菜薹花色遗传规律研究及花瓣主要营养成分分析
2022-04-26吕思锐张德双李佩荣辛晓云张凤兰于拴仓余阳俊张耀伟赵岫云汪维红苏同兵卢桂香
吕思锐 张德双 李佩荣 辛晓云 张凤兰 于拴仓 余阳俊 张耀伟 赵岫云 汪维红 苏同兵 卢桂香
(1 北京市农林科学院蔬菜研究所,北京 100097;2 东北农业大学园艺园林学院,黑龙江哈尔滨 150030)
菜薹〔L. ssp.(L.)var.Tsen et Lee〕起源于中国,又名绿菜薹、广东菜、菜尖等,是十字花科芸薹属芸薹种白菜亚种中的一个变种,以花茎和薹叶作为食用部位,薹茎柔嫩,风味独特,营养丰富,享有“蔬菜之冠”的美誉,广州地区常年栽培面积16 800 hm左右(张华和刘自珠,2010;陈汉才 等,2021)。
花色性状表型直观,遗传稳定,受环境影响较小,是重要的标记性状之一,可以在提高种子纯度、去除杂株、鉴定天然异交率等方面起到重要作用(戚存扣和傅寿仲,1992)。花色还可作为吸引昆虫或鸟类的视觉信号,黄花和橙花更容易吸引昆虫辅助授粉(Vainstein et al.,2001)。一般情况下,花瓣颜色为黄色和橙色可反射含热量多的红、橙、黄光,从而可以维持花朵的能量平衡,在一定程度上保护花不被灼伤,花器官免受伤害(赵昶灵 等,2005)。花瓣所含色素种类及其含量的不同导致花瓣呈现多种颜色。类胡萝卜素可以使花瓣呈现黄色到红色等多种颜色。在花瓣呈色过程中,大多数类黄酮物质也是不可或缺的(Tanaka et al.,1998)。菜薹的花色主要有4 种:黄色、白色、橙色和乳白色,其中黄色为主要花色。关于菜薹的花色遗传,乳白色花和橙花的遗传规律、显隐性关系未见报道。
本试验以乳白色花和橙花菜薹为材料,研究菜薹乳白色花与橙花的显隐性关系,揭示菜薹花色的遗传规律;同时通过对F群体出现的黄色、乳白色和橙色3 种花色花瓣类胡萝卜素和黄酮组分及含量的测定,为创制高类胡萝卜素含量和高黄酮含量的菜薹新种质提供理论依据,以期指导高营养成分菜薹新品种的选育工作。
1 材料与方法
1.1 试验材料
供试材料为自交6 代以上稳定的菜薹自交系14A-P2-21 和广海超级80 天,均由北京市农林科学院蔬菜研究所大白菜课题组提供。14A-P2-21 花橙色,颜色鲜艳且较深,植株生长势中等,叶片翠绿色、近椭圆形,菜薹翠绿色,分枝较多;广海超级80 天花乳白色,植株生长势强,叶片浅绿色、卵圆形,菜薹绿色,分枝多。
1.2 试验方法
1.2.1 菜薹花色遗传群体构建 2019 年4 月,播种亲本14A-P2-21(P)和广海超级80 天(P);5月,配制广海超级80 天× 14A-P2-21 的F。2020年4 月,播种14A-P2-21、广海超级80 天和F;5 月,F自交获得F,F分别与P、P回交获得BC、BC′群 体。2020 年9 月27 日 将P、P、F、F、BC和BC′6 个世代的材料播种在50 孔穴盘中,10 月20 日定植到日光温室中。2021 年1月4 日开始调查6 个世代全部单株花色并拍照。为了进一步扩大群体数量,验证花色分离的准确性,2021 年5 月20 日在日光温室中继续播种6 个世代的材料,6 月23 日开始调查6 个世代全部单株花色并拍照,其中F群体开花持续时间长,花色调查工作一直持续到7 月20 日。花色调查方法依据GB/T 19557.5—2017《植物品种特异性、一致性和稳定性测试指南 大白菜》。全部试验材料的播种育苗、定植等田间管理都安排在北京市农林科学院蔬菜研究所农场进行。
1.2.2 花粉活力测定和花粉管萌发试验 2021 年7月27 日开始,在晴天的上午10:00,分别取P、P、F、F花粉进行花粉活力测定和花粉管萌发试验。花粉活力测定采用FDA 染色法(胡适宜,1993):取适量FDA 溶液于载玻片上,分别将黄花、乳白色花和橙花盛花期花朵置于FDA 溶液,用镊子抖落花粉;随机观察3 个视野,每个视野观察20 个花粉粒,重复3 次。花粉管萌发试验采用离体萌发法(陈奇 等,2017;Jiang et al.,2019),培养基采用固体花粉萌发培养基GM〔1 mmol·LCaCl,1 mmol·LCa(NO),1 mmol·LMgSO,0.01%(/)HBO,18%(/)蔗糖,0.8%(/)琼脂,pH 值7.2〕;取干净载玻片置于垫有滤纸的培养皿内,将适量GM 固体培养基放于载玻片上,分别取黄花、乳白色花和橙花盛花期的花粉均匀涂抹于培养基,盖上培养皿盖子,35 ℃培养3 h;随机观察3 个视野,每个视野观察30 个花粉粒,以花粉管长度大于花粉粒直径作为花粉萌发标准,重复3 次。使用Zeiss Axio Imager M2 光学显微镜进行花粉FDA 染色荧光观察和花粉管萌发观察。
1.2.3 类胡萝卜素和黄酮组分及含量测定 2021 年8 月19 日,取F群体的黄花、乳白色花和橙花花瓣各1 g,3 次生物学重复,送至武汉迈特维尔生物科技有限公司,利用液相色谱串联质谱法(LCMS/MS)测定类胡萝卜素和黄酮组分及含量,并绘制维恩图。
2 结果与分析
2.1 菜薹6 个世代田间性状
田间观察结果表明(图1~3),橙花亲本14A-P2-21(P)生长势较弱,花期晚,开花迟,花瓣橙色、颜色较深,且花药颜色也较深;乳白色花亲本广海超级80 天(P)生长势较强,花期早,花瓣黄色、颜色鲜艳;杂交F生长势明显强于双亲,花瓣黄色、颜色鲜艳;F群体中黄花出现较早,其次为乳白色花,橙花较晚,且植株较弱小。双亲和F花色的比较结果见图4。

图1 6 个世代苗期

图2 F2 成株期

图3 F2 盛花期

图4 P1、P2、F1 花色比较
2.2 菜薹花色遗传规律
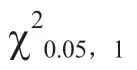
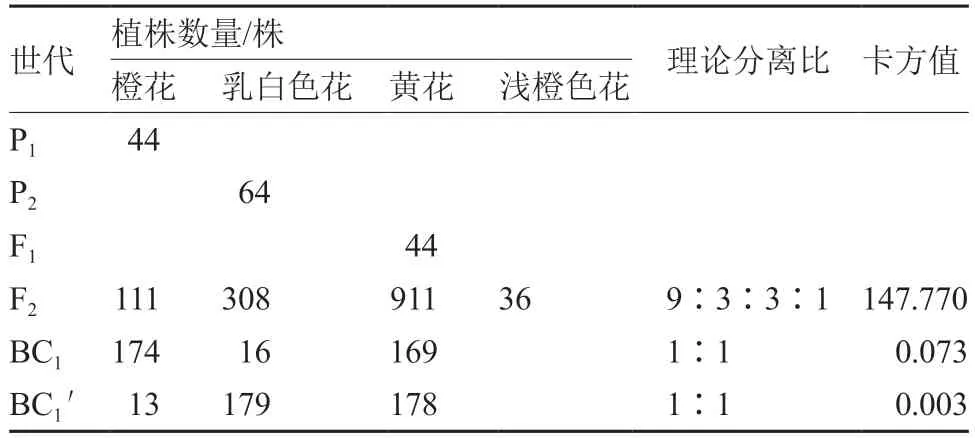
表1 菜薹6 个世代的花色遗传分析

图5 F2 群体出现的4 种花色
2.3 菜薹花粉活力和花粉管萌发情况
为了深入解析菜薹F群体的4 种花色分离比不符合9∶3∶3∶1 遗传规律的原因,即橙花植株数量少、出现偏分离的原因,进一步进行花粉管萌发试验,深入比较黄花、乳白色花和橙花的雄配子活力差异。花粉经FDA 染色后,有活力的花粉粒整体表现绿色荧光,无活力的花粉粒只在边缘出现绿色荧光轮廓。光学显微镜观察可知(图6),橙花花粉粒活力明显低于黄花和乳白色花。计数和统计分析结果表明(表2),黄花花粉活力为21.67%;乳白色花次之,为20.56%,两者差异不显著;而橙花花粉活力仅为5.00%,与黄花和乳白色花的花粉活力差异均达显著水平。由图7 可见,黄花花粉萌发情况好,可以正常萌发,橙花花粉萌发不均匀且萌发率低。经计数和统计分析(表2),3 种花色的花粉管萌发率之间差异均达显著水平,黄花最高,为42.22%;乳白色花次之,为25.19%;橙花最低,为20.37%。

图6 不同花色菜薹的花粉活力比较

表2 不同花色菜薹的花粉活力和花粉管萌发率

图7 不同花色菜薹的花粉管萌发情况
2.4 菜薹不同花色花瓣类胡萝卜素和黄酮的组分及含量
2.4.1 类胡萝卜素组分及含量 在黄花、乳白色花和橙花花瓣中共检测到41 种类胡萝卜素组分,其中八氢番茄红素、γ-胡萝卜素、紫黄质二月桂酸酯、叶黄素癸酸酯、5,6 环氧叶黄素二月桂酸酯和叶黄素月桂酸酯为6 种差异较大的组分。从表3 可以看出,橙花花瓣中总类胡萝卜素含量为2 663.98 μg·g,极显著高于黄花和乳白色花;此外,橙花花瓣中八氢番茄红素和γ-胡萝卜素含量极显著高于黄花和乳白色花,黄花花瓣中紫黄质二月桂酸酯、叶黄素癸酸酯、5,6 环氧叶黄素二月桂酸酯和叶黄素月桂酸酯含量极显著高于乳白色花和橙花。
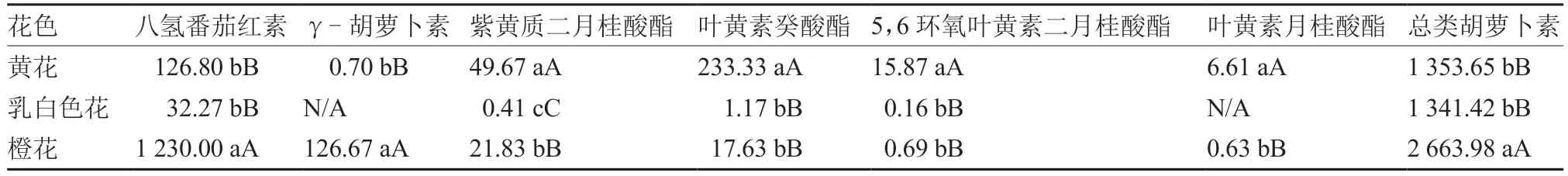
表3 不同花色菜薹花瓣的类胡萝卜素组分及其含量 μg·g-1
2.4.2 黄酮组分及含量 对黄花、乳白色花和橙花花瓣的黄酮组分及其含量进行测定,共检测到74种成分,其中含量较高的6 种组分及含量见表4。从表4 可以看出,橙花花瓣中总黄酮含量为593.68 nmol·g,极显著高于黄花和乳白色花;此外,橙花花瓣中紫云英苷含量显著高于黄花,黄花花瓣中芒柄花苷含量显著高于乳白色花和橙花,3 种花色花瓣中异鼠李素-3-O-葡萄糖苷、金丝桃苷、槲皮素-3-槐糖苷、异鼠李素含量差异均不显著。

表4 不同花色菜薹花瓣的黄酮组分及其含量 nmol·g-1
2.4.3 黄酮组分差异物质维恩图 通过维恩图可以直观的得到3 种花色之间共有和特有的差异物质。如图8 所示,在检测到的74 种黄酮物质中,黄花和乳白色花的差异物质有3 种,分别为儿茶素没食子酸酯、芦丁、高车前素;橙花和黄花的差异物质有6 种,分别为圣草酚、儿茶素没食子酸酯、芹菜素7-葡萄糖苷、柚皮素7-O-葡萄糖苷、槲皮素、3-羟基根皮苷;橙花和乳白色花的差异物质有2种,分别为3-羟基根皮苷、高车前素。
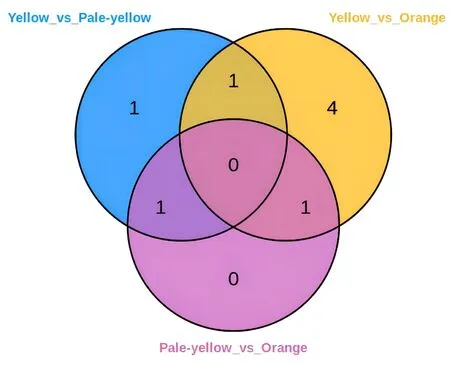
图8 不同花色菜薹花瓣的黄酮组分差异物质维恩图
3 结论与讨论
3.1 菜薹乳白色花和橙花的遗传规律
菜薹的花色以黄色为主,已有研究表明白花对黄花为隐性,由1 对等位基因控制(张华和刘自珠,1999);菜薹橘红色花、乳白色花与白花的遗传规律相似,橘红色花和乳白色花对黄花均为隐性,均由1 对等位基因控制(张德双 等,2013,2019)。在甘蓝型油菜CMS 不育系橘红色花恢复系中,橘红色花基因受2 对隐性核基因控制,并可随配子有性传递(李莓 等,1999)。在大白菜中,白花、橙花和淡黄色花对黄花均为隐性,且均由单基因控制(Séguin-Swartz,1988;Rahman,2001;张德双 等,2003;Feng et al.,2012;Lee et al.,2014);也有报道表示白花由2 对隐性基因控制(Alam &Aziz,1954)。大白菜橙花与白花杂交F群体中出现黄花、橙花和白花3 种花色,分离比例不符合单基因遗传规律,相关研究表明白花和橙花性状为2 对隐性上位基因调控的互作遗传(金鑫,2020)。在芥菜型油菜中,研究者多认为花色变异出现浅黄花及(乳)白花是由2 对基因控制,且2 对隐性基因表现出相互作用(Alam &Aziz,1954;Bhuiyan,1986;Singh &Chauhan,2011)。本试验中,菜薹橙花与乳白色花的F群体中出现了4 种花色,其中浅橙色花为新的花色,该花色比乳白色深但又浅于橙色,表明菜薹乳白色花和橙花之间不存在显隐性关系,其遗传模式为2 对隐性、非等位基因控制,不同于大白菜的2 对隐性上位基因调控的互作遗传。
3.2 菜薹F2 群体中橙花分离比例低于理论值的原因
在甘蓝型油菜中,由于纯合隐性的橘红色花雄配子生活力较低,在各花色分离世代中橘红色花分离株低于理论预测值(李莓 等,1999)。本试验中,菜薹黄花、乳白色花、橙花的花粉活力测定结果和花粉管萌发试验结果均证实:在相同条件下橙花的配子体竞争力弱,授粉概率低。同时,田间调查也发现,F中橙花单株存在开花晚且植株弱小的情况,在相同的栽培环境条件下抗逆能力弱,存活率低于黄花和乳白色花植株。这些可能导致橙花单株数量少,低于理论值,出现偏分离的现象。在BC群体中,即F为母本和14A-P2-21 为父本的回交后代,黄花和橙花分离比例为1∶1,符合1 对基因控制模型,其原因是在F× 14A-P2-21 中橙花为纯合隐性基因型,与其他花色不存在竞争关系。
3.3 菜薹花瓣营养成分差异及意义
花色的差异是由于花色素种类和含量不同所导致(Tanaka et al.,1998)。根据本试验菜薹3 种花色花瓣的类胡萝卜素和黄酮组分及含量测定结果,橙花花瓣中总类胡萝卜素和总黄酮含量极显著高于黄花和乳白色花。同时,这两种营养物质在菜薹的生长、发育、抗逆境胁迫等多种生物进程中发挥着重要的作用,对人类的健康也具有保健作用,不仅可促进血液循环,预防心血管疾病,还具有抗氧化、抗肿瘤和降血糖、降血脂等多种药理作用(Mahadevan &Park,2008;Ma &Zhao,2009;Heiss et al.,2010;Hollman et al.,2010)。因此,花瓣总类胡萝卜素和总黄酮含量较高的橙花菜薹是菜薹品种选育的新方向。下一步,笔者还将测定浅橙色花菜薹的营养成分,为开展不同花色菜薹品种的选育工作奠定基础。
3.4 菜薹BC1 群体出现乳白色花以及BC1′群体出现橙花的原因
本试验中,在回交BC群体(359 株)中出现了16 株乳白色花植株,BC′群体(370 株)中出现了13 株橙花植株。可能的原因:一方面,在获得F× 14A-P2-21 回交一代BC,F×广海超级80天回交一代BC′时,F作为母本去雄不彻底,导致F的极少量花粉自交,在BC和BC′群体中分别出现乳白色花和橙花植株。另一方面,在种子收获、清选、人工播种过程中出现机械混杂。类似的结果也有报道(张德双 等,2013)。因此,为避免此类问题应严格去除F雄蕊,规范操作,避免机械混杂等情况发生。
