候选基因序列,后将搜索比对得到的所有同源氨基酸序列提交至Pfam 数据库中的Batch sequence search 程序(E-value= 0.001)中进行结构域的分析,检验其是否含有P450结构域(PF00067),剔除重复的和错误序列,最终得到预测的芝麻P450氨基酸序列。使用EMBOOS 软件包的needleall 程序(Needleman-Wunsch 算法)对303 条芝麻P450 基因的氨基酸序列进行全局比对,得到序列之间的一致度和相似度,参数均为默认值[21]。
模式植物拟南芥的P450 基因的氨基酸序列下载自P450 数据库(http://drnelson. uthsc. edu/CytochromeP450.html),共273 条(不包括未分类基因)。将得到的芝麻和拟南芥P450氨基酸序列通过ClustalW 工具进行序列比对,比对结果进一步通过IQtree 软件构建芝麻和拟南芥P450 基因的系统进化树,采用最大似然法(maximum likelihood,ML),设置重复次数(bootstrap)为1000。将芝麻P450 基因氨基酸序列逐条提交到拟南芥数据库(https://www.arabidopsis. org/Blast/index. jsp)中进行BLASTP,同时结合在NCBI 中BLASTP 的结果,寻找与其一致性最高的序列。依据P450 基因国际命名标准对预测的芝麻P450 基因进行家族分类:两个P450 基因的氨基酸序列一致性(identity)在40%~55%之间则划分为同一家族,大于55%则归为同一亚家族[10]。
基因连锁图由R 包karyoploteR(1.16.0)绘制,具体方法参见Bernat(2019)[22]。参照其它基因家族研究中关于基因簇的鉴定标准,对芝麻所有P450基因进行基因簇的分析[23,24]。判定标准:(1)两个相邻的P450 基因的间距≤200 kb;(2)两个P450 基因之间间隔的非P450基因个数≤8。
1.3 SiCYP703A2基因的序列分析
利用在线软件ExPASy(http://web. expasy. org/protparam)ProtParam 预测SiCYP703A2 蛋白的分子量和理论等电点。运用NCBI 中CDD(conserved domain database)程序分析氨基酸序列的结构域。采用ClustalX 软件对芝麻SiCYP703A2和其它物种中CYP703 家族基因进行多重序列比较分析,序列下载自NCBI(http://www. ncbi. nlm. nih. gov/),Accession number 为:AT1G01280、XP_011093925.1、XP_003624793.1 和XP_015648492.1;系统进化树使用MEGA7.0 软件的邻位相连法(neighbor-joining,NJ)构建[25],bootstrap 设为1000。Accession number 分别为 : XP_002263965.2、 XP_003541057.1、 XP_003624793.1、XP_003624793.1、BAA92894.1、XP_021319871.1、XP_004248085.1、XP_021968776.1、AT1G01280,XP_015648492.1、NP_001149251.1。
1.4 SiCYP703A2的克隆和载体构建
以芝麻95ms-5B 四分体时期的花药cDNA 为模板,依据Sinbase 数据库(http://www.sesame-bioinfo.org/Sinbase2.0/index. html)[25]中SiCYP703A2基 因 的编码序列(CDS),通过Primer 5 软件设计CDS 全长引物SiCYP703A2-forward: 5’-ATGCTCCCCCCAGGCCCTCC-3’和SiCYP703A2-reverse: 5’- CTAATTATACAAATGGGCTGCTAGTCTG -3’,使用I-5TM 2x High-Fidelity Master Mix 高保真酶进行扩增。PCR 程序如下:98℃3 min;98℃10 s,55℃10 s,72℃1 min 35 个循环;72℃2 min。PCR 产物经电泳检测后使用普通琼脂糖凝胶DNA 试剂盒(天根生化)进行回收纯化,然后连接到TSV-007 载体(擎科生物)上,并转化大肠杆菌感受态Top10,通过测序获得无突变的阳性克隆T-SiCYP703A2。以获得的无突变的阳性克隆T-SiCYP703A2 为模板进行扩增,获得带有酶切位点的目标片段,并通过TA 克隆连接到TSV-007 载体(擎科生物)上,测序获得无突变的阳性克隆T-ovSiCYP703A2。使用限制性内切酶NdeⅠ和SmaⅠ分别对表达载体PRI101-AN(TaKaRa,日本)和T-ovSiCYP703A2 进行双酶切,并将PCR 产物回收纯化,后使用T4 连接酶(TaKaRa,日本)进行连接,最终获得过表达载体(35S::SiCYP703A2)。
1.5 转基因阳性植株的获得和鉴定
通过液氮冻融法将重组表达载体转化农杆菌感受态(GV3101),挑选单克隆进行PCR 检测,后将阳性克隆接种于含有利福平(Rif)和卡纳(Kan)抗生素的液体LB 培养基中,200 r/min、28℃振荡培养至OD600为0.8~1.0。收集菌体并制备蔗糖悬浮液,通过花序浸染法[26]将过表达载体导入野生型拟南芥(Col-0)中,一周后进行第2次浸染。
将T0种子于具有卡纳抗性的1/2 MS 固体培养基上筛选阳性转化子,待阳性转化子具有2 片真叶时,移栽到营养土中生长,待种子成熟,收获T1代。将单株收获的种子于具有卡纳抗性的1/2 MS 固体培养基上筛选,通过统计绿苗和黄苗的分离比(符合3:1)来筛选单拷贝转基因家系,并将单拷贝超表达家系移栽到营养土中,待成熟后收获各单株种子,各株系通过卡纳抗性筛选,抗性不分离的株系即为纯合子,用于后续的功能验证。
1.6 基因表达分析
所有植物材料的RNA 提取使用Trizol 试剂(Invitrogen,美国),cDNA 合成利用HiScipt II One Step RT-PCR Kit 反转录试剂盒(诺唯赞生物)。半定量PCR(RT-PCR)试验使用Taq Master Mix(诺唯赞生物)。RT-PCR 程序如下:95℃3 min;95℃10 s,55℃10 s,72℃1 min 30个循环;72℃5 min。实时荧光定量PCR(qRT-PCR)体系配制使用ChamQ Universal SYBR qPCR Master Mix 试剂盒(诺唯赞生物),在Roche Light Cycler 480 系统进行qRT-PCR 分析。qRT-PCR 程序如下:95℃30 s; 95℃10 s, 60℃30 s,72℃1 min 40 个循环。以芝麻Si-Actin(SIN_1009011)的表达作为内参,检测SiCYP703A2在芝麻组织和花药中的相对表达水平。以拟南芥At-Actin(AT3G18780)的表达作为内参,检测拟南芥花粉壁发育相关基因的相对表达水平。qRT-PCR 试验设置3 个生物学重复和3 个技术重复,使用2-△Ct方法计算相对表达水平[27]。数据处理和统计分析使用Excel 2010和SPSS11.5软件。本研究中使用的引物列于表1。
1.7 花粉育性观察
在植株生长第8 周时,拍摄植株整体长势图。取当天早上正在开花的野生型和转基因拟南芥植株的花蕾,利用0.5%醋酸洋红染色,每朵花观察3个视野,饱满、暗红色的计为正常花粉,瘦小、染色浅或未染上色的计为不育花粉,在荧光倒置显微镜(IX71,Olympus)下观察并拍照。
取当天正在开花的野生型和转基因拟南芥植株的花蕾,加入4℃预冷的2.5%~3%戊二醛试剂,在4℃条件下进行固定(>4 h),之后使用PBS 缓冲液(0.1 mol/L、不含NaCl)浸洗3~5次。用系列梯度乙醇(30%、50%、70%、80%、90%、95%、100%)进行脱水,之后使用醋酸异戊酯浸洗两次。脱水后的花蕾采用临界干燥法进行干燥,用真空喷镀法对样品进行包金,最后使用扫描电镜(VEGA3,Tescan)观察并拍照。
2 结果与分析
2.1 芝麻细胞色素P450超家族成员的鉴定
将模式植物拟南芥的P450 蛋白序列与芝麻基因组进行比对和预测,获得了322条候选基因,通过Pfma数据库分析候选基因的结构域,剔除不含P450保守结构域的基因,最终在芝麻基因组中鉴定到了303个芝麻P450基因。氨基酸序列的全局比对分析结果显示,这些P450 基因之间的相似度范围在0.6%~99.8%之间,基因之间的相似度差异较大(附表1,首页OSID 二维码)。将273 条拟南芥P450 基因与预测的芝麻P450基因进行氨基酸序列的比对,并构建系统进化树,结果如图1 所示。芝麻303 个P450基因在双子叶植物特有的9个家族簇中均有分布,且主要分布在71 Clan、72 Clan、85 Clan 和86 Clan 这4 个 家 族 簇 中,71 Clan 包 含192 个P450 基因,占63.0%,其次是72 Clan,占12%,而拟南芥中除71 Clan外,在86 Clan中基因分布最多。

表1 本研究中所用的引物序列Table 1 Primers used in this study
芝麻P450 基因在其它5 个家族簇中分布较少,这与拟南芥P450基因的分布情况一致(图1)。依据P450 基因国际命名标准[10],氨基酸序列一致性在45%~55%之间为同一家族,以及基因的进化分析结果,对预测的芝麻P450 基因逐条进行一致性鉴定,并进行家族划分,家族分布情况如表2,此外,预测的芝麻P450 基因分布在9 个家族簇的43 个P450基因家族中,且主要分布于家族CYP71、CYP76、CYP72、CYP706和CYP81中(表2)。
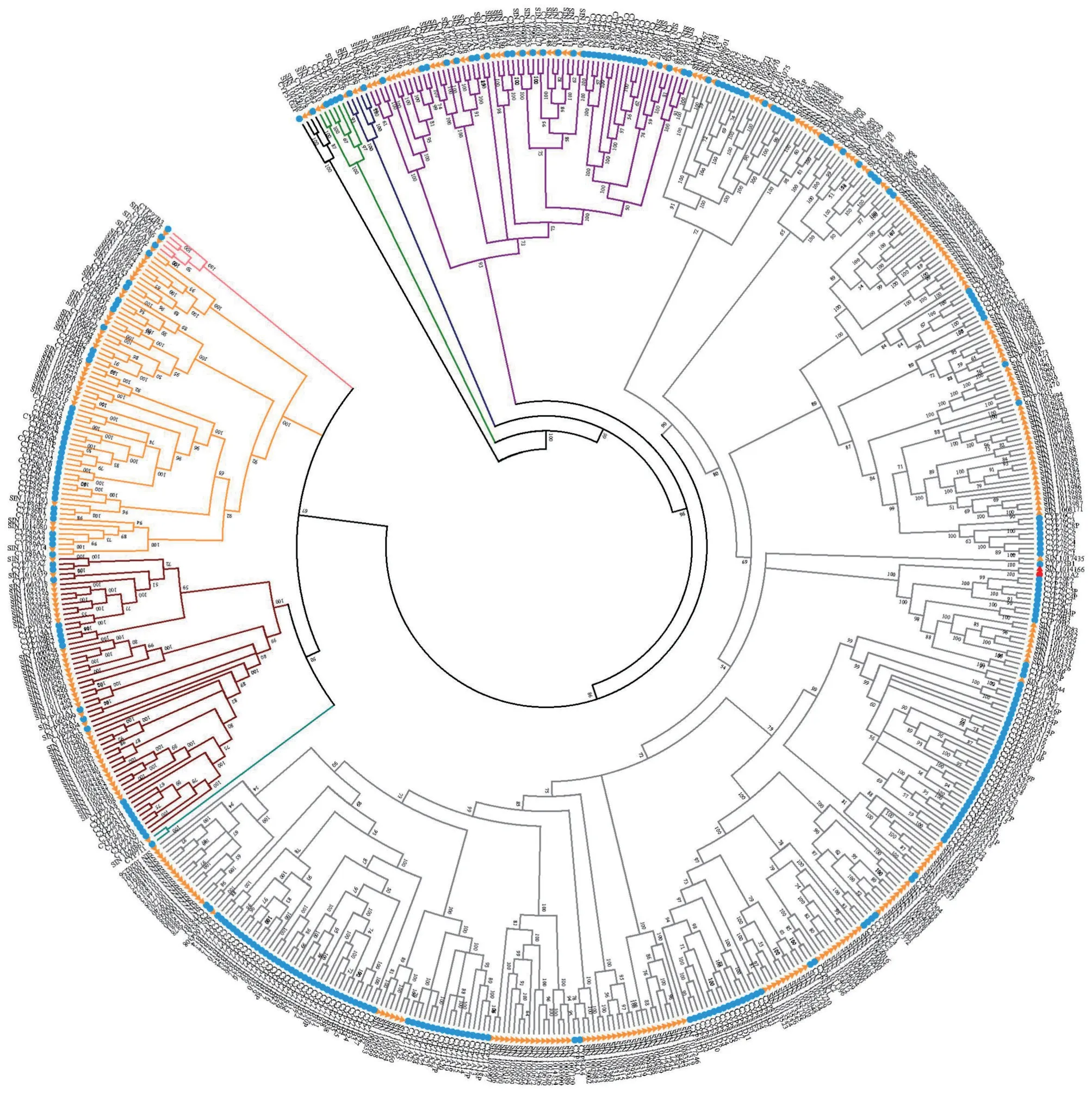
图1 芝麻CYP450基因与拟南芥CYP450基因的系统进化树Fig.1 Phylogenetic tree of CYP450 genes in Sesame and Arabidopsis
2.2 芝麻CYP450基因的染色体定位
本研究所鉴定到的303 个芝麻P450 基因全部是非重复基因,依据这些基因在基因组数据库中的位点信息,将这些基因的物理位置信息可视化。如图2 所示,277 个P450 基因在芝麻染色体16 个连锁群(LGs)上都有分布,并且呈现随机不均匀分布的特点,有26 个P450 基因被定位到了未被锚定的片段上,未在图中显示。这些基因在LG1、LG2、LG3和LG6上的分布较为密集,分别包括36、31、34和36个基因,而在LG13 和LG16 上均只有3 个基因分布。对应表2 的基因分布情况,可知71 Clan 的基因在每个连锁群上都有分布,且数目均匀;72 Clan 的基因在LG6 上分布较多,在其它连锁群上零星分布;86 Clan和85 Clan的基因在多个连锁群上无分布。
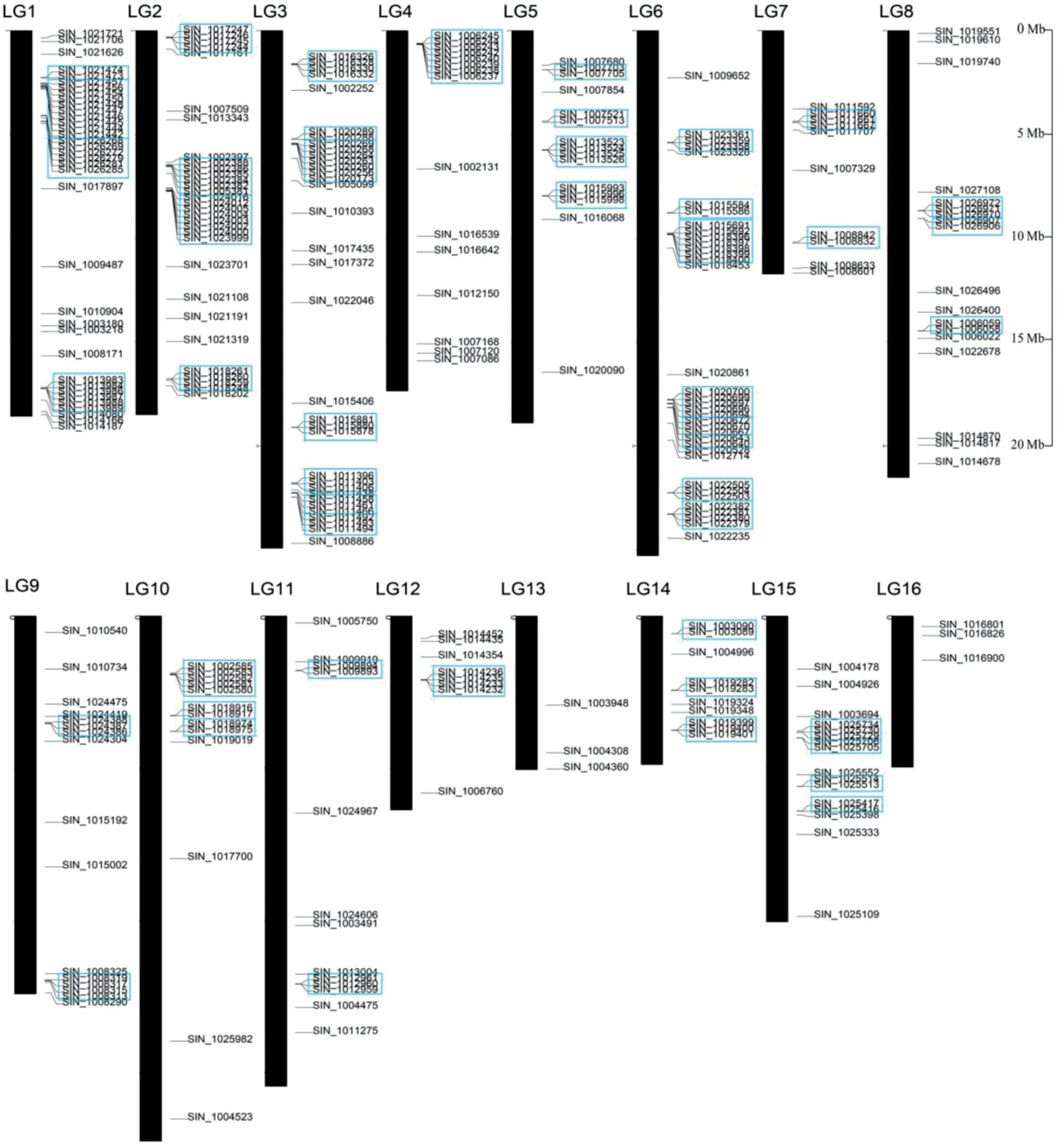
图2 芝麻P450基因在芝麻基因组中的分布特征Fig.2 Distribution characteristics of CYP450 genes in sesame genome
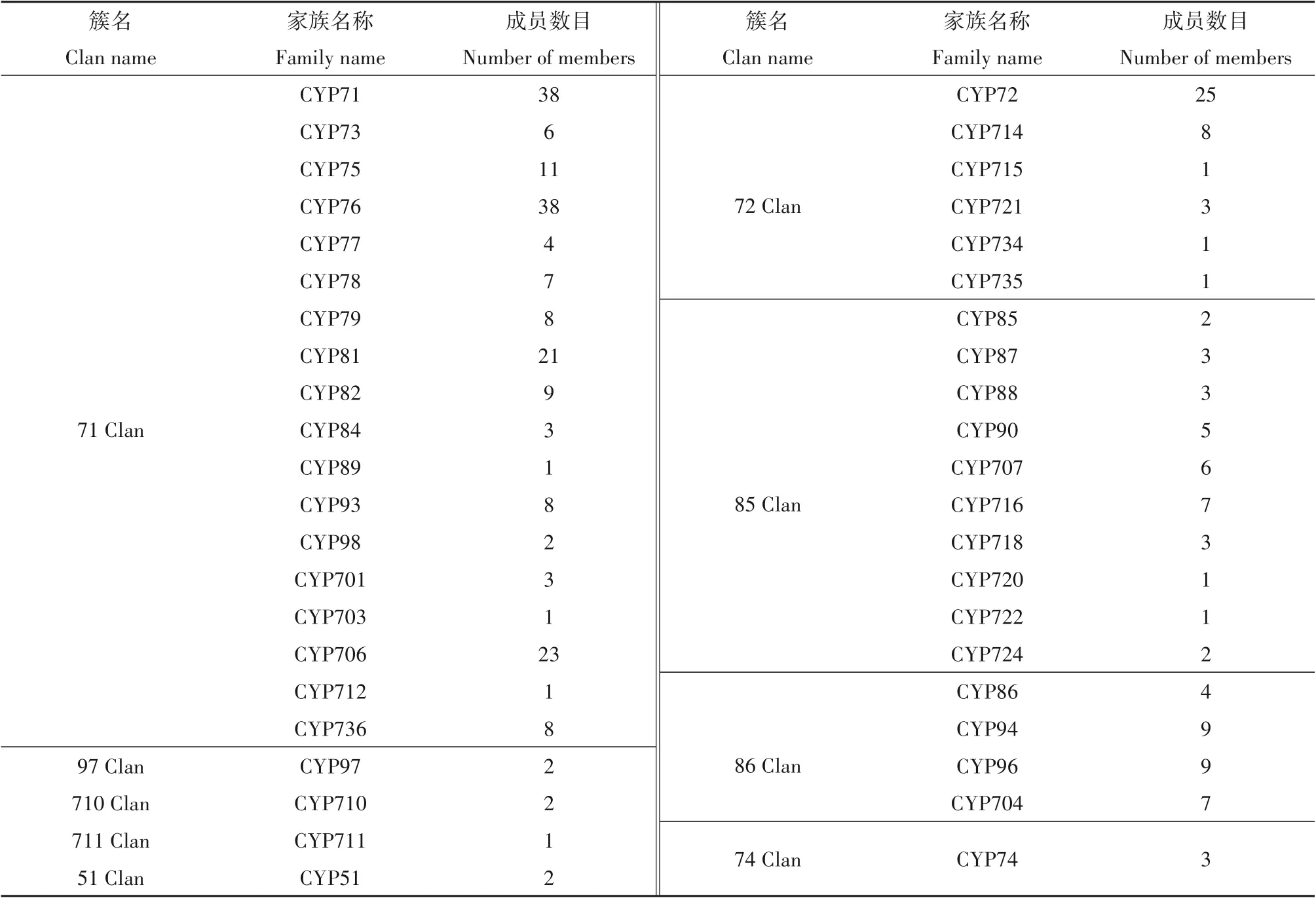
表2 预测的303个芝麻细胞色素P450基因的分布Table 2 Distribution of 303 predicted CYP450 genes in Sesamum indicum
芝麻P450 基因在连锁群上呈明显排列成簇的现象。依据基因簇的鉴定标准,被定位的287 个P450 基因中,共有169 个基因以基因簇的形式分布在连锁群上,占P450基因总数的58%。基因簇总个数为46个,主要出现在除LG13和LG16外的14个连锁群上,LG6上存在最多的基因簇。
2.3 SiCYP703A2的克隆与序列分析
进化分析结果显示芝麻SIN_1014166 基因与拟南芥CYP703A2基因聚为一类(图1),比对结果表明两个基因的氨基酸序列一致性为77%,符合同一亚家族的划分标准,因此,将芝麻SIN_1014166基因命名为SiCYP703A2。通过对芝麻全基因组序列比对,SIN_1014166与芝麻P450基因之间的氨基酸序列一致性最高为37%(附表1,首页OSID 二维码),低于同一家族的划分标准,同时,芝麻P450 基因的聚类分析显示(附图1,首页OSID 二维码),SIN_1014166为单独的分枝,表明SIN_1014166 基因是芝麻CYP703 家族的唯一成员,这与报道的植物CYP703家族通常只包含一个成员一致。
研究表明在拟南芥和水稻中CYP703 家族的基因都特异参与了花药发育[20,22]。为探究SiCYP703A2在芝麻中的功能,我们比较了SiCYP703A2在芝麻雄性不育两用系95ms-5AB 之间的序列差异,结果表明,SiCYP703A2在可育和不育株植株中的DNA 序列不存在差异。该基因CDS序列长度为1449 bp,编码482 个氨基酸。基因组结构分析表明,SiC⁃YP703A2含有两个外显子和一个内含子(图3A)。将SiCYP703A2的氨基酸序列提交至NCBI 的CDD程序进行结构域的分析,结果如图3B 所示,在氨基酸位点2-482 的位置存在一个P450 蛋白结构域,域名为PLN03112。预测的分子量为54.94kDa,理论等电点为6.24。氨基酸序列多重比较分析发现,SiCYP703A2与拟南芥、紫花苜蓿和水稻中的CYP703A 蛋白相似,在其C 末端都含有一个高度保守的血红素结合域(Heme binding domain)“FSAGKRKCPG”,此外还包含C 螺旋结构(C-Helix)“W×R×R”、PERF 结构域“P×R×”和K 螺旋结构(K-Helix)“E××R”(图3C)。SiCYP703A2与其它作物中已知的CYP703家族基因的系统进化树如图4所示,依据单子叶植物和双子叶植物的进化关系而分为两类,SiCYP703A2与茄科植物的番茄、牵牛花和美花烟草的亲缘关系较近。
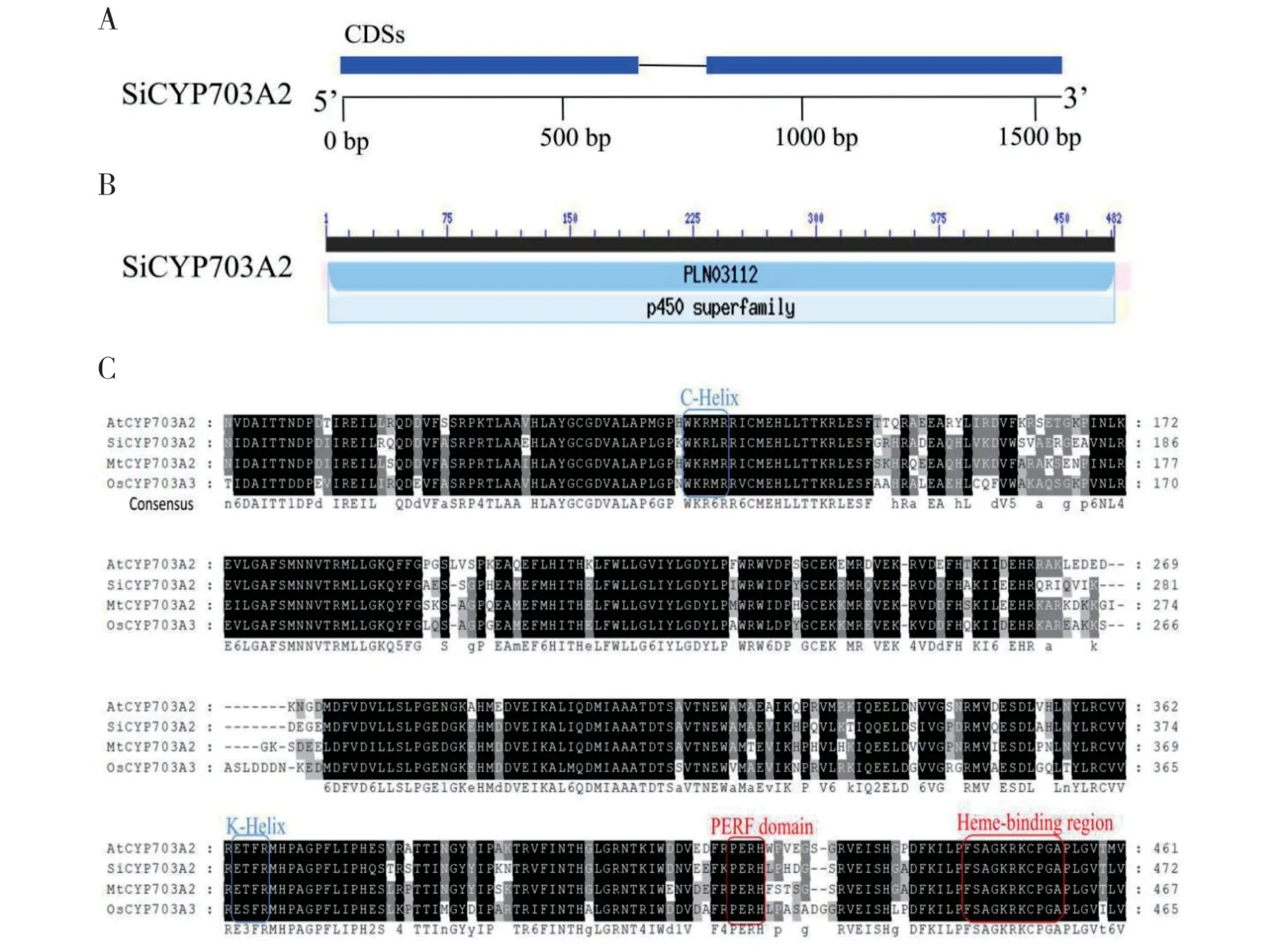
图3 SiCYP703A2的序列结构和多序列比对分析Fig.3 Gene structure and multiple sequence alignment analysis of SiCYP703A2

图4 SiCYP703A2基因的系统进化分析Fig.4 Phylogenic analysis of SiCYP703A2
2.4 SiCYP703A2的组织表达模式
为了解SiCYP703A2的表达模式,设计了SiC⁃YP703A2的特异引物,即将引物序列提交到NCBI的Primer-BLAST 程序,选择芝麻的基因组DNA 为模板,通过比对结果显示该引物的唯一产物为基因SiCYP703A2,表明该引物特异检测SiCYP703A2的表达。首先通过RT-PCR 和qRT-PCR 检测了SiC⁃YP703A2在芝麻不同组织中的表达水平,结果如图5A 和5B 所示,SiCYP703A2在花蕾组织中表达量最高,在根、茎、叶和果荚中几乎不表达,表明SiC⁃YP703A2可能参与生殖发育。进一步比较了SiC⁃YP703A2在芝麻雄性不育两用系95ms-5AB 的不同发育时期的花药中的表达模式。结果表明,SiC⁃YP703A2在花药发育四分体时期的表达量最高,在小孢子发育和花粉成熟期表达量较低。并且在不育植株四分体时期的表达水平显著高于可育植株,在小孢子时期和成熟花粉时期,二者之间没有差异(图5C 和5D),表明SiCYP703A2可能与芝麻雄性生殖器官的发育有关。
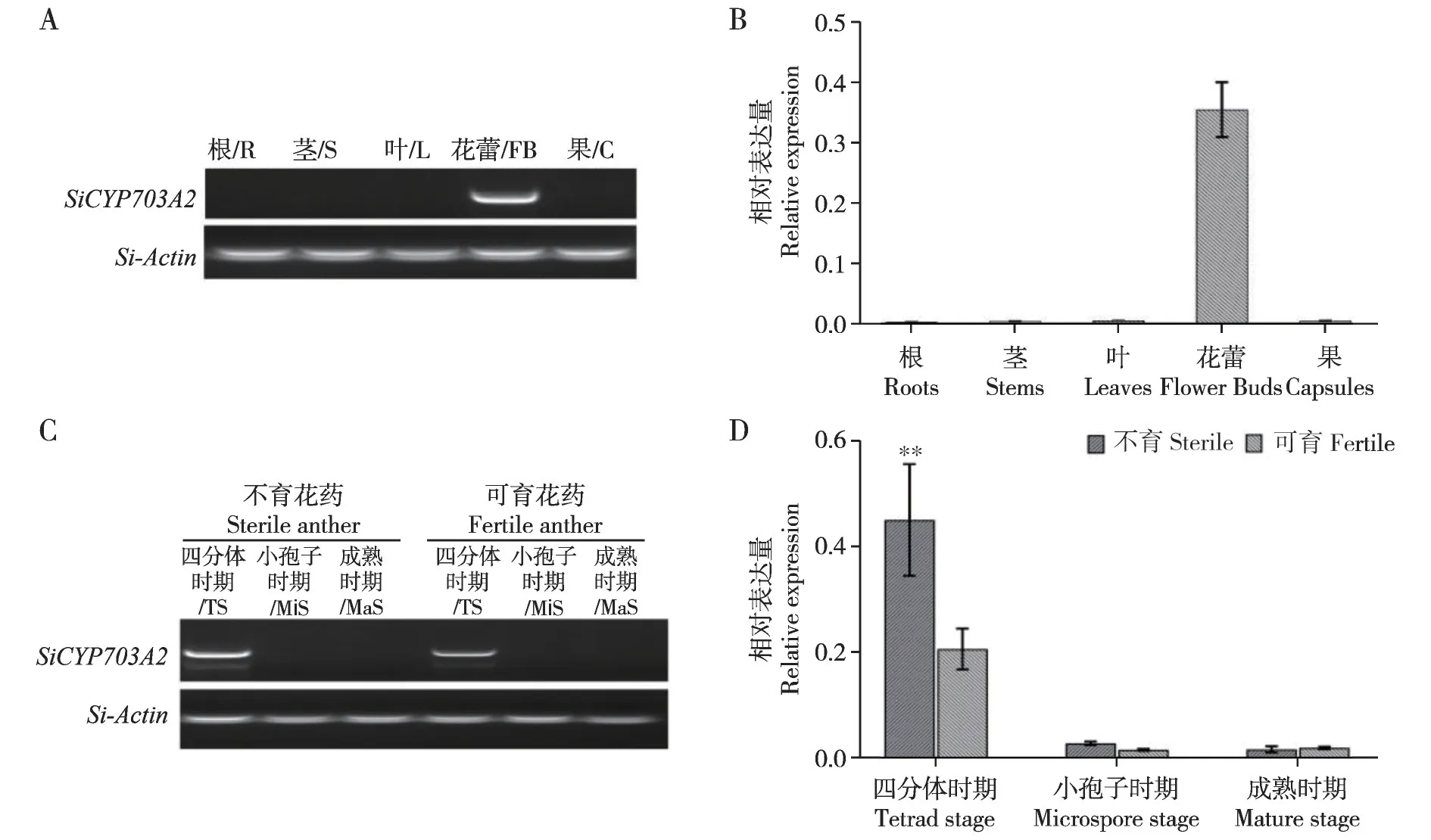
图5 SiCYP703A2的表达模式分析Fig.5 Expression patterns of SiCYP703A2
2.5 SiCYP703A2 超量表达拟南芥植株的育性鉴定
为进一步验证SiCYP703A2在花药发育中的功能,构建SiCYP703A2超表达载体,转化拟南芥,共获得19 个独立的转化子,并筛选到6 个单拷贝(绿苗和黄苗的分离比为3:1)的超表达家系并进一步获得其纯合株系(图6A)。选取表达量相对较高并且为纯系的的超表达家系OE-4 用于后续功能分析。通过观察OE-4 过表达家系与野生型对照的生长发现,二者在营养生长上无明显差异,但与野生型植株角果(图6C)相比,SiCYP703A2超表达植株的角果变短(图6D)。进一步通过醋酸洋红染色观察花粉活力,发现野生型(WT)的花粉粒圆润饱满,并且被染成红色,表明野生型花粉活力正常(图6E),而SiC⁃YP703A2超表达植株的花粉粒未被染上色(图6F),表明SiCYP703A2转基因植株的花粉活力降低。扫描电镜观察进一步表明,野生型的花粉粒饱满且具有规则的纹饰结构(图6G),而SiCYP703A2超表达植株的花粉粒表面布满不规则的凸起物,呈现异常的花粉壁结构(图6H),这些结果表明SiCYP703A2在转基因拟南芥花粉发育过程中发挥作用,并且可能影响正常花粉壁的形成。

图6 SiCYP703A2超表达植株鉴定和表型分析Fig.6 Identification and phenotypic analysis of SiCYP703A2-overexpressing plants
2.6 qRT-PCR检测花药发育相关基因的表达水平
为进一步了解SiCYP703A2在花粉发育中的作用,检测了一些花粉壁发育相关基因的表达水平,包括AtFLP1(AT5G57800)、AtCYP704B1(AT1G69500)、AtPKSA(AT1G02050)、AtPKSB(AT4G34850)、AtTK⁃PR1(AT4G35420)和AtTKPR2(AT1G68540)[28~31]。如图7 所示,与野生型相比,在SiCYP703A2超表达植株中,AtFLP1、AtTKPR1、AtTKPR2和AtPKSA的表达水平在花粉发育的7~14时期都出现明显下调,而AtCYP704B1和AtPKSB的表达水平在花粉发育前期即7、8 时期出现上调,在发育成熟时期即13、14 时期又出现下调。这些结果说明SiCYP703A2的过表达扰乱了花粉壁发育相关基因的正常表达,SiC⁃YP703A2可能参与了花粉壁发育。
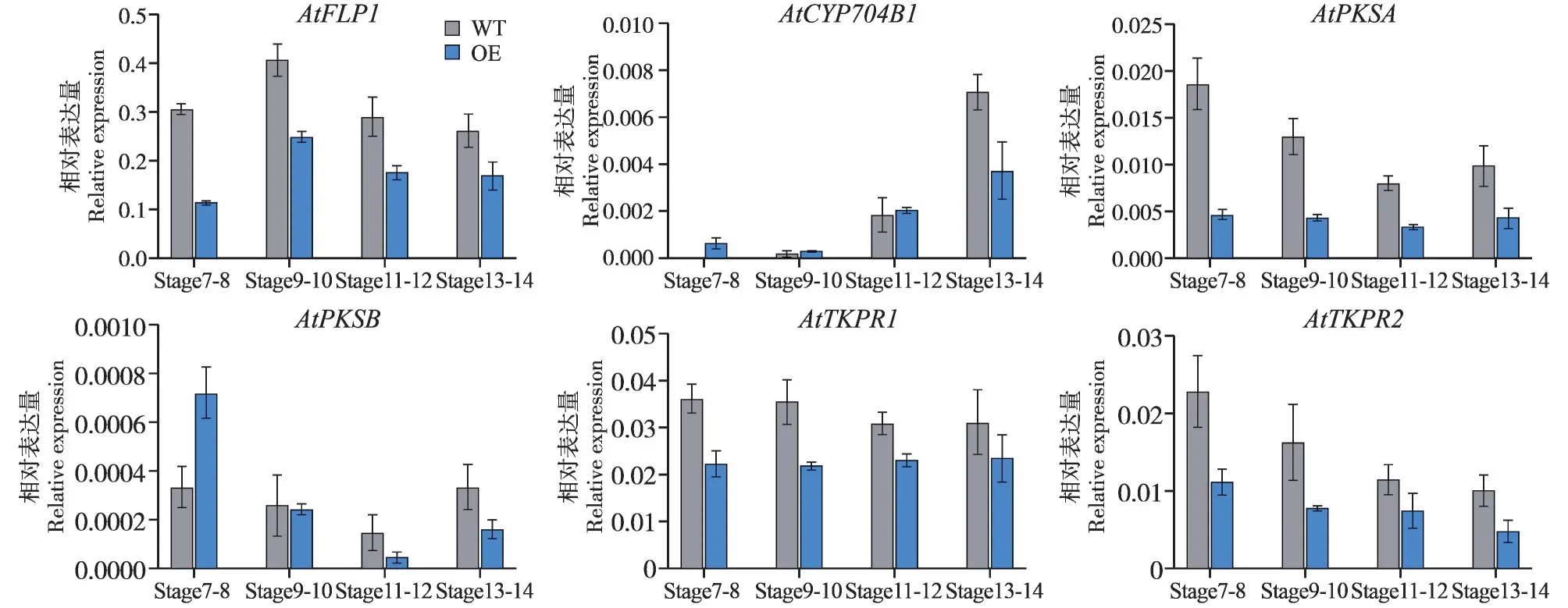
图7 花药发育相关基因的表达水平Fig.7 Relative expression levels of genes related to anther development
3 讨论与结论
近年来,研究人员不断从植物中分离出新的CYP450 家族和基因,Nelson 等人[32]构建的细胞色素P450 数据库涵盖了动物、植物和真菌等生物,包括拟南芥、番茄、玉米和大麦等在内的66 种植物。拟南芥基因组中含有286 个P450 基因(包括未分类的基因),分为45 个家族和72 个亚家族,水稻和烟草中分别包含47 和44 个P450 家族[32~34]。并且在拟南芥基因组数据中已有245 个P450 基因被注释,烟草中有173个P450基因已经找到EST证据[34]。本研究中,我们在芝麻中鉴定到了303 个可能的P450 基因(图1),系统进化分析表明,芝麻的303 个可能的P450 基因归属于43 个基因家族(表2),这与一般双子叶植物中的P450 基因分布情况相似。芝麻中CYP51、CYP75、CYP85、CYP711、CYP734 等家族的成员个数与拟南芥相同,但芝麻中不含有在拟南芥中存在的CYP708 和CYP709 家族。而在芝麻基因组中鉴定到的CYP736 家族,在拟南芥和水稻中均不存在,而CYP736 家族在苜蓿中也被鉴定到[12,35]。此外,研究表明拟南芥和水稻中共有的P450基因家族只有2/3,这可能与P450 基因在物种间的差异性和进化程度有关。我们发现芝麻P450 基因在16 个连锁群上是随机分布的,71 Clan 作为最大的簇,其簇内基因在每一个连锁群上都有分布。同时,14 条连锁群上出现多个基因密切排列的基因簇,这些基因一般具有较高的相似度,多数属于同一个基因家族。基因簇的出现一般是由于基因复制产生的,芝麻P450 基因较多的聚集在LG1、LG2、LG3 和LG6 这四个连锁群上,因此,推测它们相应的染色体上可能发生过P450基因的大量复制。
细胞色素P450 是生物界广泛存在的超大蛋白家族,蛋白结构和功能多样。P450 基因一般具有500 个左右的氨基酸,分子量在46~60 kDa,不同家族的氨基酸序列存在差异,但三级结构一般都由N端的折叠结构和C 端的螺旋束构成[36,37]。同时功能结构域也具有很高的保守度,特别是起接受电子作用的血红色素结合位点(Heme binding domain),该位点含有一个绝对保守的半胱氨酸(Cys)残基,与铁元素形成硫醇盐离子键,以及与血红素形成电子拉链的螺旋C(helix-C)和起到稳定血红素核心结构作用 的 螺 旋K(helix-K)[38]。 本 研 究 克 隆 到SiC⁃YP703A2,其编码482 个氨基酸,含有一个保守的P450 家族结构域,属于细胞色素P450 家族的成员(图3B)。多重序列比对结果显示SiCYP703A2含有多个在P450 蛋白中保守的结构域和螺旋结构(图3C),这些结构与P450 基因特征结构一致。SiC⁃YP703A2的系统进化结果表明SiCYP703A2与茄科植物的亲缘关系较近(图4),并且牵牛花CYP703A1基因已被揭示在花药发育阶段表达,推测SiC⁃YP703A2可能具有相似的表达模式。
芯片检测数据分析显示许多拟南芥P450 基因的表达具有组织特异性,如CYP88A3和CYP706A2在叶片组织中的表达水平较高,CYP77A6仅在花中特异表达,CYP708A2仅在根中特异表达,CYP81F4和CYP83B1在营养组织中特异表达[39]。烟草中,部分P450基因在整个生长过程中的表达水平都较高,如CYP73A1、CYP51G1、CYP82E2、CYP90A2和CYP97B1等,推测这些基因可能参与内源生理代谢途径[40]。本研究对芝麻SiCYP703A2进行组织表达分析,发现SiCYP703A2在花蕾组织中的表达量远高于其它组织,进一步检测SiCYP703A2在整个花药发育过程中的表达情况,发现其在芝麻花药发育的四分体时期表达量最高(图5)。这与拟南芥、水稻和牵牛花中CYP703A基因的表达模式相符[14~17],因此,推测SiCYP703A2可能发挥与这些基因相似的功能,参与芝麻雄性生殖器官的发育。
敲除CYP703A2基因的拟南芥植株呈现部分雄性不育表型,超微结构观察显示敲除系植株的花粉壁发育严重受损,外壁缺失[15]。水稻cyp703a3-2突变体的花药较小并且不产生花粉粒,电镜观察显示花药的表皮层出现缺陷[17]。我们在拟南芥中超量表达SiCYP703A2,发现转基因拟南芥植株的荚果长度与野生型相比明显缩短(图6C、D),并且花粉活力显著降低(图6E 和6F)。进一步通过扫描电镜观察花粉粒结构,发现过表达植株花粉粒表面出现异常突起物,花粉壁结构存在严重缺陷(图6G 和6H)。这可能是由于转基因植株花药中SiCYP703A2持续表达导致AtCYP703A2的精确表达被破坏。有研究也表明,ABORTED MICROSPORES(AMS)作为拟南芥绒毡层特定的拟南芥转录因子,其正确的表达时机对于绒毡层和花粉发育至关重要[41]。花粉外壁是花粉最外层的结构,其主要组成成分为孢粉素。目前在拟南芥、水稻、烟草等作物中存在一条保守的孢粉 素 生 物 合 成 路 径,由FLP1、ACOS5、PKSA/B、CYP703A、CYP704B、TKPR1/2等基因顺序作用,最终合成孢粉素的前体物质[42,43]。检测这些基因在SiCYP703A2过表达拟南芥中的表达水平,结果显示(图7)这些基因的表达水平都被扰乱,并且多个基因的表达水平出现明显下调,可能由于SiCYP703A2的过表达干扰了这些基因的正常表达,扰乱了正常的孢粉素代谢途径,最终导致花粉外壁异常。
综上所述,本研究鉴定了303 个芝麻细胞色素P450 基因,分布在43 个基因家族中。SiCYP703A2具有保守的P450 结构域和螺旋结构,可能是芝麻CYP703 家族的唯一成员。组织表达模式显示该基因在芝麻花蕾组织中高量表达。在拟南芥中超量表达SiCYP703A2导致转基因拟南芥花粉壁发育异常,植株育性降低,花粉结构异常,并且多个与花粉壁发育相关基因的表达水平发生变化。这些结果表明,在转基因拟南芥中SiCYP703A2可能通过影响花粉壁发育进而影响花粉育性。本研究对芝麻中P450 超家族进行了鉴定,并对其家族分类和进化进行分析,为芝麻P450基因的功能研究奠定了良好基础。同时揭示了芝麻SiCYP703A2基因在花粉发育中的功能,为芝麻杂种优势的利用提供基因资源和新思路。
