茎鞘非结构性碳水化合物积累运转与稻米品质对播期和行距配置的响应
2022-05-13谢裕林于雅洁董明辉张文地江贻
谢裕林 于雅洁 董明辉 张文地 江贻


摘要:为明确不同类型粳稻非结构性碳水化合物(NSC)积累运转特性及其与稻米品质形成的关系,以当地主栽的不同穗型和生育类型品种(系)为材料,进行4个播期(B1、B2、B3、B4)和2种行距配置处理,以期寻求优质食味品质形成的栽培调控技术途径。结果表明,大穗型水稻茎鞘NSC积累、运转和穗质量显著高于常规穗粒兼顾性品种;播期对茎鞘NSC积累运转影响较大,除杂交晚粳甬优1526成熟期含量无显著差异外,抽穗、成熟期NSC含量及转运量、穗干质量表现为B2>B1>B3>B4,B2处理均显著或极显著高于其他播期;B2播期处理NSC转运率显著高于其他播期,其他播期品种类型间存在差异;随播期延迟,碾米品质和外观品质变差,直链淀粉含量及蛋白质含量增加,胶稠度含量降低,崩解值、糊化温度以B2播期最高,B4播期最低,消减值趋势相反。与常规行距比较,宽窄行显著增加茎鞘NSC含量、转运量和穗干质量,晚熟品种与中熟品种类型间对转运率和籽粒贡献率影响有差异;直链淀粉含量及蛋白质含量增加,崩解值显著增加,胶稠度、消减值和糊化温度显著降低,同时碾米品质和外观品质得到改善。NSC转运量与多个米质性状指标呈显著或极显著相关,NSC转运率与垩白度呈显著负相关。品种、行距配置与播期之间互作显著影响NSC转运和稻米品质形成。选用适宜的品种、播期和行距配置能较好地协调NSC的积累运转,促进籽粒结实与米质改善。本研究为合理安排太湖地区水稻播种期及优化稻米品质提供了理论基础。
关键词:粳稻品种;播期;栽插方式;NSC积累与运转;稻米品质
中图分类号:S511.01 文献标志码: A
文章编号:1002-1302(2022)08-0093-08
籽粒结实和稻米品质的优劣与水稻籽粒灌浆有非常密切的关系,籽粒灌浆充实的同化物一部分来源于抽穗后稻株自身的光合产物,一部分来源于花前贮藏在茎鞘中的非结构性碳水化合物(non-structural carbohydrate,简称NSC),后者的贡献率可达30%[1]。籽粒开始灌浆后,叶片和茎鞘中贮存的NSC以蔗糖的形式经过一系列装卸载过程运输至籽粒,在多个淀粉酶的协同作用下合成淀粉,促进籽粒灌浆[2-3],因此促进花前茎鞘积累物质的转运启动和转运量对籽粒初期的生长和籽粒结实有重要作用[4-5]。增加抽穗前茎鞘NSC的积累,同时提高NSC向穗部转运的效率是促进籽粒灌浆、形成高产和优质的重要途径。促进花前茎鞘物质的积累和转运不仅与品种自身特性有关,也是一个复杂的生理生化过程。抽穗前,茎鞘NSC积累主要受叶片中蔗糖磷酸合成酶(SPS)含量的影响[6],适度水分胁迫和减少氮素供应等措施可以提升叶片蔗糖磷酸合成酶活性,进而提高叶片蔗糖输出能力,同时促进花前水稻茎鞘NSC积累[7-9]。籽粒灌浆结实期淀粉水解并转化为蔗糖,随后茎鞘NSC被重新利用[10]。抽穗后茎鞘中蔗糖磷酸合成酶的活性迅速上升,茎鞘中淀粉含量则迅速下降[9],籽粒淀粉形成期库的活性增加可促进茎鞘NSC向籽粒运转[4]。氮肥供应量的改变可以调控水稻茎鞘NSC 的积累和运转,具体表现为低氮处理条件下,水稻茎鞘内参与淀粉合成与水解相关酶的活性提高,促使茎鞘花前NSC积累,并且花后茎鞘NSC转运能力得到提升[11];适量氮肥处理会使花前茎鞘NSC积累增加,花后茎鞘蔗糖磷酸合成酶活性提高,同时籽粒蔗糖合成酶活性也会相应提高,进而促进茎鞘NSC向籽粒转运[12-13] 。除氮肥外,水分同样参与到水稻茎鞘NSC积累与转运的调控过程中,抽穗前浅干湿交替灌溉可促进花前茎鞘NSC积累[14],灌浆结实期适度水分胁迫可提高相关酶活性,籽粒库活性增加,促进茎鞘NSC活化并向籽粒运输[15]。氮肥处理及水分等处理通过调控水稻叶片蔗糖的合成与运输调控水稻茎鞘中NSC的积累及转运,但现阶段茎鞘NSC积累与转运的内在生理调控机制尚不清晰,生态气候条件与栽培因子间的互作对茎鞘NSC的积累与转运的综合效应研究还不多,NSC的积累转运对稻米品质形成与调控的关系尚不清楚。
因此,本研究选择不同生育类型和庫容量的粳稻品种为材料,通过设置不同播期和宽窄行机插方式,研究茎鞘NSC积累和运转特性及其与籽粒产量和品质的相关关系,以期进一步探讨NSC积累转运的内在生理机制,同时为不同类型品种的丰产优质协同提升提供科学的理论依据和实践指导。
1 材料与方法
1.1 试验时间与地点
2020 年试验田块位于江苏太湖地区农业科学研究所望亭基地(31°27′10″N、120°25′12″E)。试验田前茬为小麦,试验土壤质地为黏土,有机质含量为23.8 g/kg、速效氮含量为161.4 mg/kg、速效磷含量为8.7 mg/kg、速效钾含量为123.9 mg/kg。
1.2 供试品种
以当地主栽的3种不同类型粳稻品种为材料,分别为迟熟中粳稻新品系苏1785 (V1,全生育期 145 d)、中熟晚粳稻品种苏香粳 100 (V2,全生育期 168 d)、大穗型杂交晚粳稻品种甬优1526(V3,全生育期168 d),常规稻移栽苗数为2苗,杂交稻移栽苗数为1苗。
1.3 试验设计
试验设置播期(B)、品种(V)、行距配置(T)3种不同处理,采用裂区设计,其中播期为主区,4个播期分别为05-13(B1)、05-23(B2)、06-02(B3)、06-12(B4),不同播期播种量相同,且3个品种同期栽插,25 d秧龄移栽;品种为主裂区,3个品种分别标注为V1、V2、V3;行距配置为副裂区,分为等行距30 cm(T1)、宽窄行(T2,宽行与窄行间隔栽插,宽行为40 cm,窄行为20 cm),不同行距配置密度一致。
小区面积为15 m2(6 m×2.5 m),品种小区采取随机排列的方式,2次重复。主区间做埂隔离,埂体采用塑料薄膜进行覆盖,以确保各品种小区可以进行单独排灌。
各试验小区总施氮量为270 kg/hm2,其中氮磷钾配比为N ∶P2O5 ∶K2O=2 ∶0.5 ∶1,氮肥施用比例为基肥 ∶分蘖肥 ∶穗肥=3 ∶4 ∶3,分蘖肥和穗肥分2次等量施用;磷肥全作基施;钾肥分基肥、促花肥2次等量施用。
水分管理及病虫草害防治等其他相关栽培措施均按照高产栽培要求实施。
1.4 测定项目与方法
1.4.1 非结构性碳水化合物样品取样与测定
抽穗期选取试验小区内生长一致的单茎挂牌若干,并于抽穗期和成熟期分别取样10个单茎,按照茎鞘、叶片和穗分开烘干,待测NSC含量。
将所取样品(抽穗期和成熟期茎鞘烘干样品)粉碎后过0.15 mm筛,使用热乙醇法提取可溶性总糖和蔗糖,蒽酮比色法和间苯二酚法分别测定其含量。用高氯酸提取淀粉,蒽酮比色法测定其含量。
NSC转运量=抽穗期NSC质量-成熟期NSC干质量;
NSC转运率=NSC转运量/抽穗期NSC质量×100%;
NSC对籽粒贡献率=NSC转运量/穗干质量×100% 。
1.4.2 稻米品质样品取样与测定
每处理成熟期取平均茎蘖数的5穴稻株,脱粒后的稻谷置于尼龙丝沙袋内自然条件风干2~3个月后,按中华人民共和国行业标准《米质测定方法》(NY/T 83—2017)测定糙米率、精米率、整精米率、垩白粒率、垩白度、蛋白质含量、直链淀粉含量、胶稠度等。
1.5 數据分析
使用Microsoft Excel 2010将数据录入,随后采用DPS 7.05对所获得的数据进行分析。
2 结果与分析
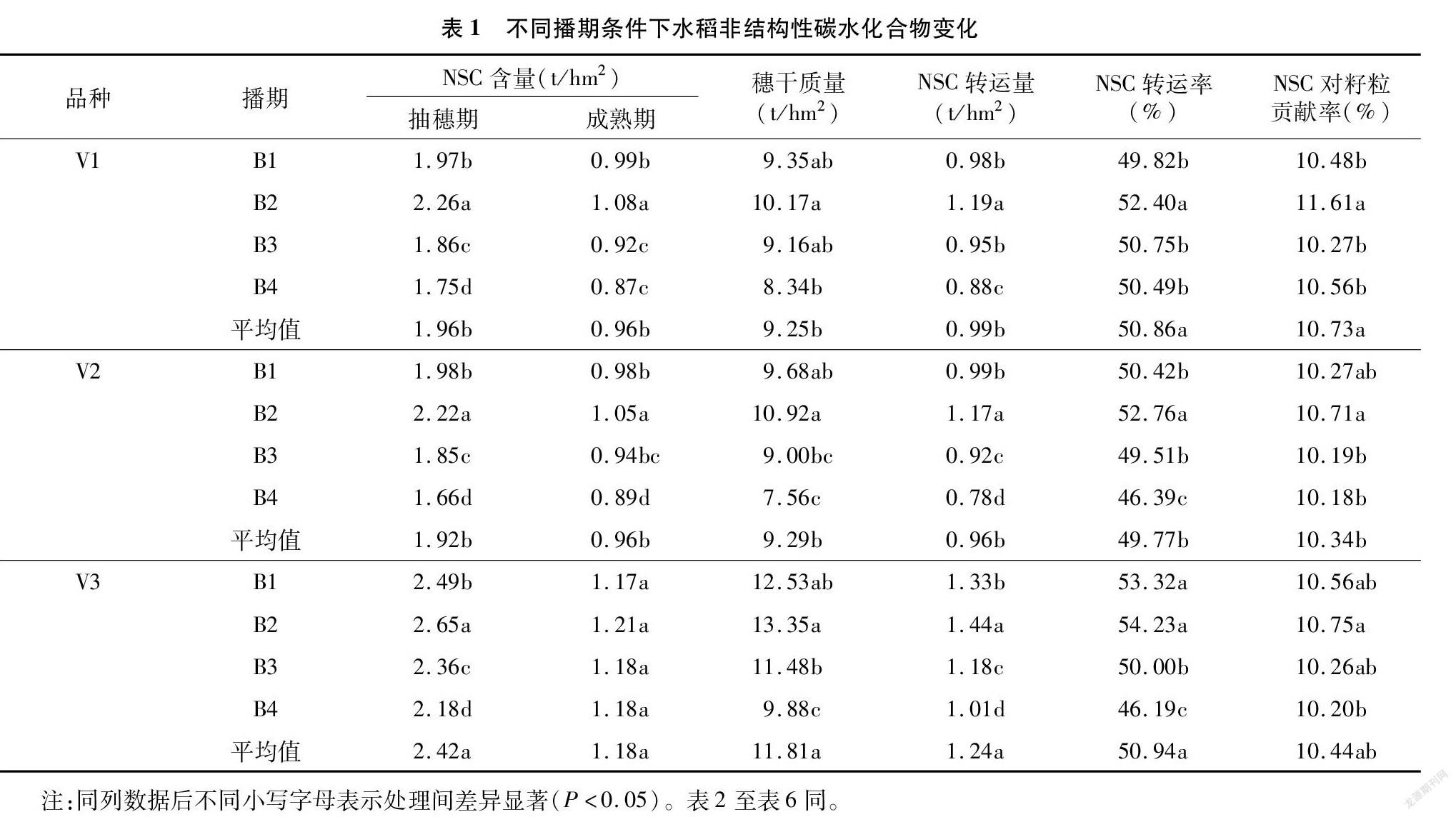
2.1 不同播期条件下茎鞘NSC积累和转运
从表1可以看出,播期对茎鞘中NSC含量有较大的影响,在本试验播期条件下,抽穗期、成熟期2个时期茎鞘NSC含量表现为随播期延迟呈先上升后下降的趋势,以B2处理的茎鞘NSC含量最高,各播期间的大小排序为B2>B1>B3>B4,3个品种趋势一致;其中,抽穗期NSC含量,B2处理显著高于其他处理,成熟期的NSC含量除杂交晚粳甬优1526(V3)播期间无显著差异外,其他2个品种(V1、V2)播期间差异均达显著水平。穗干质量各播期表现为B2>B1>B3>B4,其中B2与B4间差异显著。不同播期下NSC转运量与转运率趋势较为一致,除品种苏1785(V1)转运率呈现B2>B3、B4>B1播期外,其他2个品种(V2、V3)的NSC转运率与3个品种 NSC转运量均表现为B2>B1>B3>B4播期,且差异显著。3个品种茎鞘NSC对籽粒的贡献率,品种间略有差异,但3个品种适期播期B2处理显著高于最晚播期B4处理。
2.2 不同行距配置条件下NSC累积和转运
不同行距配置条件下NSC累积转运情况见表2,行距配置对3种不同类型水稻品种的NSC累积和转运均有显著影响。在宽窄行栽插方式下,抽穗期及成熟期水稻茎鞘NSC含量均显著高于常规行距栽培,3个品种前者分别比后者高出8.51%、14.44%、12.5%和9.78%、10.87%、9.73%;穗干质量也均表现为宽窄行方式下显著高于常规栽插方式,3个品种前者比后者分别高出10.05%、17.33%和9.18%。
不同行距配置对NSC转运的影响同样存在显著差异。宽窄行栽培条件下3个水稻品种NSC转运量均显著高于常规栽培方式,前者比后者分别高出6.19%、19.32%和13.79%;而对NSC转运率及NSC对籽粒贡献率的影响不同品种间存在差异,结果显示,中熟晚粳苏香粳100(V2)及籼粳杂交稻甬优1526(V3)在宽窄行栽培条件下的NSC转运率及NSC对籽粒贡献率高于常规行距栽培,而迟熟中粳苏1785(V1)则呈相反的趋势。
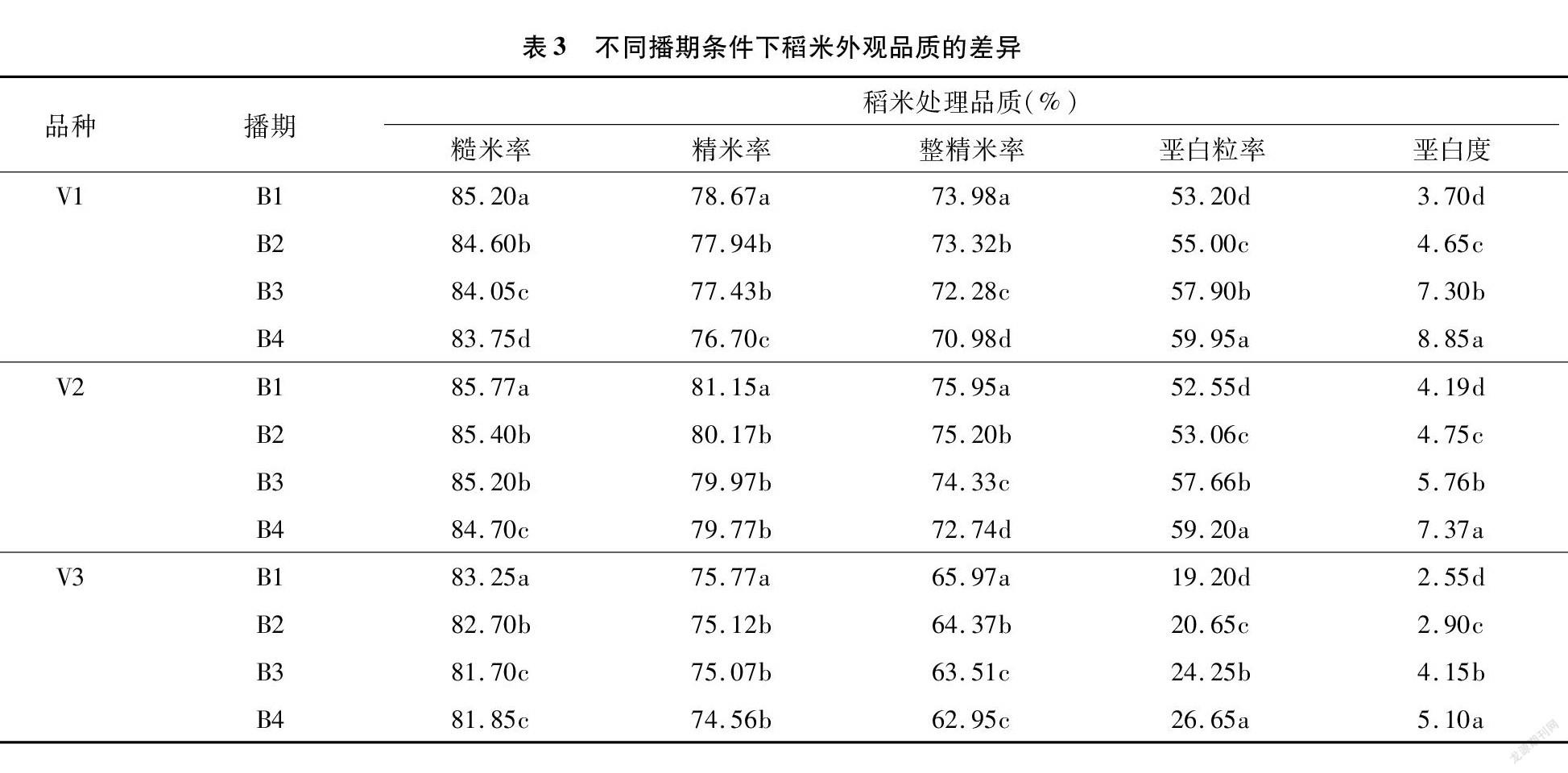
2.3 不同播期对稻米品质的影响
从表3可以看出,不同播期会对稻米品质产生影响,随着播期的延迟,3个不同类型品种稻米碾米品质指标值均呈下降趋势,播期越晚碾米品质越差。播期对外观品质有显著影响,随播期延迟垩白度与垩白粒率均显著增加,且不同播期之前存在显著差异,3个品种的趋势一致。
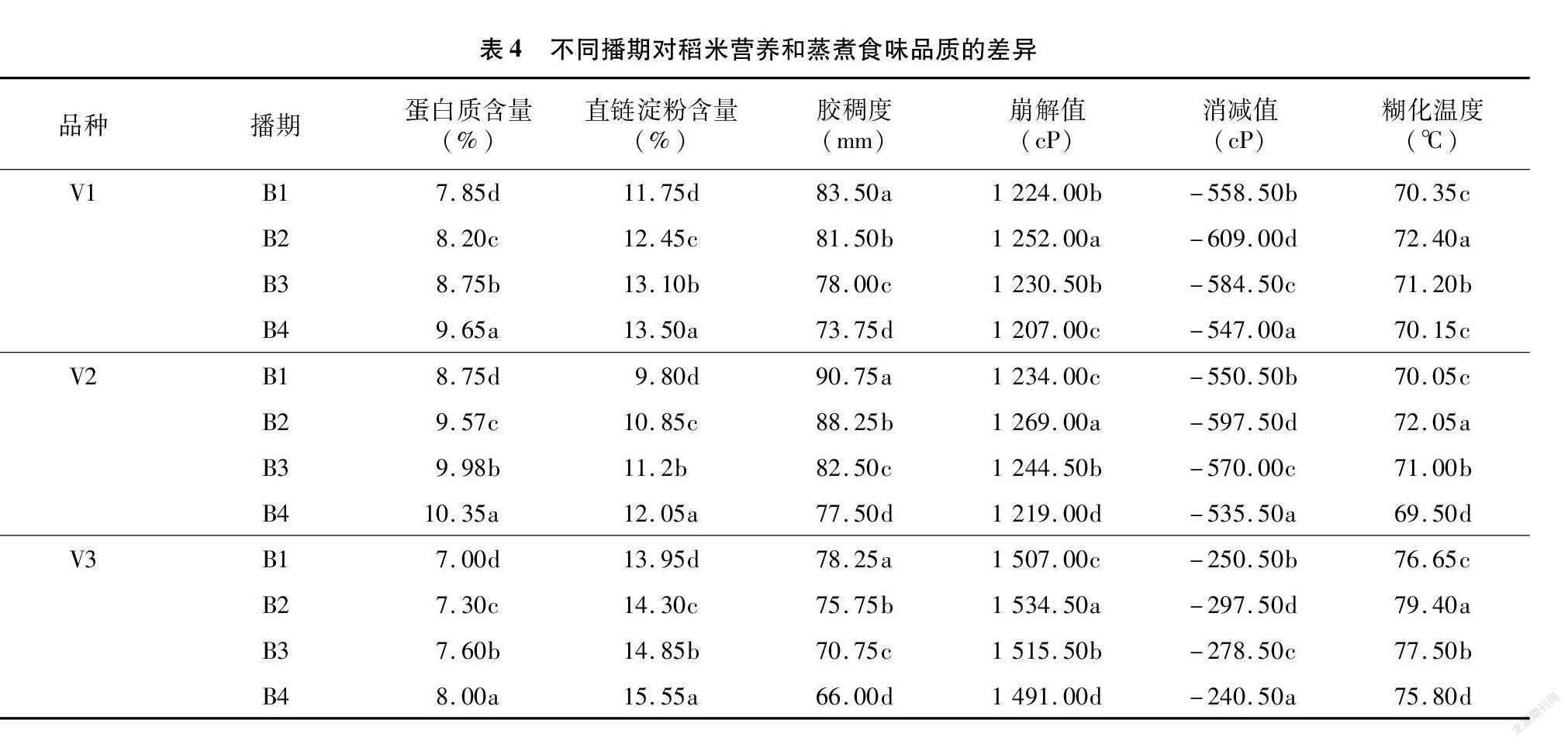
蛋白质含量在不同播期间也存在显著差异,随播期延迟,蛋白质含量逐渐增加。在蒸煮食味品质上,随着播期的推迟,3个不同类型品种直链淀粉含量呈增加趋势,而胶稠度则呈相反的趋势,随播期的推迟,胶稠度数值逐渐下降。播期对淀粉粘滞特性亦产生较大影响,崩解值、糊化温度均表现为在B2播期条件下最高,B4播期条件下最低,总体表现为B2>B3、B1>B4,消减值则表现与崩解值相反的趋势(表4)。
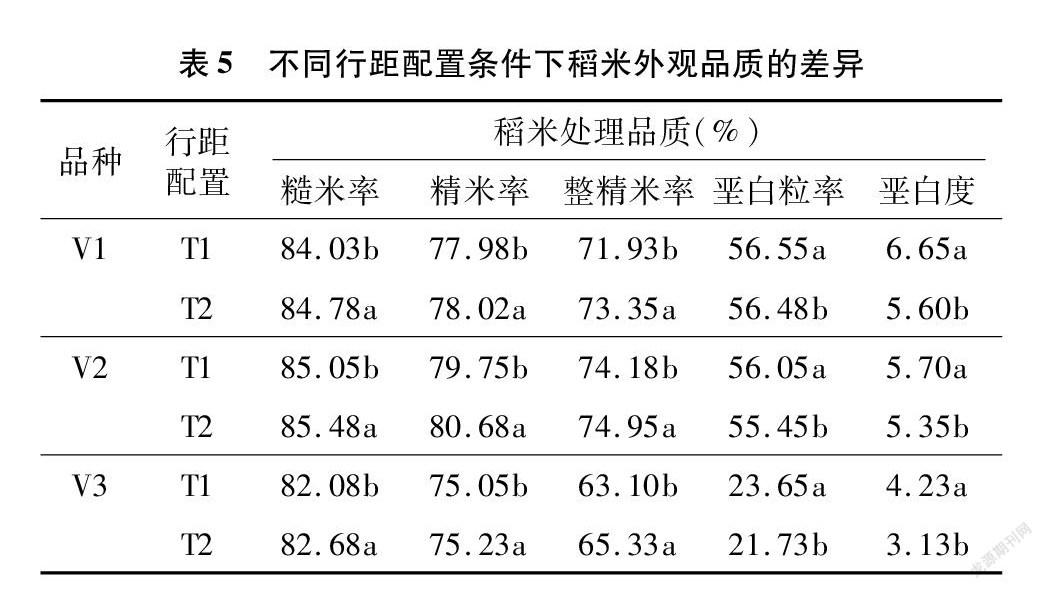
2.4 不同行距配置对稻米品质的影响
从表5可以看出,在宽窄行栽培条件下,稻米的碾米品质指标均显著高于常规栽培方式,3个品种的整精米率则分别高出1.97%、1.04%和3.53%;而外观性状明显改善,垩白粒率显著低于常规行距栽培,垩白度呈相同趋势,3个品种的垩白粒率和垩白度分别下降了0.12%、1.07%、8.12%和15.79%、6.14%、26.00%。
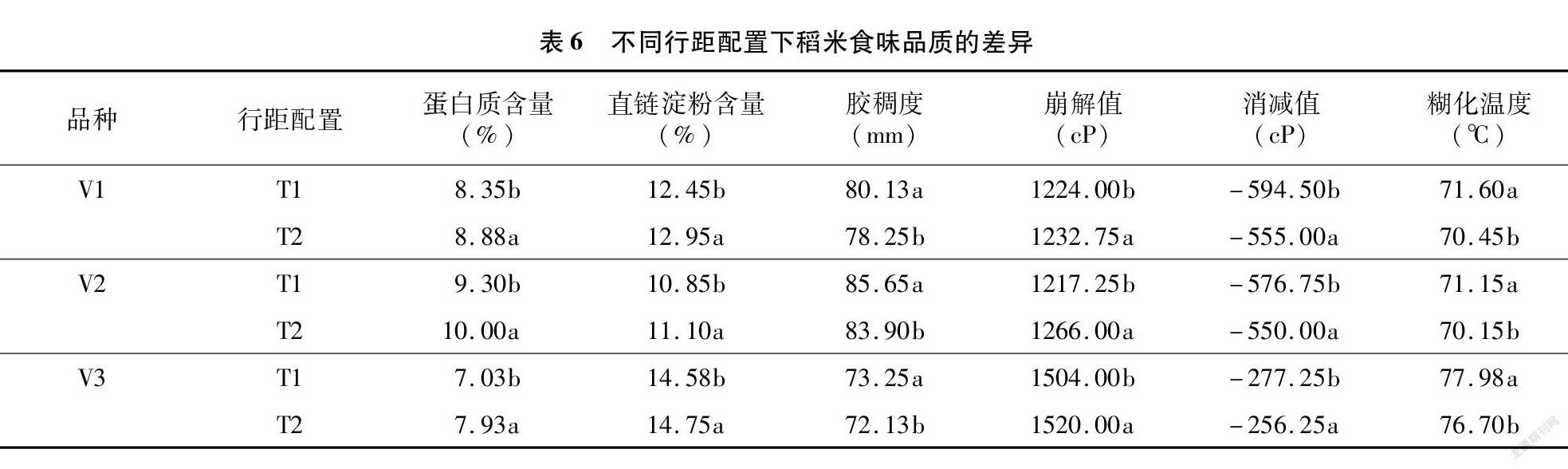
从表6可以看出,不同行距配置对蛋白质和直链淀粉含量均有较大影响,均表现为宽窄行栽插显著高于常规行距栽插,3个品种蛋白质和直链淀粉前者分别比后者增加6.35%、7.53%、12.80%和4.02%、2.30%、1.17%;胶稠度则表现相反的结果。崩解值亦表现为宽窄行栽插显著高于常规栽插,消减值、糊化温度结果则与之相反。
2.5 非结构性碳水化合物运转与品质性状的相关分析
相关性分析(表7)表明,茎鞘中NSC含量及转运与稻米品质形成关系密切。抽穗期及成熟期NSC含量与整精米率、垩白粒率、垩白度及蛋白质含量呈极显著负相关,与糊化温度、崩解值、消减值呈极显著正相关,抽穗期NSC含量与直链淀粉含量呈显著正相关,成熟期NSC含量与直链淀粉含量呈极显著正相关。NSC转运量与整精米率呈显著负相关,与垩白粒率、垩白度及蛋白质含量呈极显著负相关,与消减值呈显著正相关,与崩解值及糊化温度呈极显著正相关。NSC转运率除与垩白度呈显著负相关之外,与其他稻米品质相关性状不大。NSC对籽粒贡献率与稻米品质性状的相关性也不显著。
2.6 不同处理及互作对茎鞘NSC转运及稻米品质性状的影响
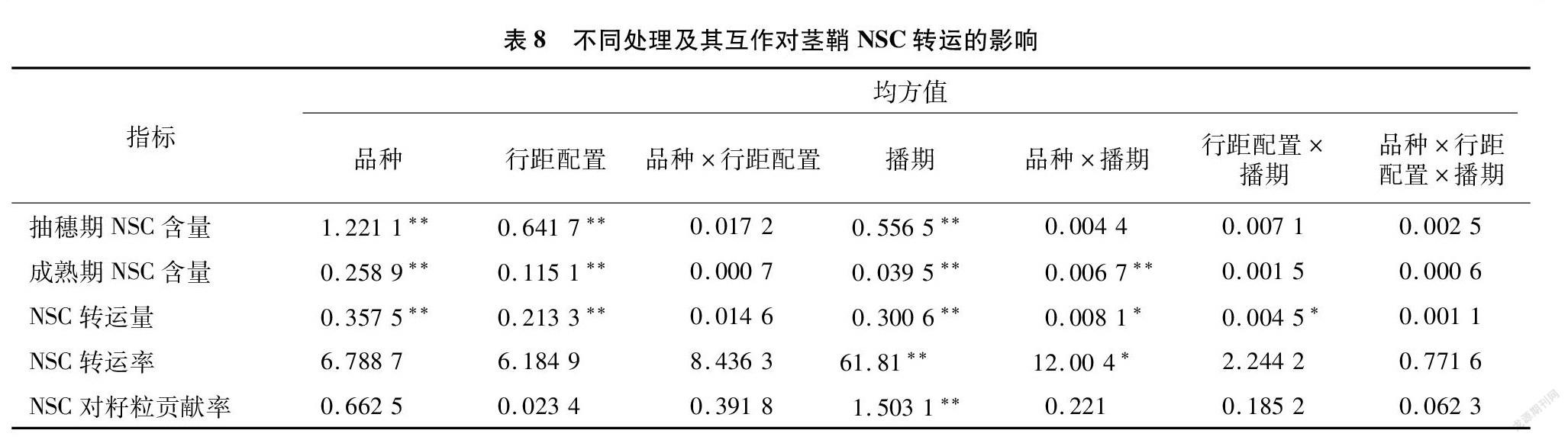
将3个品种、行距配置及播期处理与茎鞘NSC转运量、转运率及对籽粒的贡献率进行方差分析(表8),结果显示,品种、行距配置、播期较大地影响了抽穗期与成熟期NSC含量及NSC转运量,差异达极显著水平;NSC转运率及NSC对籽粒贡献率在不同播期处理下的差异亦达到极显著差异。品种×播期互作极显著影响成熟期NSC含量,显著影响了NSC转运量及NSC转运率,行距配置×播期互作则显著影响NSC的转运量。
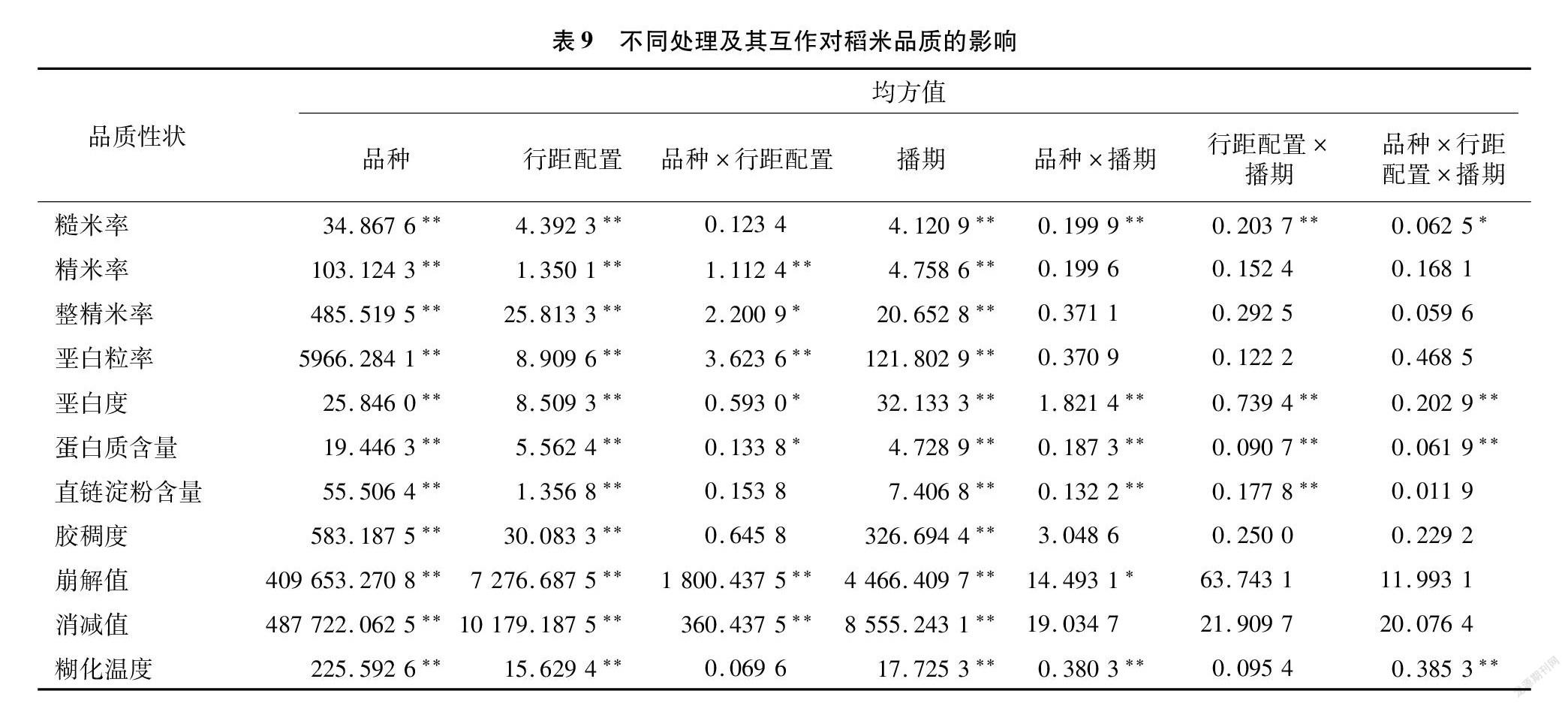
将不同处理与品质性状进行方差分析(表9),结果表明,11个主要性状在不同品种、行距配置及播期处理下的差异均达到极显著水平,表明稻米品质性状同时受品种、行距配置和播期的影响。分析发现,品种、行距配置、播期3因素之间对稻米品质形成均有互作效应,品种×行距配置互作显著或极显著影响了精米率、整精米率、垩白粒率、垩白度、蛋白质含量、崩解值和消减值;品种×播期互作显著或极显著影响了糙米率、垩白度、蛋白质含量、直链淀粉含量、崩解值和糊化温度;行距配置×播期互作极显著影响了糙米率、垩白度、蛋白质含量和直链淀粉含量;品种×行距配置×播期之间互作显著或极显著影响了糙米率、垩白度、蛋白质含量和糊化温度。进一步分析表明,垩白度、蛋白质含量2个品质性状受不同品种类型和不同处理及互作的影响最大,均达显著或极显著水平。
3 讨论与结论
3.1 不同类型水稻非结构性碳水化合物的积累与运转
作为籽粒灌浆的重要能量物质来源,水稻茎鞘中储藏的非结构性碳水化合物提供了籽粒灌浆所需的重要能量来源,其含量对籽粒产量和品质的形成均有重要调控作用[16]。尤其在水稻品种演化过程中,株型和穗型都发生了系统的变化,NSC的重新再分配决定了其对籽粒产量形成的重要性;同时,近年来由于气候变化(如全球气候变暖)等环境因素影响及外界非生物逆境等因素,导致籽粒结实和籽粒产量形成的稳定性被破坏[17]。本研究条件下,大穗型杂交晚粳品种甬优1526群体干物质生长量和颖花量最大,穗籽粒产量最高,其抽穗期和成熟期NCS积累量均明显高于2个中等穗型常规粳稻品种,NSC的转运量和转运率也较高;NSC的积累和运转与生育类型关系不明显。说明生物量大的品种前期积累的NSC量大,转运到籽粒中的NSC量也大。相关研究表明,与早期品种比较,现代品种茎鞘 NSC 由于生物量的增加而得到积累。叶片内水解蔗糖的酸性转化酶(INV)和蔗糖合成酶(分解方向)(SSc)活性低、合成蔗糖的蔗糖合成酶(合成方向)(SSs)和蔗糖磷酸合成酶(SPS)活性高会导致NSC积累量上升[18]。本试验材料均为现代品种,但大穗型超级稻品种甬优1526的NSC积累、转运优势更明显,这是由于作为大库容水稻,甬优1526在抽穗前茎鞘NSC积累量高,同时灌浆结实期NSC表观转运量大[1]。因大库容量类型水稻源库比率较小[19],在水稻生长过程中可以通过采取增源的栽培措施来获得高产。
3.2 播期和行距配置对非结构性碳水化合物的积累与运转
NSC的积累量多少与播期有非常密切的关系,太湖地区无论是不同生育类型品种还是不同穗型品种,无论是抽穗期和成熟期NSC含量、穗干质量,还是NSC的运转量、运转率和对籽粒的贡献率均为5月23日播期处理最大,说明播期对NSC积累运转的影响非常大。播期是一个综合性的因素,温度、光照、降水等都是对光合生产有重要影响的生态因子,但目前的调控机制还不清楚,相关报道还不多[20-21]。与等行距栽插相比,宽窄行栽插表现出增加稻田透光和通风、减少病虫害、提高水稻抗倒伏能力和水稻产量等优点[22-23]。本试验通过宽窄行栽插,有效地增加了3个供试品种抽穗期、成熟期NSC的积累量和转运量,穗干质量显著增加;但NSC的转运率以及对籽粒的贡献率在不同熟期類型品种间有差异,2个晚熟粳稻甬优1526和苏香粳100宽窄行的NSC转运率及对籽粒贡献率显著高于常规处理,迟熟中粳新品系苏1785结果则相反。宽窄行栽插将宽行和窄行的综合优势进行了充分的融合,宽行设置可以增强水稻通风透光能力,利于水稻生长,而窄行设置则可以充分利用阳光进行光合作用,有利于形成高光效水稻群体,通过宽窄行综合优势的利用,使水稻中光合产物的积累和运转量提高[24]。不同类型品种间NSC的转运率有差异,其机制不甚清晰,有可能与品种的生育特性和生态适应性有关,也有可能与品种分蘖特性和穗型有关,分蘖力强、穗型大的品种在宽行距下通风透光能力增强,改善冠层对光能的充分利用,可以适度协调个体和群体之间的矛盾,达到优化群体结构的作用,在该栽插方式下更能挖掘生长潜力[25]。
3.3 NSC积累运转与稻米品质形成的关系
前文所述花前储藏在茎鞘中的NSC灌浆期被活化后向籽粒运输,茎鞘中储藏的NSC有70%左右转运到籽粒中促进籽粒灌浆,贡献率甚至可达30%左右;灌浆初期,水稻茎鞘中NSC可提高水稻库活性,启动并促进灌浆,说明花前茎鞘储藏的NSC不仅是灌浆物质,也与代谢密切相关。在遭遇生物逆境和非生物环境影响后,光合能力减弱,在此条件下,水稻茎鞘中NSC的转运对于水稻产量形成及对抗逆境胁迫的作用更加明显[26],提高茎鞘NSC利用率是寻求水稻稳产的重要途径[27]。
籽粒灌浆充实好产量高,稻米品质的优劣同样决定于籽粒灌浆充实状况,淀粉作为稻米最主要的储藏物质,其合成量直接影响水稻的产量,淀粉颗粒的理化特性决定稻米的商品性、蒸煮与食用品质[28]。花前茎鞘NSC能否有效促进灌浆一方面取决于储藏的NSC量,即光合生产能力,另一方面取决于是否高效运转,即同化物向库器官的合理转运能力。从本试验结果分析,茎鞘NSC的转运量与多个稻米品质性状呈显著或极显著相关关系,说明NSC的积累和运转对稻米品质的形成有密切的联系,但机制尚并不清楚。同时,不同播期和栽插方式(宽窄行)对茎鞘NSC积累、运转和稻米品质均有显著的调节效应,适宜的播期和宽窄行栽插下,有利于光合同化物的生产,从而提高茎鞘NSC积累量;灌浆期由于NSC既作为灌浆的能量物质来源,也作为提高库活性的信号物质,适宜的温光条件有效促进茎鞘中NSC向籽粒的转运,同时在适宜的温光资源和群体结构下叶片光合功能期的延长,增加了籽粒形成所需的同化物,籽粒充实良好,产量和品质能有机协调[29]。通过育种或栽培手段协同提茎鞘NSC积累、转运和产量、米质协同是可行的。
本研究表明,與常规穗粒兼顾型粳稻品种比,大穗型杂交粳稻生物量大、颖花量多,茎鞘NSC积累、运转量和运转率高;适宜的播期和宽窄行栽插方式条件下,茎鞘NSC积累量和转运量大,籽粒产量高;播期和宽窄行栽插显著影响稻米品质的形成,茎鞘NSC含量和运转量与稻米品质密切相关。对NSC积累运转和品质形成调控中,品种、播期和行距配置之间存在互作效应。江苏太湖地区通过筛选适宜的品种、播期和宽窄行机插方式能较好地协调NSC的积累运转,促进籽粒结实与米质改善。
参考文献:
[1]潘俊峰,崔克辉,向 镜,等. 不同库容量类型基因型水稻茎鞘非结构性碳水化合物积累转运特征[J]. 华中农业大学学报,2015,34(1):9-15.
[2]王义杰,张绍杰,赖 艳,等. 水稻糖代谢相关酶和糖类转运蛋白编码基因的鉴定和表达分析[J]. 湖北农业科学,2019,58(22):185-193,197.
[3]李瑞清,谭瑗瑗,闫 影,等. 水稻胚乳淀粉合成及其育种应用[J]. 核农学报,2019,33(9):1742-1748.
[4]Yang J C,Zhang J H,Wang Z Q,et al. Activities of enzymes involved in sucrose-to-starch metabolism in rice grains subjected to water stress during filling[J]. Field Crops Research,2003,81(1):69-81.
[5]Wada H,Masumoto-Kubo C,Tsutsumi K,et al. Turgor-responsive starch phosphorylation in Oryza sativa stems:a primary event of starch degradation associated with grain-filling ability[J]. PLoS One,2017,12(7):e0181272.
[6]Ishimaru K,Hirotsu N,Madoka Y,et al. Quantitative trait loci for sucrose,starch,and hexose accumulation before heading in rice[J]. Plant Physiology and Biochemistry,2007,45(10/11):799-804.
[7]Yang J C,Zhang J H,Wang Z Q,et al. Activities of starch hydrolytic enzymes and sucrose-phosphate synthase in the stems of rice subjected to water stress during grain filling[J]. Journal of Experimental Botany,2001,52(364):2169-2179.
[8]李国辉,崔克辉. 氮对水稻叶蔗糖磷酸合成酶的影响及其与同化物积累和产量的关系[J]. 植物生理学报,2018,54(7):1195-1204.
[9]Li G H,Hu Q Q,Shi Y G,et al. Low nitrogen application enhances starch-metabolizing enzyme activity and improves accumulation and translocation of non-structural carbohydrates in rice stems[J]. Frontiers in Plant Science,2018,9:1128.
[10]Wang G Q,Hao S S,Gao B,et al. Regulation of gene expression in the remobilization of carbon reserves in rice stems during grain filling[J]. Plant and Cell Physiology,2017,58(8):1391-1404.
[11]潘俊峰,王 博,崔克辉,等. 氮肥对水稻节间和叶鞘非结构性碳水化合物积累转运特征的影响[J]. 中国水稻科学,2016,30(3):273-282.
[12]Zheng Y M,Ding Y F,Liu Z H,et al. Effects of panicle nitrogen fertilization on non-structural carbohydrate and grain filling in indica rice[J]. Agricultural Sciences in China,2010,9(11):1630-1640.
[13]Liang W X,Zhang Z C,Wen X X,et al. Effect of non-structural carbohydrate accumulation in the stem pre-anthesis on grain filling of wheat inferior grain[J]. Field Crops Research,2017,211:66-76.
[14]Zhang H,Yu C,Kong X S,et al. Progressive integrative crop managements increase grain yield,nitrogen use efficiency and irrigation water productivity in rice[J]. Field Crops Research,2018,215:1-11.
[15]Yang J C,Zhang J H.Crop management techniques to enhance harvest index in rice[J]. Journal of Experimental Botany,2010,61(12):3177-3189.
[16]Wang D,Wolfrum E J,Virk P,et al. Robust phenotyping strategies for evaluation of stem non-structural carbohydrates (NSC) in rice[J]. Journal of Experimental Botany,2016,67(21):6125-6138.
[17]Gomez-Zavaglia A,Mejuto J C,Simal-Gandara J.Mitigation of emerging implications of climate change on food production systems[J]. Food Research International,2020,134:109256.
[18]袁木松. 水稻品種演替过程中茎鞘非结构性碳水化合物积累与转运特征的变化及其机理研究[D]. 武汉:华中农业大学,2017.
[19]Wang X,Dai T B,Jiang D,et al. Yield-formation and source-sink characteristics of rice genotypes under two different eco-environments[J]. The Journal of Applied Ecology,2005,16(4):615-619.
[20]Cai C,Li G,Di L J,et al. The acclimation of leaf photosynthesis of wheat and rice to seasonal temperature changes in T-FACE environments[J]. Global Change Biology,2020,26(2):539-556.
[21]柳 军,王学林,张玉龙,等. 播期对江淮一季稻生长发育及产量形成的影响[J]. 江苏农业科学,2020,48(22):49-55.
[22]孙 良,刘 兵,陈 旋,等. 差速水稻钵苗Z字形宽窄行移栽机构设计[J]. 农业工程学报,2017,33(17):18-27.
[23]朱德泉,储婷婷,武立权,等. 宽窄行配置对机插中晚稻生长特性及产量的影响[J]. 农业工程学报,2018,34(18):102-112.
[24]Feng L Y,Raza M A,Chen Y K,et al. Narrow-wide row planting pattern improves the light environment and seed yields of intercrop species in relay intercropping system[J]. PLoS One,2019,14(2):e0212885.
[25]许俊伟,孟天瑶,荆培培,等. 机插密度对不同类型水稻抗倒伏能力及产量的影响[J]. 作物学报,2015,41(11):1767-1776.
[26]Bagherikia S,Pahlevani M,Yamchi A,et al. Transcript profiling of genes encoding fructan and sucrose metabolism in wheat under terminal drought stress[J]. Journal of Plant Growth Regulation,2019,38(1):148-163.
[27]Kumar U,Laza M R,Soulié J C,et al. Compensatory phenotypic plasticity in irrigated rice:sequential formation of yield components and simulation with SAMARA model[J]. Field Crops Research,2016,193:164-177.
[28]Zhang C Q,Zhou L H,Lu Y,et al. Changes in the physicochemical properties and starch structures of rice grains upon pre-harvest sprouting[J]. Carbohydrate Polymers,2020,234:115893.
[29]Du B B,Wang Q F,Sun G L,et al. Mapping dynamic QTL dissects the genetic architecture of grain size and grain filling rate at different grain-filling stages in barley[J]. Scientific Reports,2019,9:18823.
