祁连山典型灌丛植物根系化学计量特征及其与土壤理化因子的关系①
2022-05-09赵维俊赵永宏敬文茂武秀荣贺永岩陈丽英
马 剑,冯 起,赵维俊,赵永宏,敬文茂,武秀荣,贺永岩,陈丽英
祁连山典型灌丛植物根系化学计量特征及其与土壤理化因子的关系①
马 剑1,2,3,冯 起2,赵维俊1,3,赵永宏4,敬文茂1, 3,武秀荣1,3,贺永岩1,3,陈丽英1,3
(1 甘肃省祁连山水源涵养林研究院,甘肃张掖 734000;2 中国科学院西北生态环境资源研究院,兰州 730000;3 甘肃祁连山森林生态监测与评估国际科技合作基地,甘肃张掖 734000;4 三峡大学水利与环境学院,湖北宜昌 443002)
为了揭示植物根系养分含量及其化学计量特征与土壤因子之间的关系,选择祁连山大野口流域的甘青锦鸡儿()、鲜黄小檗()、金露梅()、鬼箭锦鸡儿()和吉拉柳()等5种典型灌丛为研究对象,研究了植物粗根、细根化学计量特征及其与土壤理化因子之间的关系。结果表明,粗根碳、氮、磷平均含量分别为569.55、7.86、1.02 g/kg,细根分别为541.38、8.61、1.10 g/kg,其中磷变异系数较高,碳变异系数最小;粗根C/N、C/P、N/P均值分别为86.63、729.78、9.77,细根分别为71.34、527.61、8.11;碳、磷含量高于中国植物根系平均水平,粗根氮含量高于中国植物粗根的平均水平,但细根氮含量低于中国植物细根的平均水平。土壤有效磷含量和质量含水量是影响粗根生态化学计量特征的主要因子,土壤有效磷含量和pH是影响细根生态化学计量特征的主要因子。
典型灌丛;粗根;细根;化学计量;祁连山
生态化学计量学是研究生态过程中能量和化学元素平衡的科学,为解决生态系统养分供需和循环问题提供了一条新的途径[1],其目的在于定量研究生物体内碳(C)、氮(N)、磷(P)之间的关系[2]。碳、氮、磷是植物生长所必需的元素[3],影响着物种组成和代谢过程[4]。碳、氮是促进植物新陈代谢过程和植物体构建的重要元素[5],与磷元素共同参与了碳水化合物、酶、纤维素等代谢过程[6]。C/N、C/P作为植物生长的重要指标,间接反映了植物对营养元素的利用效率[7],N/P是反映群落结构及功能的重要指标[8],其变化规律有助于深入了解一定区域的养分限制情况和植物的适应策略[9]。因此,研究植物碳、氮、磷含量及其化学计量特征,对认识生态系统养分耦合循环和植被生长机制具有重要的意义。近年来,国内外生态化学计量学的研究对象多集中于森林、草原、荒漠等生态系统[10-12],较少关注灌丛生态系统;研究内容多聚焦于地上部分,认为碳、氮、磷含量及不同的化学计量特征影响着植物的生长[13]。根系是连接土壤和植物的纽带,为植物的生长提供所需水分和养分,其养分含量在一定程度上可反映植物生长过程中的养分需求状况,揭示土壤养分的供给特征[14]。然而由于植物根系的分布较为复杂且难以获得,有关植物根系养分含量及其化学计量特征的报道相对较少,这在一定程度上限制了土壤–植物分布关系和养分循环的动态研究。
祁连山是中国西部重要的生态安全屏障,是黄河流域重要水源产流地,也是中国生物多样性保护优先区域。灌丛群落作为祁连山森林生态系统的主要组成部分,对祁连山森林系统保护水土、涵养水源以及维持河西走廊生态系统和保护我国生物多样性起着极为重要的作用[15]。以祁连山区灌丛群落中的优势灌木植物为研究对象,研究其根系养分含量及其化学计量特征,不仅可以深入了解生态脆弱区植物生长的养分限制情况,还可揭示祁连山区灌丛群落在严酷环境条件下的生存策略。然而,目前对祁连山区灌丛群落植物营养器官养分含量特征的研究较少,灌丛植物如何调整根系的化学计量特征来适应高寒干旱环境?根系化学计量特征与土壤因子之间的关系如何?这些问题尚不清楚,一定程度上限制了对灌丛生态系统养分循环过程的认识。为此,本研究选择祁连山北麓中段典型流域——大野口流域不同垂直植被带上的甘青锦鸡儿()、鲜黄小檗()、金露梅()、鬼箭锦鸡儿()和吉拉柳()等5种典型灌丛群落为研究对象,分析灌丛植物粗根和细根碳、氮、磷含量及其生态化学计量的变化特征,探讨其生态化学计量特征与土壤理化因子之间的关系,旨在揭示祁连山区灌丛植物生存策略,以期为干旱、半干旱地区植物生态化学计量特征的探究以及祁连山植被的恢复与保护提供理论依据。
1 材料与方法
1.1 研究区概况
祁连山位于青藏高原、蒙古高原和黄土高原的交界处,是黑河、疏勒河和石羊河的水源地,对区域经济社会发展和生态安全具有重要意义[16]。祁连山区作为典型的高寒半干旱山区,具有海拔跨度大(1 700 ~ 5 800 m)、水热梯度显著以及对气候变化响应敏感等特点[17]。独特的生态环境以及较少的人为干扰,使得该区成为研究山区植被生态化学计量特征的理想区域。大野口流域位于祁连山北麓中段的西水林区,中心位置100°15′E、38°31′N,流域总面积73.32 km2,流域海拔2 590 ~ 4 645 m[17]。年平均气温–0.6 ~ 2.0 ℃,年均降水量为433.6 mm,年均蒸发量为1 081.7 mm,年均相对湿度为60%,属高寒半干旱山地森林草原气候[18]。植被类型主要有森林草原带、森林灌丛带、亚高山灌丛草甸带、高山草甸带;土壤类型主要有山地灰钙土、山地栗钙土、山地灰褐土、亚高山灌丛草甸土、高山寒漠土[19]。
1.2 野外采样
基于典型性、代表性原则,选择研究区的甘青锦鸡儿、鲜黄小檗、金露梅、鬼箭锦鸡儿、吉拉柳等5种典型灌丛群落,依据植被调查中比较常用的典型样地法[20],在每种灌丛类型内建立3个面积为20 m×20 m的调查样地,共设置15个样地,样地情况见表1。2019年8月对每个群落优势种(甘青锦鸡儿、鲜黄小檗、金露梅、鬼箭锦鸡儿、吉拉柳)根系和土壤进行样品采集,每个样品均采集3个重复。依据灌丛植物的生长情况,在样地内随机选择5 ~ 10株(丛)长势相近、生长良好的灌丛植株,用铁锹连根系挖出,尽快带回实验室。
在每个样地内采用土钻法和剖面法结合的方式采集土壤样品。由于0 ~ 10 cm土层土壤理化性质变化较为强烈,因此我们选择0 ~ 10 cm土层数据来分析植物根系化学计量特征与土壤属性之间的关系,在样地内选择样地两条对角线的交点处挖一个土壤剖面,首先对土壤剖面颜色、质地等信息进行调查记录,然后用体积为200 cm3的环刀采集0 ~ 10 cm土层样品,采集好的样品装入提前编好号的自封袋;同时,在每个样地内采用五点法,用直径2 cm的土钻采集0 ~ 10 cm土层混合土样1份,剔除石砾、植物残根等杂物,混合均匀,四分法取约1 kg,装入预先写好编号的自封袋,带回实验室。
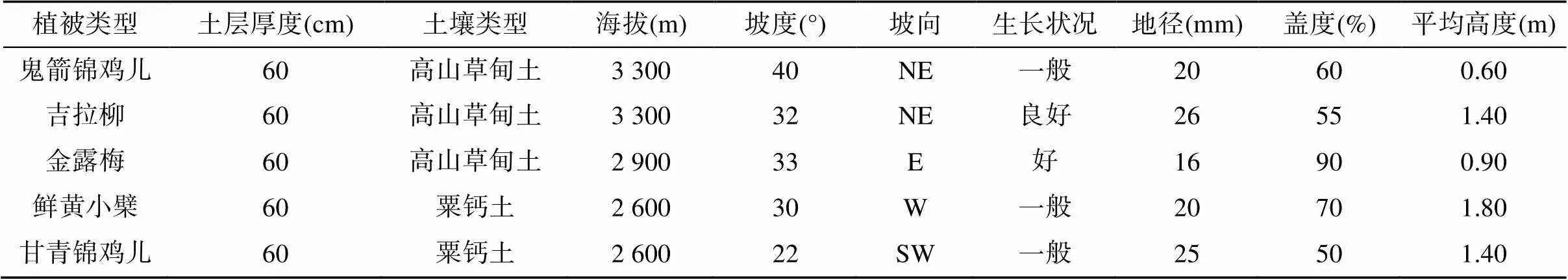
表1 样地基本情况
1.3 试验方法
将采集的植物样品,在实验室用自来水冲洗1 ~ 2次,再用蒸馏水快速冲洗2 ~ 3次,晾干后参考前人的研究方法[21-23],用游标卡尺准确测量根系的直径,按照0 ~ 2 mm、>2 mm直径范围将根系分成细根和粗根,将晾干的细根和粗根置于105 ℃烘箱杀青2 h,然后80 ℃烘干至恒重,用粉碎机将样品粉碎后过100目筛,装密封袋,用于测定植物叶片全碳、全氮、全磷含量。其中碳含量采用重铬酸钾容量法-外加热法测定,氮含量采用凯氏定氮法测定,磷含量采用钼锑抗比色法测定[24]。
将环刀采集的土样,用于测定土壤pH、质量含水量、容重、毛管孔隙度等指标,土壤pH采用电位法测定,土壤质量含水量、容重、毛管孔隙度度采用烘干法测定[25];总孔隙度用t=93.947–32.995×来计算,为容重,t为总孔隙度;非毛管孔隙度用o=t–c计算,c为毛管孔隙度,o为非毛管孔隙度。土钻采集的土样,经风干、过筛、用于测定土壤有机碳、全氮、全磷、碱解氮、有效磷等指标。土壤有机碳含量采用重铬酸钾容量法测定,全氮含量采用凯氏定氮法测定,全磷含量采用钼锑抗比色法测定,碱解氮含量采用碱解扩散法测定,有效磷采用钼锑抗比色法测定[24]。
1.4 数据分析
植物粗根和细根碳、氮、磷含量为质量含量,C/N、C/P和N/P 均采用质量比。采用 SPSS 20.0 对细根和粗根碳、氮、磷特征进行描述性统计;采用 SAS 9.2 中的一般线性模型(General Linear Model)分析检验不同灌丛细根和粗根碳、氮、磷、C/N、C/P、N/P 的差异显著性,多重比较用新复极差法(Duncan),显著性水平设为 0.05。本研究将细根、粗根化学计量特征视为研究对象,以土壤pH、质量含水量、容重、总孔隙度、毛管孔隙度、非毛管孔隙度以及土壤有机碳、全氮、全磷、碱解氮、有效磷含量为环境变量,采用冗余分析法(RDA)分析细根、粗根生态化学计量特征与环境变量之间的关系。
2 结果与分析
2.1 植物根系碳、氮、磷元素的基本特征
5种典型灌丛植物粗根的碳、氮、磷含量均值分别为569.55、7.86、1.02 g/kg,C/N、C/P、N/P均值分别为86.63、729.78、9.77;细根的碳、氮、磷含量均值分别为541.38、8.61、1.10 g/kg,C/N、C/P、N/P均值分别为71.34、527.61、8.11(表2)。5种灌丛植物根系化学计量特征变异程度如表2所示,粗根磷含量变异系数最高(66%),N/P变异系数次之(59%),碳含量变异系数最小(5%);细根N/P变异系数最高(38%),C/N变异系数次之(37%),碳含量变异系数最小(4%)。粗根氮含量与C/N(2=0.97,<0.01)、N/P(2=0.38,<0.05)均呈显著二次函数关系;磷含量与C/P(2=0.96,<0.01)呈极显著二次函数关系,与N/P(2=0.47,<0.05)呈显著二次函数关系(图1)。细根氮含量与C/N(2=0.99,<0.01)呈极显著二次函数关系,与N/P(2=0.42,<0.05)呈显著二次函数关系;磷含量与C/P(2=0.98,<0.01)呈极显著二次函数关系(图2)。对粗根和细根来说,碳与其他化学计量特征均没有显著关系,且氮、磷之间也未表现出显著相关性。
2.2 不同灌丛类型根系碳、氮、磷化学计量特征的差异性
5种灌丛植物根系碳、氮、磷含量如表3所示,不同灌丛类型粗根碳含量表现为金露梅>吉拉柳>鲜黄小檗>鬼箭锦鸡儿>甘青锦鸡儿,氮含量表现为甘青锦鸡儿>鬼箭锦鸡儿>鲜黄小檗>金露梅>吉拉柳,磷含量表现为鬼箭锦鸡儿>金露梅>吉拉柳>甘青锦鸡儿>鲜黄小檗。碳含量在不同灌丛类型间差异不显著,氮含量在甘青锦鸡儿与其他灌丛类型之间差异显著(<0.05),磷含量在鬼箭锦鸡与其他灌丛间差异显著(<0.05)。不同灌丛类型细根碳含量表现为鲜黄小檗>吉拉柳>金露梅>鬼箭锦鸡儿>甘青锦鸡儿,氮含量表现为甘青锦鸡儿>鬼箭锦鸡儿>鲜黄小檗>金露梅>吉拉柳,磷含量表现为鬼箭锦鸡儿>金露梅>吉拉柳>鲜黄小檗>甘青锦鸡儿。碳含量在不同灌丛类型间差异不显著;氮含量在甘青锦鸡儿与鬼箭锦鸡儿之间差异不显著,与其他灌丛类型之间差异显著(<0.05);磷含量在鬼箭锦鸡与金露梅之间差异不显著,与其他灌丛间差异显著(<0.05)。

表2 根系碳、氮、磷化学计量特征的描述性统计

图1 粗根碳、氮、磷化学计量特征的相关性

图2 细根碳、氮、磷化学计量特征的相关性

表3 不同灌丛类型根系碳、氮、磷化学计量特征的多重比较
注:表中同行小写字母不同表示植物粗根或细根碳、氮、磷含量及化学计量比在不同灌丛类型之间存在显著差异(<0.05)。
由表3可以看出,粗根C/N在不同灌丛类型间为吉拉柳>金露梅>鲜黄小檗>鬼箭锦鸡儿>甘青锦鸡儿,C/P为鲜黄小檗>甘青锦鸡儿>吉拉柳>金露梅>鬼箭锦鸡儿,N/P为甘青锦鸡儿>鲜黄小檗>金露梅>吉拉柳>鬼箭锦鸡儿。C/N在吉拉柳与其他灌丛类型间差异显著(<0.05),在甘青锦鸡儿、鲜黄小檗、鬼箭锦鸡儿之间差异不显著;C/P在鬼箭锦鸡儿与其他灌丛间差异显著(<0.05),但在其他灌丛之间差异不显著;N/P在甘青锦鸡儿与其他灌丛间差异显著(< 0.05)。细根C/N在不同灌丛类型间为吉拉柳>金露梅>鲜黄小檗>鬼箭锦鸡儿>甘青锦鸡儿,C/P为鲜黄小檗>甘青锦鸡儿>吉拉柳>金露梅>鬼箭锦鸡儿,N/P为甘青锦鸡儿>鲜黄小檗>鬼箭锦鸡儿>金露梅>吉拉柳。C/N在吉拉柳与其他灌丛类型间差异显著(<0.05),在甘青锦鸡儿、鬼箭锦鸡儿之间差异不显著;C/P在鬼箭锦鸡儿与吉拉柳、甘青锦鸡儿、鲜黄小檗间差异显著(<0.05);N/P在甘青锦鸡儿与其他灌丛间差异显著(<0.05)。
2.3 植物根系生态化学计量特征与土壤理化因子的冗余分析
对5种灌丛植物根系碳、氮、磷化学计量特征与土壤理化因子进行冗余分析,得出粗根、细根生态化学计量特征在第Ⅰ轴的解释量分别为86.82%、85.74%,第Ⅱ 轴的解释量分别为1.51%、3.95%,第Ⅲ 轴与第Ⅳ 轴解释量的和分别仅为0.04%、1.69%,且前两轴分别共解释粗根、细根生态化学计量特征88.34%、89.70%,对植物粗根、细根生态化学计量特征和土壤理化因子关系的累计解释量分别为99.53%、99.90%,由此表明,前两轴可很好地反映植物根系生态化学计量特征与土壤理化因子之间关系,且主要决定于第Ⅰ轴。
图3为根系生态化学计量特征与土壤因子关系的二维排序图。由图3粗根冗余分析排序图可以看出,土壤有效磷含量和质量含水量的箭头连线最长,表明土壤有效磷含量和含水量对粗根生态化学计量特征变异起到了很好的解释。土壤有效磷含量与粗根碳、氮、磷含量呈正相关,与粗根C/N、C/P、N/P负相关,土壤质量含水量与粗根碳、磷含量、C/N正相关,与氮含量、C/P、N/P负相关,其中土壤有效磷含量、质量含水量均与N/P的相关性最小。土壤碳、氮含量与粗根碳、磷含量、C/N呈正相关,与粗根氮含量、C/P、N/P负相关;土壤磷含量与C/N、C/P、N/P呈正相关,与碳、氮、磷含量呈负相关。由图3细根冗余分析排序图可以看出,土壤有效磷含量和土壤pH的箭头连线最长,表明土壤有效磷含量和土壤pH对细根生态化学计量特征变异起到了很好的解释。土壤有效磷含量与细根碳、氮、磷含量呈正相关,与细根C/N、C/P、N/P负相关;土壤pH与细根氮含量、C/P、N/P正相关,与细根碳、磷含量和C/N呈负相关。土壤碳含量与细根碳、磷含量和C/N呈正相关,与氮含量、C/P、N/P呈负相关;土壤氮含量与细根碳、C/N、C/P呈正相关,与细根氮、磷含量和N/P负相关;土壤磷含量与细根C、C/N、C/P、N/P正相关,与细根氮、磷含量呈负相关。

(A. 根碳含量;B. 根氮含量;C. 根磷含量;D. 根C/N;E. 根C/P;F. 根N/P;a. 土壤碳含量;b. 土壤氮含量;c. 土壤磷含量; d. 土壤碱解氮含量;e. 土壤有效磷含量;f. 土壤pH;g. 土壤容重;h. 土壤毛管孔隙度;i. 土壤非毛管孔隙度;j. 土壤总孔隙度; k. 土壤质量含水量)
表4为土壤理化因子解释量与显著性检验,对粗根而言,其解释量大小依次为土壤有效磷含量、质量含水量、总孔隙度、容重、氮含量、非毛管孔隙度、碳含量、毛管孔隙度、磷含量、碱解氮含量、pH,其中土壤有效磷含量、质量含水量对粗根的影响较为显著(<0.05),占土壤因子解释量的40.2% 和18.5%,说明土壤有效磷含量和质量含水量是影响粗根生态化学计量特征的主要因素;其他土壤理化因子对其影响均未达到显著水平。对细根而言,其解释量大小依次为土壤有效磷含量、pH、总孔隙度、容重、碳含量、氮含量、非毛管孔隙度、磷含量、碱解氮含量、毛管孔隙度、质量含水量,其中土壤有效磷含量、pH对细根的影响较为显著(<0.05),两者分别占土壤因子解释量的33.2% 和20.5%,说明土壤有效磷含量和pH是影响细根生态化学计量特征的主要因素;其他土壤理化因子对其影响均未达到显著水平。
3 讨论
3.1 根系碳、氮、磷化学计量特征
在长期自然选择中,植物通过优化资源配置,以适应不同环境所产生的胁迫,从而导致不同器官营养元素的差异[26]。本研究中,5种灌丛植物粗根碳含量(569.55 g/kg)、细根碳含量(541.38 g/kg)高于全球植物根系平均水平(535.58 g/kg)[27],也高于中国植物粗根碳含量(456.50 g/kg)和细根碳含量(473.90 g/kg),说明祁连山区灌丛植物根部碳储备能力较强。5种灌丛植物粗根氮含量(7.86 g/kg)高于中国植物粗根氮含量(5.9 g/kg),说明北方干旱半干旱区植物氮含量相对较高,也证实了高寒区较其他区域有较高氮含量的结论[28]。本研究中,灌丛植物粗根磷含量(1.02 g/kg)高于中国植物粗根磷含量(0.6 g/kg),而细根磷含量(1.10 g/kg)略高于中国植物细根磷含量(1.00 g/kg)。研究表明,由湿润地区到干旱半干旱地区,土壤磷含量呈现了增加趋势[29],说明我国干旱半干旱地区土壤磷含量相对较高,可能也是本研究中根系磷含量较高的原因。研究发现,灌丛植物根系中碳的变异系数最小,表现出了较高的稳定性,可能是因为碳元素在植物中主要起到骨架作用,不直接参与植物的生产活动;另外,植物主要通过光合作用吸收碳,使得植物体内的碳在一定范围内维持稳定状态。氮元素也相对稳定,对外界环境变化的响应不敏感[30];而磷元素的稳定性最低,是因为土壤磷含量差异性较大,导致根系磷含量不稳定。C/N、C/P反映了氮、磷利用效率和植物生长速率,N/P则反映植物受氮或磷的限制情况。本研究中,根的C/N值远远高于全球平均水平[27],表明研究区灌丛具有较高的元素利用效率。根的C/P和N/P值远远低于全球平均水平[27],可能与研究区寒冷的气候特征有关,根据温度-植物生理假说,低温有利于植物体内氮和磷的积累,且磷的积累速度更快[31],因而灌丛植物具有相对较低的C/P和N/P;同时也说明在区域尺度上根系生态化学计量特征变异较大。研究表明,N/P<14,植物生长受氮限制;当N/P>16,植物生长受磷限制;N/P介于14 ~ 16,植物生长受氮和磷的共同限制[32]。5种灌丛植物根系N/P均小于14,表明祁连山5种典型灌丛植物根的生长主要受氮的限制,这与赵维俊等[33]、张光德等[34]的研究结果相一致,其原因可能是祁连山灌丛植物根系具有较高的磷含量,导致根系的N/P较低,进而表现为氮限制。

表4 土壤因子解释量与显著性检验
由于灌丛植物种间的差异,其对环境的适应策略和资源的利用效率存在一定的差异[35],这种差异体现在碳、氮、磷化学计量特征上。碳含量在各灌丛类型间差异未达到显著水平,再一次说明碳在植物体内的骨架作用;氮含量在甘青锦鸡儿与鲜黄小檗、金露梅、吉拉柳之间差异显著(<0.05),磷含量在鬼箭锦鸡与甘青锦鸡儿、鲜黄小檗、吉拉柳之间差异显著(<0.05),表明不同灌丛植物对营养元素的吸收具有选择性。C/N在吉拉柳与其他灌丛类型间差异显著(<0.05),在甘青锦鸡儿、鬼箭锦鸡儿之间差异不显著;C/P在鬼箭锦鸡儿与吉拉柳、甘青锦鸡儿、鲜黄小檗间差异显著(<0.05);N/P在甘青锦鸡儿与其他灌丛间差异显著(<0.05),说明不同灌丛植物对养分适应策略不同,即对营养元素的利用效率及对环境的适应能力存在差异。
3.2 根系生态化学计量特征与土壤理化因子的关系
土壤与根系直接接触,是植物生命活动的重要场所,土壤理化特性对根系生态化学计量特征产生重大影响。本研究中,土壤有效磷含量是影响粗根、细根生态化学计量特征的主导因子,是因为植物磷主要来自根对土壤磷的吸收,土壤磷的有效性直接影响着植物根系对磷的吸收程度。土壤有效磷含量与根系磷含量呈正相关,与C/P、N/P负相关,表明随着土壤中有效磷含量的增加,灌丛植物根系磷含量增加,C/P、N/P减小。研究还发现,土壤质量含水量是影响粗根生态化学计量特征的主要因素,其中土壤质量含水量与粗根碳、氮、磷含量以及C/N、C/P、N/P存在显著相关关系,随着土壤水分的增加,粗根碳、氮、磷含量以及C/N、C/P、N/P呈现了一定的变化规律,说明土壤含水量的增加促进了祁连山区灌丛植物粗根对氮、磷元素的吸收。研究表明,植物体内的碳、氮等元素主要来自土壤,土壤养分和水分含量决定了植物吸收养分的量[36]。植物粗根中碳、氮化学计量特性与土壤水分状况密切相关,这与李玉霖等[37]、丁小慧等[38]、安申群等[39]得出的我国北方植物生态化学计量特征与土壤水分密切相关的结论一致。
本研究中,采用冗余分析方法研究土壤pH、质量含水量、总孔隙度、毛管孔隙度、非毛管孔隙度以及土壤有机碳含量、全氮含量、全磷含量、碱解氮含量、有效磷含量等对根系生态化学计量特征的影响,对揭示高寒半干旱山区植物的养分适应策略具有重要意义,但独立分析以上土壤理化因子对灌丛植物根系生态化学计量特征的影响存在一定的缺陷,土壤理化因子对根系生态化学计量特性的影响不是独立的,土壤理化因子之间也存在相互制约的关系。因此,在今后的研究中,应分析土壤理化因子对根系碳、氮化学计量特性的双重或多重影响,使研究结果更加准确、合理。
4 结论
研究区5种典型灌丛植物粗根碳、氮、磷含量均值分别为569.55、7.86、1.02 g/kg,细根分别为541.38、8.61、1.10 g/kg,其中磷变异系数较高,碳变异系数最小;粗根C/N、C/P、N/P均值分别为86.63、729.78、9.77,细根分别为71.34、527.61、8.11。碳、磷含量高于中国植物根系平均水平,粗根氮含量高于中国植物粗根的平均水平,但细根氮含量低于中国植物细根的平均水平。土壤有效磷含量和质量含水量是影响粗根生态化学计量特征的主要因素,土壤有效磷含量和pH是影响细根生态化学计量特征的主要因素。
[1] 江叶枫, 叶英聪, 郭熙, 等. 江西省耕地土壤氮磷生态化学计量空间变异特征及其影响因素[J]. 土壤学报, 2017, 54(6): 1527–1539.
[2] 陶冶, 吴甘霖, 刘耀斌, 等. 古尔班通古特沙漠典型灌木群落土壤化学计量特征及其影响因素[J]. 中国沙漠, 2017, 37(2): 305–314.
[3] 曹小玉, 李际平, 杨静, 等. 不同龄组杉木林土壤碳、氮、磷的生态化学计量特征[J]. 土壤, 2019, 51(2): 290– 296.
[4] 刘珮, 马慧, 智颖飙, 等. 9种典型荒漠植物生态化学计量学特征分析[J]. 干旱区研究, 2018, 35(1): 207–216.
[5] 刘颖, 贺静雯, 余杭, 等. 干热河谷优势灌木细根、粗根与叶片养分(C、N、P)含量及化学计量比[J]. 山地学报, 2020, 38(5): 668–678.
[6] 李晓菲, 李路, 常亚鹏, 等. 雪岭云杉林叶片碳氮化学计量特征及其与土壤理化因子的关系[J]. 干旱区地理, 2019, 42(3): 599–605.
[7] 赵洛琪, 吴晓妮, 付登高, 等. 滇池流域磷矿山区优势植物叶片与土壤养分生态化学计量特征[J]. 土壤, 2021, 53(2): 383–390.
[8] Vitousek P. Nutrient cycling and nutrient use efficiency[J]. The American Naturalist, 1982, 119(4): 553–572.
[9] 张小芳, 刘贤德, 敬文茂, 等. 祁连山不同海拔火绒草叶片生态化学计量特征及其与土壤养分的关系[J]. 应用生态学报, 2019, 30(12): 4012–4020.
[10] 秦燕燕, 李金花, 王刚, 等. 添加豆科植物对弃耕地土壤微生物多样性的影响[J]. 兰州大学学报(自然科学版), 2009, 45(3): 55–60.
[11] 仇瑶, 常顺利, 张毓涛, 等. 天山林区六种灌木生物量的建模及其器官分配的适应性[J]. 生态学报, 2015, 35(23): 7842–7851.
[12] 王晶苑, 王绍强, 李纫兰, 等. 中国四种森林类型主要优势植物的C︰N︰P化学计量学特征[J]. 植物生态学报, 2011, 35(6): 587–595.
[13] 肖遥, 陶冶, 张元明. 古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征[J]. 植物生态学报, 2014, 38(9): 929–940.
[14] 胡士达. 不同林龄闽楠人工林生态化学计量特征研究[D]. 长沙: 中南林业科技大学, 2017: 1–51.
[15] 王学福. 灌木林在祁连山区的作用及其发展策略研究[J]. 甘肃林业科技, 2005, 30(2): 32–35, 57.
[16] 杨文高, 字洪标, 陈科宇, 等. 青海森林生态系统中灌木层和土壤生态化学计量特征[J]. 植物生态学报, 2019, 43(4): 352–364.
[17] 周立华, 魏轩, 黄珊. 西北内陆河流域水资源管理利用的经验与启示[J]. 国土资源情报, 2017(12): 3–8.
[18] 王金叶, 常学向, 葛双兰, 等. 祁连山(北坡)水热状况与植被垂直分布[J]. 西北林学院学报, 2001, 16(S1): 1–3.
[19] Yang L S, Feng Q, Yin Z L, et al.. Identifying separate impacts of climate and land use/cover change on hydrological processes in upper stream of Heihe River, Northwest China[J]. Hydrological Processes, 2017, 31(20): 3592.
[20] 宋永昌. 植被生态学[M]. 上海: 华东师范大学出版社, 2001.
[21] Akburak S, Oral H V, Ozdemir E, et al. Temporal variations of biomass, carbon and nitrogen of roots under different tree species[J]. Scandinavian Journal of Forest Research, 2013, 28(1): 8–16.
[22] Yang Y S, Chen G S, Lin P, et al. Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in Subtropical China[J]. Annals of Forest Science, 2004, 61(7): 617–627.
[23] 郭大力. 植物根系: 结构、功能及在生态系统物质循环中的地位圈//国家自然科学基金委员会. 第三届现代生态学讲座暨国际学术研讨会论文集[M]. 北京: 高等教育出版社, 2005: 92–109.
[24] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000.
[25] 林业部科技司. 森林生态系统定位研究方法[M]. 北京: 中国科学技术出版社, 1994.
[26] Cao Y B, Wang B T, Wei T T, et al. Ecological stoichiometric characteristics and element reserves of three stands in a closed forest on the Chinese loess plateau[J]. Environmental Monitoring and Assessment, 2016, 188(2): 1–14.
[27] Jackson R B, Mooney H A, Schulze E D. A global budget for fine root biomass, surface area, and nutrient contents[J]. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(14): 7362–7366.
[28] 韩文轩, 吴漪, 汤璐瑛, 等. 北京及周边地区植物叶的碳氮磷元素计量特征[J]. 北京大学学报(自然科学版), 2009, 45(5): 855–860.
[29] 汪涛, 杨元合, 马文红. 中国土壤磷库的大小、分布及其影响因素[J]. 北京大学学报(自然科学版), 2008, 44(6): 945–952.
[30] Yu Q, Elser J J, He N P, et al. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland[J]. Oecologia, 2011, 166(1): 1–10.
[31] 刘旭艳, 胡宇坤. 大兴安岭典型森林沼泽植物叶片和细根碳氮磷化学计量特征[J]. 应用生态学报, 2020, 31(10): 3385–3394.
[32] Drenovsky R E, Richards J H. Critical N: P values: Predicting nutrient deficiencies in desert shrublands[J]. Plant and Soil, 2004, 259(1/2): 59–69.
[33] 赵维俊, 敬文茂, 赵永宏, 等. 祁连山大野口流域典型灌丛植物与土壤中氮磷的化学计量特征[J]. 土壤, 2017, 49(3): 572–579.
[34] 张光德, 赵传燕, 戎战磊, 等. 祁连山中部不同植被类型土壤生态化学计量特征研究[J]. 兰州大学学报(自然科学版), 2019, 55(4): 533–540.
[35] 吴鹏, 崔迎春, 赵文君, 等. 茂兰喀斯特区68种典型植物叶片化学计量特征[J]. 生态学报, 2020, 40(14): 5063– 5080.
[36] 李婷, 邓强, 袁志友, 等. 黄土高原纬度梯度上的植物与土壤碳、氮、磷化学计量学特征[J]. 环境科学, 2015, 36(8): 2988–2996.
[37] 李玉霖, 毛伟, 赵学勇, 等. 北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究[J]. 环境科学, 2010, 31(8): 1716–1725.
[38] 丁小慧, 罗淑政, 刘金巍, 等. 呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化[J]. 生态学报, 2012, 32(11): 3467–3476.
[39] 安申群, 贡璐, 朱美玲, 等. 塔里木盆地北缘典型荒漠植物根系化学计量特征及其与土壤理化因子的关系[J]. 生态学报, 2017, 37(16): 5444–5450.
Root Stoichiometric Characteristics of Typical Shrubs and Their Correlation with Soil Physicochemical Factors in Qilian Mountain
MA Jian1,2,3, FENG Qi2, ZHAO Weijun1,3, ZHAO Yonghong4, JING Wenmao1,3, WU Xiurong1,3, HE Yongyan1,3, CHEN Liying1,3
(1 Academy of Water Resources Conservation Forests in Qilian Mountains of Gansu Province, Zhangye, Gansu 734000, China; 2 Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China; 3 International Science and Technology Cooperation Base for Forest Ecological Monitoring and Assessment in Qilian Mountain, Gansu Province, Zhangye, Gansu 734000, China; 4 College of Hydraulic and Environmental Engineering, China Three Gorges University, Yichang, Hubei 443002, China)
In order to reveal the relationship between root nutrient content, stoichiometric characteristics and soil factors,,,andwhich lived in dayekou basint of Qilian Mountain were selected as object, the stoichiometric characteristics of coarse and fine roots and their relationship with soil physical and chemical factors were studied. The results showed that the average contents of C, N and P in coarse roots and fine roots were 569.55, 7.86, 1.02 g/kg and 541.38, 8.61, 1.10 g/kg, respectively. C/N, C/P and N/P of coarse roots and fine roots were 86.63, 729.78, 9.77 and 71.34, 527.61, 8.11, respectively. The contents of C and P were higher than the average levels of Chinese plant roots, while the N content of coarse roots was higher than the average level of Chinese plant coarse roots, but the N content of fine roots was lower than the average level of Chinese plant fine roots. Soil available phosphorus content and soil mass water content were the main soil factors affecting the stoichiometric characteristics of coarse roots, while soil available phosphorus content and soil pH value were the main soil factors affecting the ecostoichiometric characteristics of fine roots.
Typical shrubs; Coarse root; Fine root; Stoichiometries; Qilian Mountain
Q948
A
10.13758/j.cnki.tr.2022.02.016
马剑, 冯起, 赵维俊, 等. 祁连山典型灌丛植物根系化学计量特征及其与土壤理化因子的关系. 土壤, 2022, 54(2): 329–337.
甘肃省自然科学基金项目(20JR5RE641)、国家重点研发计划项目子课题(2019YFC0507404)、国家自然科学基金项目(41801015,4180070739)和国家社科基金项目(19BGL193)资助。
马剑(1986—),男,甘肃张掖人,博士,高级工程师,主要从事森林土壤生态研究。E-mail: 405153416@qq.com
