一种基于生理神经网络的智能步行机器人构建方法
2022-05-06周俊雅张强戴跃
周俊雅,张强,戴跃
华东师范大学通信与电子工程学院,上海200241
前言
行进运动是哺乳动物(如人类)最基本的运动之一(如走、跳、跑),它能够有节奏、重复地运动并产生位移。这种经典的运动是由分布在脊髓上的神经网络控制的,该网络被称为中枢模式发生器(Central Pattern Generator,CPG)。传统的智能行走机器人运动是一种关节间的机械运动,大大简化了神经网络的调节和控制。具体来说,这类机器人通过一系列机械装置和相关的控制算法来执行任务或与人互动[1-3]。也就是说,传统智能行走机器人的运动控制并没有引入CPG 网络的引导和控制机制。然而,从生理角度出发,人类的肢体之所以可以精确地表现出各种复杂而精细的关节运动是由于CPG 网络在其中起着重要的作用[4-5]。在智能行走机器人领域,CPG 网络可以通过控制肌肉(人造肌肉材料)的收缩来驱动骨骼的运动。基于这样的理论推断,CPG 网络模型可以构建具有生理机制的智能行走机器人。
控制肢体运动的原始神经网络模型是“半中心”模型[6-8],它由一对相互抑制的屈伸神经元组成,控制骨骼肌的收缩。目前,CPG 网络的应用是基于“半中心”模型发展起来的。神经系统中存在许多这样的CPG 网络模型,如呼吸神经系统模型[9]、肌肉收缩神经系统模型等[10]。尽管CPG 神经网络得到了发展,但很少有研究将CPG 应用于机器人。本文主要研究控制骨骼肌收缩的CPG 模型的实现。CPG 模型的神经信号转导可以用Hodgkin-Huxley 动力学方程[11]来描述。此外,神经元之间的交流是通过兴奋性或抑制性突触进行的,兴奋性和抑制性信息的传递保证了神经元之间的正常交流。本文提出基于CPG 网络的智能行走机器人设计方法,将为下一代智能行走机器人的发展奠定一定的理论基础。
1 单神经元模型
神经元信号通过离子通道的开启和关闭来传递,从而产生动作电位。神经元电信号的传播模型等效于基尔霍夫电路,离子通道相当于一系列的非线性电导。这些导体的数学方程服从霍奇金-赫胥黎(Hodgkin-Huxley, H-H)模型。换句话说,神经元电位的变化可以通过一个由并联导体组成的电路来模拟。在这个模型中,本文建立了两种类型的神经元:一种是运动神经元,另一种是中间神经元。控制骨骼肌收缩的神经元被称为运动神经元。根据电生理研究结果,本文构建了运动神经元模型(图1)。运动神经元模型的结构为胞体和树突。瞬时性钠通道(NaT)、持续性钠通道(NaP)、延迟整流钾通道[K(DR)]、钙依赖的钾离子通道[K(AHP)]、L型钙通道(CaL)和超极化内向电流(H)分布在运动神经元上(图1)。瞬时性钠通道和延迟整流钾通道是参与动作电位形成的两个重要渠道。持续性钠通道是神经网络节律性放电的关键通道。
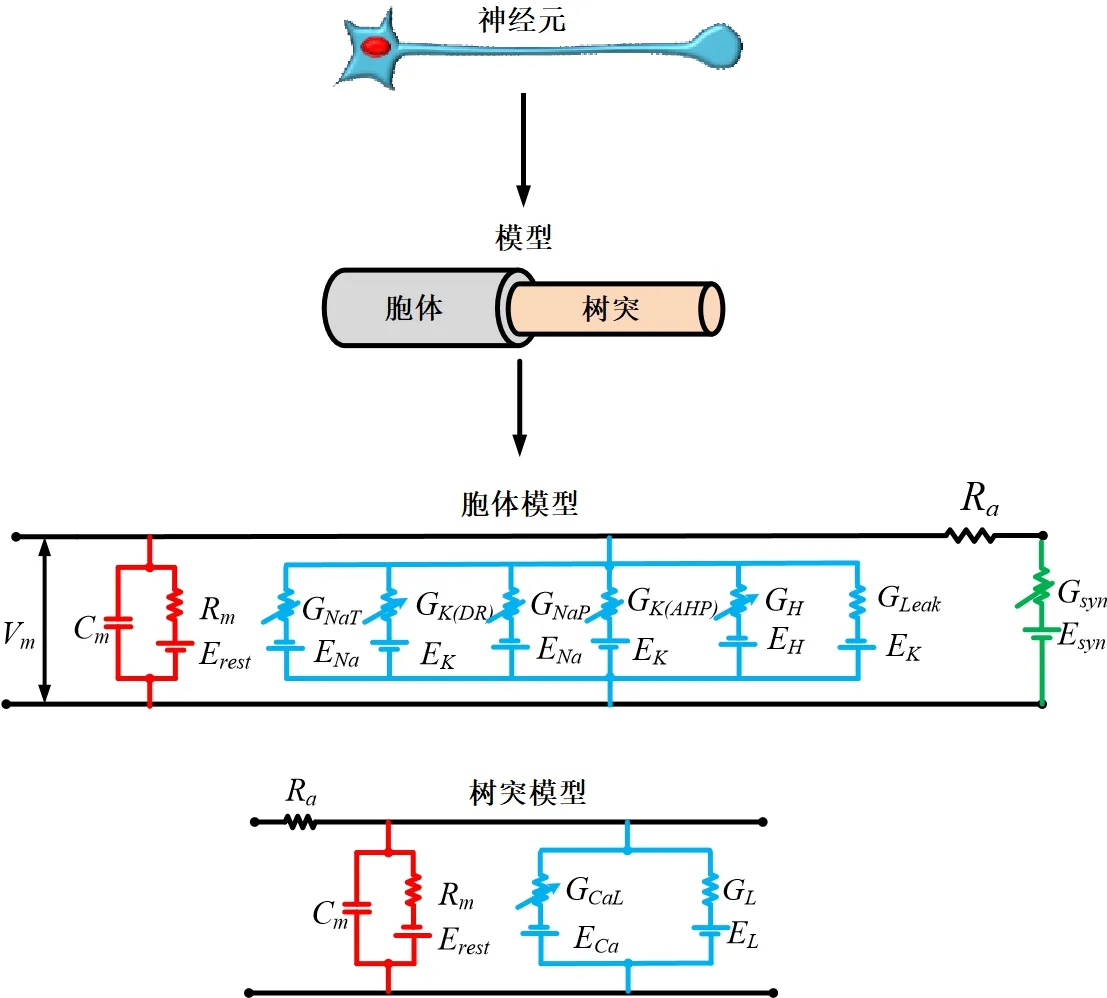
图1 运动神经元模型(包括胞体和树突,CPG网络中的其他神经元模型进一步简化,只包括胞体)Figure 1 Motor neuron models(including soma and dendrite,and the other neuron models in the CPG network are further simplified,including only soma)
由图1可知,神经元模型可以等效为一个由膜电容(Cm)、膜电阻(Rm)和一系列平行连接的离子通道组成的电路。根据基尔霍夫电流定律,可以得到膜电流方程:
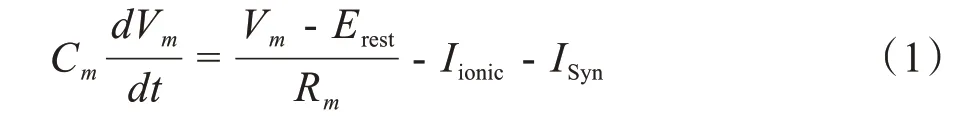
其中,Vm和Erest分别代表膜电位和静息膜电位。不同哺乳动物的静息膜电位略有不同,从-90 mV到70 mV不等。Iionic和ISyn分别表示离子电流和突触电流。电流方程的具体表达式如下:
其中,gNaT是瞬时性钠电流的最大电导,gNaP是持续性钠电流的最大电导,同理,gK(DR)、gK(AHP)、gCaL、gH、gLeak、gSyn分别是延迟整流钾电流、钙依赖的钾电流、L型钙电流、超极化内向电流、漏电流及突触电流的最大电导,ENa、EK、ECa、EH、EL、ESyn分别是Na+、K+、Ca2+和突触的平衡电位。wSyn是突触的重量。状态变量m、h、n、q是随时间和电压变化的函数,表示离子通道的开启和关闭。这些状态变量是由霍奇金-赫胥黎方程定义的。
除了运动神经元,CPG网络还有其他神经元(如抑制性中间神经元)。为了简化模型,本文将其他神经元模型只设置胞体,并且胞体上离子通道的分布与运动神经元模型相同。
2 CPG
CPG 中枢模式发生器是产生哺乳动物节律性运动行为的生理神经环路,它由一系列神经振荡器组成,是神经振荡器与多重反射回路系统集成在一起组成的一个复杂的分布式神经网络。图2 是控制智能行走机器人两条腿交替运动的CPG网络模型。CPG网络模型的神经元基于单神经元模型的理论进行设计。CPG 网络模型由左、右四肢交替控制层、节律及模式生成层和神经-肌肉-骨骼层组成。CPG 网络最大的特点是控制两条腿前后运动的兴奋性信号交替出现,任意一条腿的屈伸肌也交替刺激,从而保证了四肢的正常运动。CPG 网络的功能类似于推挽电路。具体地说,两个相位相反信号在序号①、②处生成,此时CPG 神经网络收到一个兴奋性信号输入(为方便描述,正弦波用来表示动作电位的传播过程,上升沿代表了兴奋性动作电位的部分,它可以激活下一个层次的神经元;下降沿代表动作电位的抑制部分,阻止下一级神经元的激活)。当信号被传递到节律和模式生成层时,它也分别在序号⑦和⑧或⑨和⑩产生两个相位相反的信号。左腿伸肌兴奋(⑪)和屈肌抑制(⑫),使得左腿处于站立状态;右腿伸肌抑制(⑬)和屈肌兴奋(⑭),则使右腿处于蜷缩状态。CPG 网络是按照上述规律循环运行的,以上就是CPG模型的工作原理。此外,CPG 模型中的每个节点不是单个神经元,而是多个神经元组成的神经元池,如图2所示。CPG神经网络之所以可以产生交替的兴奋性信号控制双腿是因为交互抑制性中间神经元的存在(例如,左和右肢交叉层,节律和模式生成层)以及离子通道的分布,持续性钠离子通道在产生节律信号的过程中发挥着重要作用[12-13]。
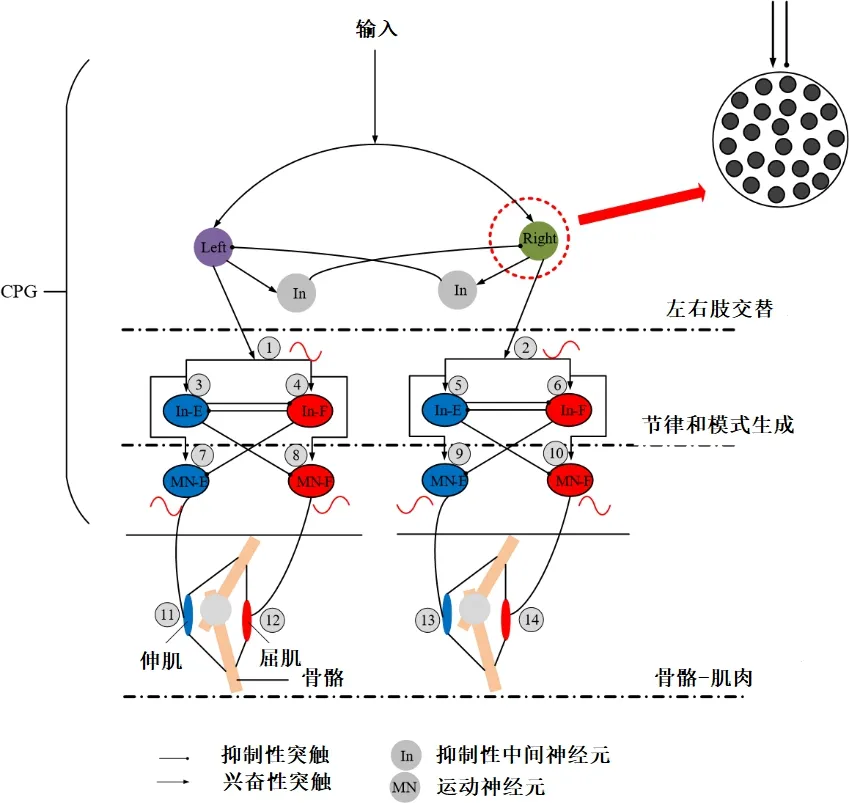
图2 CPG神经网络控制骨骼肌收缩模型Figure 2 Skeletal muscle contraction model controlled by CPG network
3 电信号生成机制
神经元信息是通过动作电位传递的。当信号到达骨骼肌时,运动神经元池的信号被叠加(时间和空间的总和)形成肌电信号。肌电最基本的单位是运动单位动作电位(Motor Unit Action Potential,MUAP)。肌电信号的产生如图3 所示。首先,获得运动神经元池(SAP)的所有时间依赖性动作电位。然后捕捉瞬时动作电位(Fi)。最后,将采样数据与小波(FM)进行卷积得到肌电信号(SEMG)。综上所述,上述过程可以用下式表示:
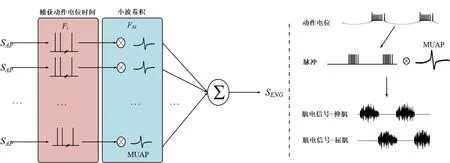
图3 肌电信号的产生方法(左图)和肌电信号的产生实例(右图)Figure 3 Method of EMG signal generation(left)and an example of EMG signal generation(right)
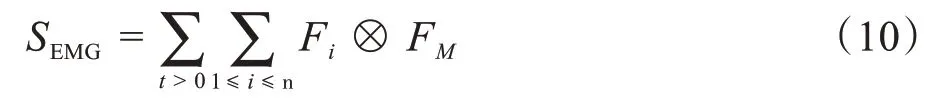
其中,n代表神经元池中神经元的数量(本文中神经元池中均有100个神经元),t是采样时间。
4 实验结论
CPG 模型生成节律信号的一个重要原因是持续性钠离子通道门控的交替打开和关闭[14]。此外,网络中交互抑制电路的存在生成了屈肌和伸肌信号的交替。换句话说,控制屈肌的运动神经元兴奋,抑制了控制伸肌的运动神经元,反之亦然。同时,当持续性钠通道失活数量达到最大时,神经元的兴奋性受到抑制。相反,当失活数量达到最小时,神经元的兴奋性增加,如图4所示。
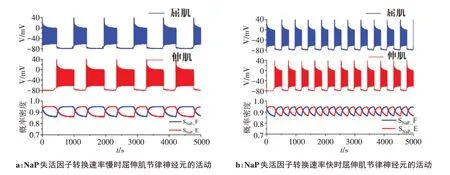
图4 持续性钠通道作用下CPG网络产生的节律信号Figure 4 Rhythm signals generated by CPG network under the action of persistent sodium channels
步态的频率可以在行进运动过程中改变,这是因为CPG 网络可以合理地改变输出的节律频率。通过增加持续性钠通道的数目(增加持续性钠通道的电导)增加神经元的节律频率。节律性频率的变化是由于持续性钠通道失活的速率变化。持续性钠通道失活(α)越快,节律性信号变化越快(图4b)。
来自运动神经元池的信号可以控制骨骼肌的收缩,并导致骨骼运动。当左腿屈肌兴奋时,左腿抬起。同时,当右腿伸肌兴奋时右腿保持站姿。同理,左肢伸肌的兴奋使左腿站立,与此同时,右腿弯曲。这个过程就使得智能行走机器人的双腿可以交替行走(图5)。
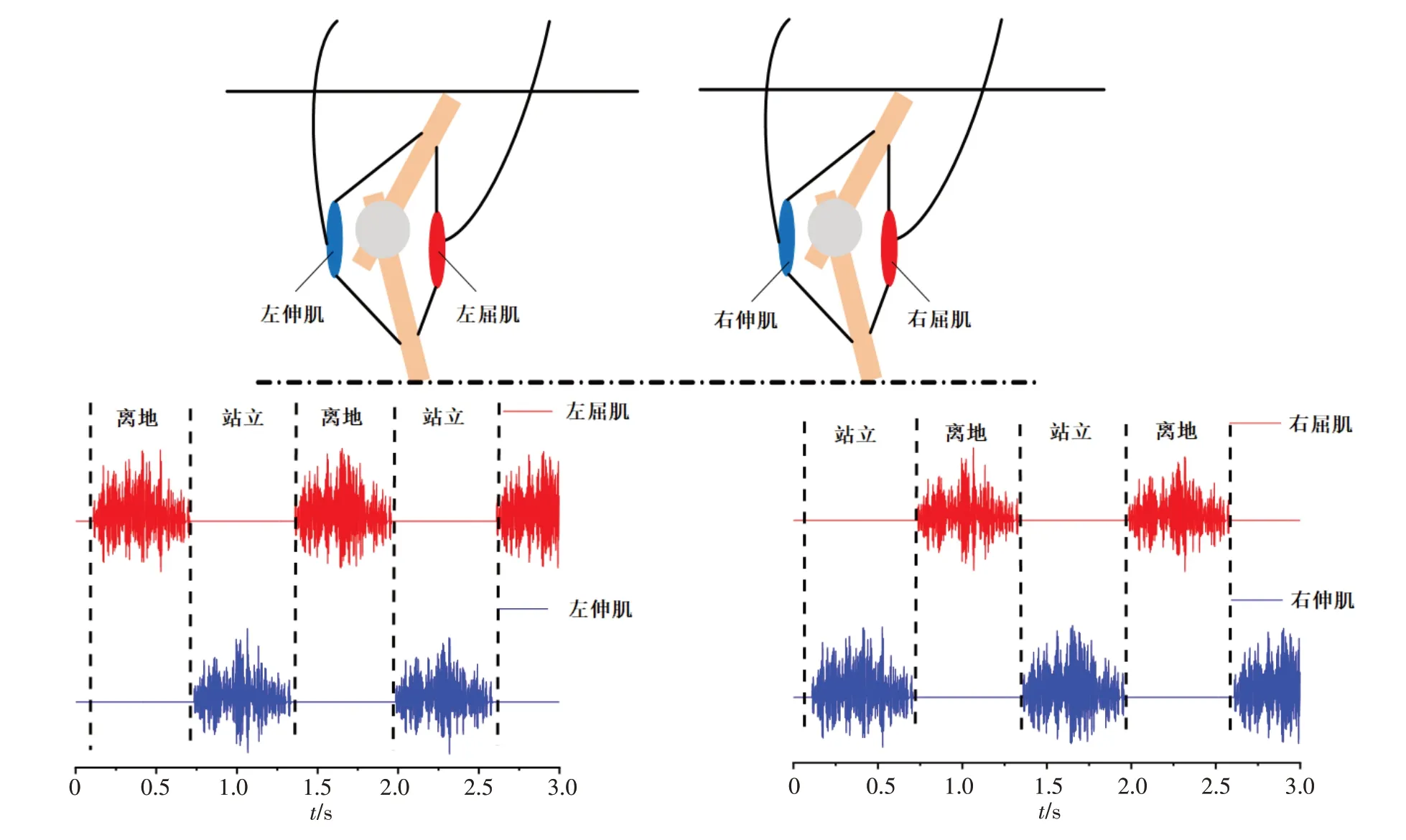
图5 骨骼肌产生肌电图信号,使左右四肢交替运动Figure 5 Skeletal muscle produces EMG signals to make the left and right limbs move alternately
5 讨论
本文提出了一种不同于传统机器人关节运动的智能行走机器人构造方法,这种新型机器人基于生理神经网络的原理控制四肢的交替[15-26]。该设计方法最大的特点是采用了基于真实神经元电传播的神经网络模型(CPG 模型),这对于我们更好地理解生理神经网络的结构和功能具有重要意义。当然,由于生理神经网络的复杂性,本文设计的步行机器人有以下局限性:神经元一般由胞体、树突和轴突组成。神经元的兴奋性与其结构密切相关。为了提高运算速度,CPG 模型将神经元简化为胞体和树突;目前,CPG 模型只能实现固定模式行走,人类的腿在现实生活中可以实现多种步态模式。接下来,我们将进一步研究肌电信号和步态模式与机器人做出更复杂动作之间的关系。