温性草甸草原植物碳、氮、磷化学计量与贮量对放牧强度的响应
2022-05-10李瑞强辛晓平朱晓昱闫瑞瑞
王 淼,张 宇,李瑞强,曹 娟,辛晓平,朱晓昱,闫瑞瑞*
(1.中国农业科学院农业资源与农业区划研究所/呼伦贝尔草原生态系统国家野外科学观测研究站/国家土壤质量呼伦贝尔观测实验站,北京 100081;2.内蒙古自治区环境在线监控中心,内蒙古 呼和浩特 010055;3. 农业农村部环境保护科研监测所,天津 300191)
草地生态系统是我国陆地面积最大的生态系统和重要的自然资源,总面积近4×108hm2,占全国陆地面积的41%[1-2],具有多重的生态和生产功能。然而,近年来由于人类活动加剧和全球气候变化的共同作用,全球草地生态系统均在退化[3-4],我国90%以上的草地正在发生退化。放牧作为草地生态系统中最主要的利用和管理方式,通过采食和践踏改变植物群落物种组成、土壤理化性质等,从而直接或间接地影响土壤碳、氮、磷含量;植物因适口性的优劣及避牧性的强弱,受动物采食的程度不同,植物的补偿性生长直接影响元素在植物体内的循环及再分配。
碳、氮、磷是植物生长发育和物质循环过程中的关键元素,结构性物质碳影响生态系统的碳循环,氮、磷是植物生长的限制性养分[5]。同时,碳、氮、磷元素之间存在着强烈的耦合作用,在植物生长、生理机制调节、生态系统物质和能量循环等方面发挥着重要作用。Niu等[6]认为,放牧草地植物叶片磷含量显著高于围封草地,放牧草地植物叶片氮含量显著高于围封草地,放牧草地植物碳氮比显著低于围封草地[7-9]。放牧强度的增加对植物群落全磷有显著影响,放牧降低了植物的群落碳、磷含量。而李红琴等[10]的研究结果支持了中度放牧干扰理论,在中度放牧处理下植物群落碳含量最大,重度放牧显著提高叶片的氮含量,放牧对叶片磷含量变化无显著影响,不放牧有利于氮和磷在草地生态系统内的周转。Han等[11]提出植物体内氮、磷为协同关系,呈现显著正相关。生态化学计量学,是研究碳、氮、磷之间计量关系的一门学科,也是研究元素之间平衡关系的学科[12],植物体内的碳氮比和碳磷比能够反映出植物的养分利用效率,在植物体内,营养生长旺盛的植物为了合成蛋白质和氨基酸需要大量的氮元素,就会得到较低的碳氮比;在植株快速生长的过程中,需要大量蛋白质的供给,因此磷元素与植株的生长关系十分密切,生殖生长旺盛的植物拥有低的碳磷比[13]。植物碳氮比和碳磷比在不同放牧强度下均表现为与氮、磷的规律相反,植物磷、碳磷比与土壤磷、碳磷比呈显著正相关,而植物碳、氮、碳氮比和氮磷比与土壤碳、氮、碳氮比和氮磷比无显著相关性[14]。近年来,随着草畜系统优化的需要,进行草地放牧生态中各营养元素在各贮量内的动态特征研究十分必要,草地各组分碳、氮、磷贮量的季节动态模式与其对应生物量变化规律一致;碳、氮、磷贮量均与生物量呈极显著正相关[15-16],但不同放牧强度下碳、氮、磷贮量的变化规律一直没有定论。
内蒙古呼伦贝尔是我国温带草甸草原分布最集中、最具代表性的地区,是我国目前原生植被保存较完整、生物多样性最丰富的草地生态系统类型。但是,近年来受气候变化和过度放牧等的影响,草地退化,生态环境日益恶化。本研究依托呼伦贝尔草原生态系统国家重点野外实验站大型控制放牧实验平台,从生态化学计量学的角度出发,开展不同放牧强度下优势植物和退化指示植物碳、氮、磷含量、生态化学计量特征以及元素贮量的变化研究,探寻优势植物和退化指示植物化学计量特征及贮量的变化规律,揭示放牧对草原生态系统化学计量学及贮量影响的研究,对维持放牧草地化学计量学平衡与管理水平具有重要意义。
1 材料与方法
1.1 研究地概况
研究区位于大兴安岭西麓呼伦贝尔羊草草甸草原的核心带,试验基地依托呼伦贝尔草原生态系统国家重点野外实验站(49°32′~49°34′N,119°94′~119°96′E),海拔670 m左右,属温带半干旱大陆性气候,年平均降水量400 mm左右,年均气温-5~-2℃,≥10℃年积温1580~1800℃,无霜期110 d左右[17]。试验区土壤为黑钙土或栗钙土;植被类型为羊草+杂类草草甸草原,主要物种有羊草(Leymus chinensis)、贝加尔针茅(Stipa baicalensis)、羽 茅(Achnatherum sibiricum)、裂叶蒿(Artemisia tanacetifolia)等,伴生种有草木樨状黄芪(Astragalus melilotoides)、草地早熟禾(Poa pratensis)等[18],主要退化指示种包括冷蒿(Artemisia frigida)、星毛委陵菜(Potentilla acaulis)、二裂委陵菜(Potentilla bifurca)等。
1.2 研究方法
1.2.1 试验设计及样品采集
放牧试验设6个水平的放牧强度处理(即不放牧G0.00、较轻度放牧G0.23、轻度放牧G0.34、中度放牧G0.46、重度放牧G0.69、极重度放牧G0.92),载畜率分别为0.00、0.23、0.34、0.46、0.69和0.92 Au·hm-2,其中以500 kg肉牛为一个标准家畜肉牛单位,每个处理3次重复(图1),试验区设置为18个面积为5 hm2的试验小区,试验区总面积为90 hm2。在试验区面积一定、放牧时间相同的条件下,用250~300 kg的放牧肉牛头数来控制不同放牧强度的实施,6个放牧强度肉牛头数分别为0、2、3、4、6、8头,数量共计69头。试验于2009年开始,每年6月1日开始放牧,10月1日终止放牧,为期120 d。整个放牧期间牛全天在样地,不补充饲料,保证充足的饮水和盐分供给。

图1 不同放牧强度试验图
2019年8月(生长季),在不同的放牧处理选取5个1 m×1 m的样方开展取样,共计选用样方90个。本研究选取了6种植物羊草、贝加尔针茅、寸草苔,退化种冷蒿、星毛委陵菜、二裂委陵菜。在样方内采用齐地面剪割法,将6种植物叶片及茎以种为单位放在不同的纸袋中,在65℃烘箱中烘48 h后将其称重,植物样品用球磨仪充分磨碎后用于有机碳、全氮、全磷的测定。
1.2.2 数据测定
各元素的测定方法分别为:植物组织碳含量采用重铬酸钾氧化-外加热法测定;植物氮含量采用半微量凯氏定氮法测定,磷含量采用H2SO4-H2O2消煮,熔融,钼锑抗比色法测定。
植物元素贮量(g·m-2)=植物元素含量(g·kg-1)×植物生物量(g·m-2)/1000
1.3 数据处理
采用Excel 2017对数据进行整理,运用SPSS 21.0进行统计分析,采用one-way ANOVA对不同放牧强度下的植物碳、氮、磷含量及其化学计量特征进行方差分析。显著性区间定义为95%水平,并用LDS和Duncan法进行平均值之间的多重比较,显著性水平设为P<0.05,极显著性水平设为P<0.001,利用Origin 2017进行绘图。
2 结果与分析
2.1 不同放牧强度植物碳、氮、磷养分含量变化
6种植物的碳含量随着放牧强度的增加呈现不同的变化趋势(图2)。优势物种羊草的碳含量随着放牧强度的增加而显著减小(P<0.05),与不放牧相比,G0.69和G0.92的碳含量分别减小5.62%和6.85%;退化指示种冷蒿的碳含量随着放牧强度的增加而显著增加(P<0.05),不同放牧处理比不放牧的碳含量增加了7.97%~11.06%,G0.46、G0.69和G0.92之间无显著性差异(P>0.05),显著高于不放牧G0.00和G0.23(P<0.05);二裂委陵菜的碳含量呈现先增加再降低的趋势,在放牧强度G0.46时到达峰值,高达383.93 g·kg-1,显 著 高 于G0.69和G0.92(P<0.05),G0.00、G0.23、G0.34和G0.46之间无显著性差异(P>0.05);贝加尔针茅、寸草苔、星毛委陵菜的碳含量在不同放牧强度之间无显著性差异(P>0.05)。

图2 不同放牧强度下植物碳含量变化
6种植物的全氮含量均随着放牧强度的增加而显著增加(P<0.05)(图3)。优势种羊草和贝加尔针茅G0.46、G0.69和G0.92均 显 著 高 于G0.00(P<0.05),中度-重度放牧羊草和贝加尔针茅平均增长率分别达47.96%和76.87%,G0.23、G0.34、G0.46以 及G0.69相互之间无显著性差异(P>0.05);寸草苔和二裂委陵菜的全氮含量G0.92显著高于G0.00(P<0.05),增长率分别达到25.66%和23.75%;退化指示种冷蒿和星毛委陵菜G0.69和G0.92显著高于G0.00、G0.23和G0.34,最大差值分别为4.25和5.40 g·kg-1,全氮含量显著增加了21.78%~29.00%。

图3 不同放牧强度下植物全氮含量变化
6种植物的全磷含量随着放牧强度的变化有不同的趋势(图4)。优势种羊草和贝加尔针茅全磷含量均随着放牧强度的增加而增加,G0.69和G0.92之间无显著性差异(P>0.05),均显著高于G0.00、G0.23以及G0.34(P<0.05),与不放牧相比,羊草和贝加尔针茅不同放牧强度下全磷含量分别增加了6.27%~68.54%和10.35%~87.43%;寸草苔和退化种冷蒿、星毛委陵菜、二裂委陵菜全磷含量在不同放牧强度之间无显著性差异(P>0.05),总体从数值上呈现中度-重度放牧增加了植物全磷含量。

图4 不同放牧强度下植物全磷含量变化
2.2 不同放牧强度植物化学计量学特征变化
6种植物的碳氮比、碳磷比和氮磷比随着放牧强度的变化呈现不同的变化趋势(表1)。优势种羊草和建群种贝加尔针茅的碳氮比和碳磷比随着放牧强度的增加而显著下降。其中,羊草均呈现G0.00、G0.23和G0.34显著高于G0.69和G0.92(P<0.05),与不放牧相比,放牧下羊草的碳氮比和碳磷比分别下降了9.97%~37.28%、4.09%~45.16%,氮磷比不同放牧强度之间无显著性差异,但也呈现在重度放牧降低的趋势;建群种贝加尔针茅碳氮比和碳磷比呈现与羊草相同的变化,随着放牧强度的增加而显著下降,碳氮比呈现G0.00和G0.23显著高于G0.92(P<0.05),碳磷比呈现出G0.00显著高于G0.69和G0.92(P<0.05),G0.00分别是G0.69和G0.92的2.14和2.24倍,氮磷比在不同放牧强度之间无显著性差异。放牧同样降低了退化指示种寸草苔、冷蒿、星毛委陵菜和二裂委陵菜的碳氮比,寸草苔G0.00显著高于G0.92(P<0.05),星毛委陵菜和二裂委陵菜呈现G0.00显著高于G0.69和G0.92(P<0.05),冷蒿不同处理之间差异未达到显著性水平(P>0.05);对于碳磷比和氮磷比,退化种寸草苔、冷蒿、星毛委陵菜和二裂委陵菜在不同放牧强度之间无显著性差异(P>0.05),但总体呈现重度放牧降低了物种寸草苔、冷蒿、星毛委陵菜和二裂委陵菜的碳磷比以及寸草苔的氮磷比,增加了退化指示种的氮磷比。

表1 不同放牧强度下不同植物化学计量特征
2.3 不同放牧强度植物碳、氮、磷贮量变化
6种植物的碳贮量呈现不同的变化趋势(图5)。不同放牧强度下碳贮量变化范围分别是羊草0.667~48.489 g·m-2、贝加尔针茅0.725~2.059 g·m-2、寸草苔0.932~4.717 g·m-2、冷蒿0.095~7.553 g·m-2、星毛委陵菜0.032~1.786 g·m-2、二裂委陵菜0.644~2.505 g·m-2。优势物种羊草的碳贮量随着放牧强度的增加呈现显著的线性下降趋势(R2=0.71,P<0.05),G0.00和G0.23显著高于其他放牧强度(P<0.05),与不放牧相比,中度-重度放牧碳贮量降低了87.10%~98.41%;退化指示种冷蒿(R2=0.64,P<0.05)和星毛委陵菜(R2=0.97,P<0.001)的碳贮量均随着放牧强度的增加呈现显著线性增加,冷蒿G0.69显著高于G0.00、G0.23、G0.34和G0.46(P<0.05),星毛委陵菜G0.69显著高于G0.00、G0.23、G0.34和G0.46(P<0.05),二者极重度放牧的碳贮量分别是不放牧的46.13和55.51倍;寸草苔碳贮量在各放牧强度间无显著差异(P>0.05),但呈现显著线性回归(R2=0.96,P<0.001),总体从数值上呈现中度-重度放牧增加了植物碳贮量,与不放牧相比含量增加了0.67~3.06倍;贝加尔针茅(R2=0.48,P>0.05)和二裂委陵菜(R2=0.56,P>0.05)的碳贮量随着放牧强度的增加呈现二项式变化,均在中度放牧G0.46最高,分别达到2.059和2.505 g·m-2。
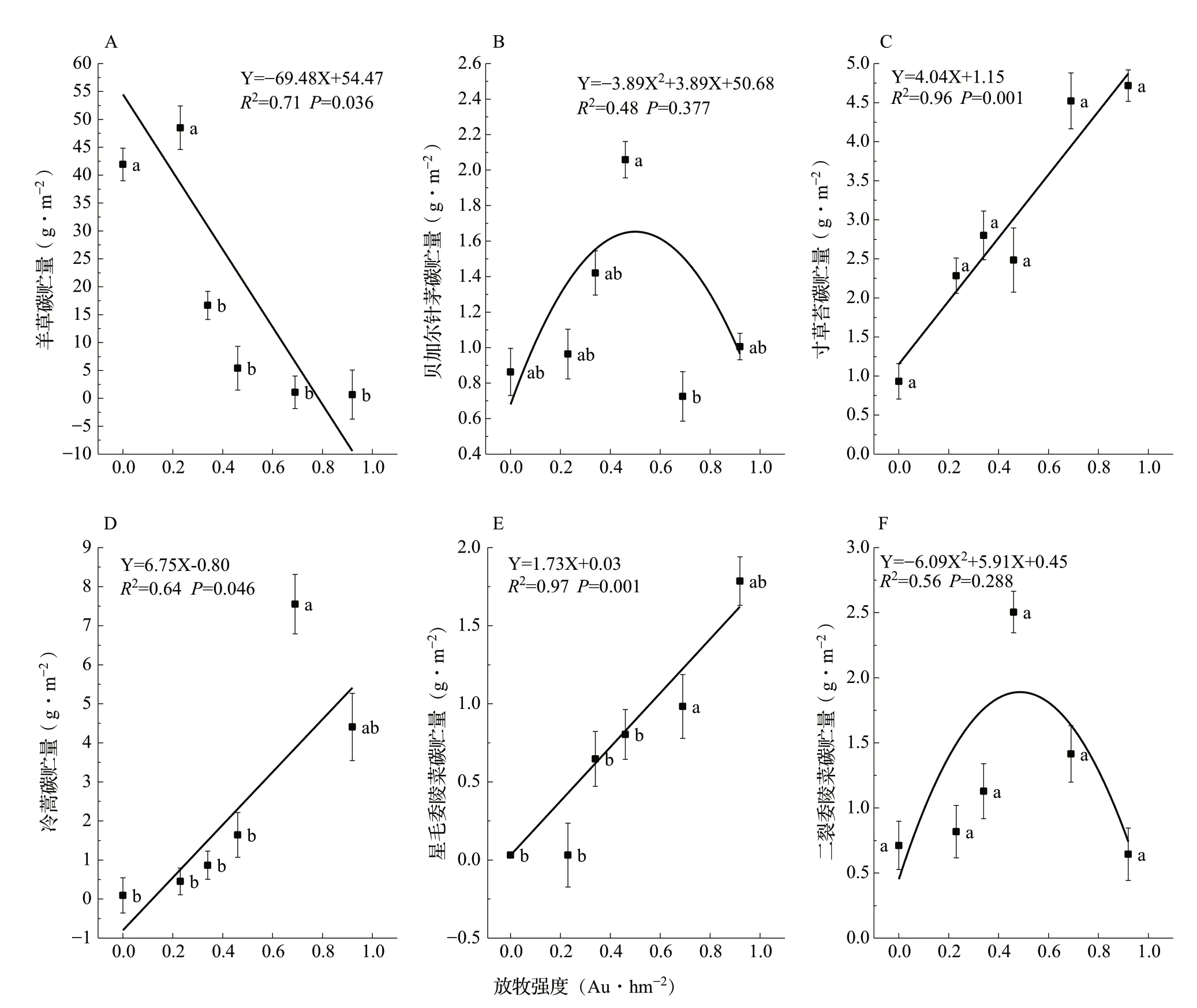
图5 不同放牧强度下植物碳贮量的变化
不同放牧强度下植物氮贮量的变化范围分别是羊草0.045~2.292 g·m-2、贝加尔针茅0.028~0.093 g·m-2、寸草苔0.046~0.305 g·m-2、冷蒿0.005~0.437 g·m-2、星毛委陵菜0.002~0.141 g·m-2、二裂委陵菜0.035~0.151 g·m-2。优势物种羊草的氮贮量随着放牧强度的增加呈现显著的线性下降趋势(R2=0.68,P<0.05),G0.00和G0.23显著高于G0.46、G0.69和G0.92(P<0.05),与不放牧相比,中度-重度放牧氮贮量降低了82.01%~97.55%;退化指示种冷蒿(R2=0.69,P<0.05)和星毛委陵菜(R2=0.80,P<0.05)的氮贮量都是随着放牧强度的增加呈现显著线性增加,冷蒿G0.69显著高于G0.00、G0.23、G0.34和G0.46(P<0.05),星毛委陵菜G0.92显著高于其他放牧强度(P<0.05),二者重度放牧的氮贮量是分别不放牧的48.00和66.19倍;寸草苔氮贮量在各放牧强度间无显著差异(P>0.05),但呈现显著线性回归(R2=0.98,P<0.001),总体从数值上呈现出中度-重度放牧增加了寸草苔氮贮量,与不放牧相比含量增加了1.18~4.68倍;贝加尔针茅(R2=0.30,P>0.05)和二裂委陵菜(R2=0.44,P>0.05)的氮贮量随着放牧强度的增加呈现二项式变化,均在中度放牧G0.46最高,分别达到0.093和0.151 g·m-2(图6)。

图6 不同放牧强度下植物氮贮量的变化
不同放牧强度下植物磷贮量的变化范围分别是羊草0.004~0.159 g·m-2、贝加尔针茅0.002~0.005 g·m-2、寸草苔0.003~0.022 g·m-2、冷蒿0.000~0.043 g·m-2、星毛委陵菜0.000~0.013 g·m-2、二裂委陵菜0.003~0.010 g·m-2。优势物种羊草的磷贮量随着放牧强度的增加呈现显著的线性下降趋势(R2=0.72,P<0.05),G0.00和G0.23显著高于G0.46、G0.69和G0.92(P<0.05),与不放牧相比,中度-重度放牧磷贮量降低了82.33%~97.41%;退化指示种寸草苔(R2=0.94,P<0.05)、冷蒿(R2=0.75,P<0.05)和星毛委陵菜(R2=0.93,P<0.05)的磷贮量都是随着放牧强度的增加呈现显著线性增加,寸草苔极重度放牧显著高于不放牧(P<0.05),冷蒿G0.69显著高于G0.00、G0.23、G0.34和G0.46(P<0.05),星毛委陵菜G0.92显著高于G0.00和G0.23(P<0.05),三者重度放牧的磷贮量分别比不放牧增加了1.38~6.69、15.67~45.67和25.6~64倍;贝加尔针茅(R2=0.48,P>0.05)和二裂委陵菜(R2=0.68,P>0.05)的磷贮量随着放牧强度的增加呈现二项式变化,均在中度放牧G0.46最高,分别达到0.005和0.010 g·m-2(图7)。

图7 不同放牧强度下植物磷贮量的变化
3 讨论
3.1 放牧强度对养分含量变化的影响
在植物生长发育的过程中,碳、氮、磷作为最基本的元素,当外界条件发生变化时,在植物体内重新分配以抵御逆境[19]。Li等[20]认为放牧使叶片和茎组织中的碳含量显著升高。有研究表明,碳含量降低的主要原因是随放牧强度的增加,动物的采食和践踏导致衰老部位存在的比重逐渐下降,再生幼叶所占比例相对较多,进而导致碳含量降低[21]。本研究结果表明,优势物种羊草的碳含量随着放牧强度的变化显著降低,而退化指示种冷蒿的碳含量却显著增加。造成这一结果的原因是羊草作为优势物种是草甸草原肉牛的优良牧草之一,随着牛采食量的增大,在叶子生长初期不能进行固碳功能,所以嫩叶中碳含量较低,地上部分的碳含量也较低;而冷蒿低矮、避牧性强,采食量较低,老叶比例较高导致种群碳含量较大[22]。6种植物的氮含量都随着放牧强度的增加显著上升,而植物体中的氮、磷是协同元素,一般呈正相关关系[11],磷含量整体有上升趋势,其中羊草和贝加尔针茅显著增加,这与Baron等[23]的研究结果基本一致,该研究表明草食动物导致草原植物组织氮和磷的含量增加。造成这一结果的一个原因是放牧导致植物被采食的程度逐步加深,为了抵抗胁迫,增加对光资源的利用,植物补偿性生长出新的嫩叶来进行光合作用[24-26],植株体内养分再循环,将氮和磷输送到嫩叶中,使之含有较多的氮和磷元素[27];另一个原因是随着放牧强度的增加,牛排泄的粪便和尿液也随之增加,这就会促进土壤中的氮和磷循环,植物吸收的元素量也可能会相应提高;另外,牛的践踏会导致土壤理化性质发生改变,影响根系对土壤氮和磷的吸收,最终使植物的氮和磷含量发生变化。李金花等[28]研究发现,冷蒿与星毛委陵菜虽都为退化指示植物,但其遗传特性不同,在同一放牧试验中随着放牧强度增加,冷蒿叶表现为氮含量下降,而星毛委陵菜地上部分与之相反,氮含量增加。从本研究结果可以看出,优势物种羊草和贝加尔针茅的磷含量有显著增长,而其他4种不显著,可以判断不同遗传性状的植物在同一条件下,元素在植物体内的含量的确会有差别,优势物种对磷的吸收能力更强一些。
3.2 放牧强度对化学计量特征变化的影响
不同放牧强度在不同植物群落中的碳、氮、磷生态化学计量特征影响不同[29],而植物叶片碳氮比和碳磷比意味着植物吸收营养所能同化碳的能力,在一定程度上反映了植物的营养利用效率[30]。丁小慧等[7]和翟夏杰等[22]的研究结果指出,碳氮比越低,说明植物生长越迅速。放牧活动中家畜的采食减少了植物部分衰老组织,促进植物进行补偿性生长,导致植物对于营养生长的需求越来越迫切,所以6种植物的碳氮比均降低,试验结果与上述规律一致。LI等[20]的研究结果表明,碳磷比与放牧压力呈显著负相关,主要是由于随放牧强度的增加,动物的采食践踏导致再生幼嫩器官所占比例相对较多,但其碳含量较低,磷含量较高,进而导致碳磷比降低,这与本研究结果吻合。生长速率假说认为生长速率较高的生物,其碳氮比、碳磷比和氮磷比较低[31-32],本试验研究结果与之有差异的一点是,不同植物氮磷比对放牧强度的响应有差异性,羊草、贝加尔针茅、寸草苔的氮磷比随着放牧强度的增加呈现降低的趋势,但星毛委陵菜和二裂委陵菜呈现上升的趋势,这表明不同植物对养分的利用策略是不同的。造成这一结果的原因可能是生长速率假说是针对细菌和浮游生物的研究得到的结论,植物与细菌和浮游生物不同,对于植物化学计量学的研究更为复杂,造成了氮和磷的转移速率不一致[33]。Koerselman等[34]的试验研究结果表明,当植物氮磷比小于14时,表现为受氮的限制;当氮磷比大于16时,表现为受磷的限制。综合中国草原优势植物的生态化学计量分析结果后发现,中国不同类型草原其生态化学计量学特征有所差异,但总体差别较小,中国草原生态系统的氮磷比高于全球陆地生态系统的平均值(12.6),数据表明与世界其他地区相比,中国草地生态系统缺磷现象比较明显[35]。本试验结果表明,羊草、冷蒿、星毛委陵菜和二裂委陵菜在各个放牧强度下,都表现为受到氮的限制,寸草苔在重度放牧的条件下也表现为受到氮的限制。与重度放牧区相比较,不放牧、轻度放牧以及中度放牧区的贝加尔针茅受到磷的限制。这与上述结果不同,可能是与植物的生活习性和不同的遗传特性有关系[14]。
3.3 放牧强度对植物碳、氮、磷元素贮量变化的影响
本研究中,并非所有植物的元素贮存趋势都相同,植物特性不同,变化规律也不相同,体现了种间特异性。董晓玉等[15-16]的研究表明,草地植物碳、氮、磷元素贮量均与生物量呈显著正相关关系,即植物元素贮存由元素含量和地上生物量共同决定,但碳、氮、磷的贮量受元素含量影响不大,这些变化都与地上生物量的变化密切相关。放牧对草地的影响首先从地上生物量体现出来[36-37],随着放牧强度的增加,优势物种生物量降低,退化物种生物量增加[38]。本试验结果显示,随着放牧强度的增加,适口性较好的羊草,受放牧影响较大,被大量采食导致地上生物量减少,使呼伦贝尔草甸草原以羊草为代表的优势物种的碳、氮、磷贮量逐渐降低,而贝加尔针茅、寸草苔、冷蒿、星毛委陵菜和二裂委陵菜则与之相反,地上生物量增加使其碳、氮、磷贮量逐步增加。此外,植物碳、氮、磷元素贮量的变化不仅与地上生物量关系密切,还受到放牧的影响,牛的采食不仅促进植物的再生长,还促进氮和磷向地上转移。仲延凯等[39]对内蒙古典型草原的研究表明,刈割后的植物体内氮和磷含量升高,因为这两种元素易向植物生长发育的器官转移,经过不同的放牧处理,牛对植物的采食、践踏程度越大,越刺激植物的再生长,新生幼嫩叶片越多,转移的氮和磷也就越多,这与本研究的结果一致。因此,放牧主要是通过地上幼嫩部分生物量和植物体内碳、氮、磷含量来控制碳、氮、磷贮量,若想获得较大的碳、氮、磷贮量则需要控制放牧强度,通过适度放牧来提高草地的化学计量学平衡与管理水平。本研究依托的大型控制放牧实验平台已经进行了10余年的放牧试验,受到自然条件及其他因素的共同影响,且放牧对于生态系统的影响具有长远性,放牧对草地生态系统化学计量学及贮量的影响值得进一步研究。
4 结论
放牧增加了退化指示种冷蒿的碳含量,降低了优势种羊草的碳含量;中度-重度放牧增加了6种植物的全氮含量以及优势种羊草和建群种贝加尔针茅的全磷含量。
放牧降低了优势种羊草和建群种贝加尔针茅的碳氮比和碳磷比,重度放牧降低了物种寸草苔、冷蒿、星毛委陵菜和二裂委陵菜的碳氮比、碳磷比以及寸草苔的氮磷比,增加了退化指示种的氮磷比。
羊草、冷蒿、星毛委陵菜和二裂委陵菜受到氮的限制,寸草苔在重度放牧的条件下表现为受到氮的限制,不放牧、轻度放牧以及中度放牧区的贝加尔针茅受到磷的限制。
放牧显著降低了优势物种羊草的碳、氮、磷贮量,增加了退化指示种冷蒿和星毛委陵菜的碳、氮、磷贮量,贝加尔针茅和二裂委陵菜的碳、氮、磷贮量均呈现中度放牧最高,符合中度放牧优化假说。
