南高丛蓝莓品种蓝美1 号的光合特性研究*
2022-04-20韦继光曾其龙姜燕琴蒋佳峰田亮亮刘梦溪
韦继光,曾其龙,姜燕琴,蒋佳峰,田亮亮,刘梦溪,於 虹
(江苏省中国科学院植物研究所,南京 210014)
鲜食加工兼用型南高丛蓝莓新品种蓝美1 号(Vaccinium croymbosumcv.‘Lanmei 1’)是由浙江蓝美农业有限公司选育的国家级林木良种(编号:国R-ETS-VC-006-2018)[1]。该品种为种间杂交后代优中选优所得,既继承了野生蓝莓抗旱、耐热和耐瘠薄的优点,又富含野生蓝莓高级保健功能。该品种树势强,树姿半开张,株高1.2~2.0 m;果实圆形,平均单果重1.38 g,硬度中等,果实可溶性固形物含量 12.0%、总糖含量 10.9%、总酸含量0.589%,糖酸比适中;果实早熟(成熟期在5 月下旬至6 月上旬),有利于抢商机、避梅雨,而开花晚(与奥尼尔相比),有利于避晚霜;自交结实性优,丰产性好、产量高,进入稳定期后平均株产4~6 kg,667 m2产量在1 000 kg 以上;适应性强,易种植,适宜在我国多种不同生态区域(华东、华中、华南、西南等地区)产业化栽培[2-4]。因具有适应性强、易种植、产量高、营养优等优点,蓝美1 号在我国南方蓝莓产区推广种植面积迅速增加,目前全国种植面积已超过1 333.3 hm2。但蓝美1 号生态适应性强、产量高的光合生理生态机制尚不明确。为此,本研究以常规南高丛蓝莓品种奥尼尔为对照,对蓝美1 号光合特性及相关的生理生态因子进行测定分析,试图从叶片光合特征和水分利用效率角度揭示蓝美1 号广适性及丰产性的生理生态机制,为其引种栽培及推广种植提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于江苏省中国科学院植物研究所(北纬32°3′29′′,东经118°49′46′′),当地属亚热带季风气候区,雨量充沛,平均年降水量1 021 mm,四季分明,年平均气温15.7 ℃,极端最高气温39.7 ℃,极端最低气温-13.1 ℃,平均年日照时数2 213 h,无霜期237 d。试验地土壤为黄棕壤,厚度10~90 cm,0~15 cm 土层土壤pH 值为5.3,有机质含量为2.38%。
1.2 试验材料
供试材料为4 年生南高丛蓝莓品种蓝美1 号,对照品种为奥尼尔,行株距2.0 m×1.5 m,东西行向,施肥、浇水、修剪、病虫害管理措施相同。
1.3 测定指标及方法
于2020 年8 月下旬进行各项指标测定。每个品种选择生长势较为一致的植株3 株,对其株高、冠幅和基生枝数量进行测量;每个单株选取树冠中上部外围枝条上生长状况一致的向阳健康功能叶片,采用便携式光合作用测量系统LI-6800(美国LI-COR 公司)进行光合指标测定。
(1)光合日变化测定。于晴天9:00—17:00每隔1 h 测定1 次标记叶片的净光合速率(A)、胞间CO2浓度(Ci)、蒸腾速率(E)、气孔导度(Gs)、光合有效辐射(PAR)、叶片温度(Tl)等参数,并计算水分利用效率(WUE)=A/E。
(2)光合-光响应曲线测定。于晴天8:30—11:30 测定光响应曲线。叶室相对湿度控制在55%~65%,叶温控制在30 ℃,叶室CO2浓度控制在400 μmol/mol,PAR 梯度分别设定为1 800、1 500、1 200、900、600、300、200、150、100、70、30、0 μmol·m-2·s-1。每个梯度下平衡120 s,自动记录每个光照强度下叶片的净光合速率等参数。利用直角双曲线修正模型拟合所测定的光响应曲线[5-6],得出最大净光合速率(Amax)、光饱和点(LSP)、表观量子效率(AQY)、光补偿点(LCP)和暗呼吸速率(Rd)等参数。
(3)光合-CO2响应曲线测定。于晴天8:30—11:30 进行测定,采用CO2控制系统提供气源,叶室相对湿度控制在 55%~65%,叶温控制在30 ℃,叶室PAR 设为1 200 μmol·m-2·s-1,叶室CO2浓度梯度分别设为1 800、1 500、1 200、1 000、800、600、400、200、100、50 μmol/mol,每个梯度下平衡180 s,自动记录每个CO2浓度下叶片的净光合速率等参数。采用直角双曲线修正模型拟合所测定的CO2响应曲线[5-6],得出最大净光合速率(Amax)、初始羧化效率(CE)和光呼吸速率(Rp)。利用Photosyn Assistant 拟合CO2响应曲线,得到Rubisco酶最大羧化速率(Vcmax)、电子传递链潜在最大电子传递速率(Jmax)等特征参数。
(4)光合-温度响应曲线测定。于晴天8:30—11:30 进行测定,叶室相对湿度控制在55%~65%,叶室CO2浓度控制在400 μmol/mol,叶室PAR设为1 200 μmol·m-2·s-1,叶片温度(Tl)梯度分别设为30、35、40 ℃,每个梯度下平衡300 s,自动记录每个叶温梯度下叶片的净光合速率等参数。
2 结果与分析
2.1 植株生长状况
由表1 可知,蓝美1 号的植株冠幅和基生枝数量均显著高于奥尼尔,2 个品种的株高没有显著差异。
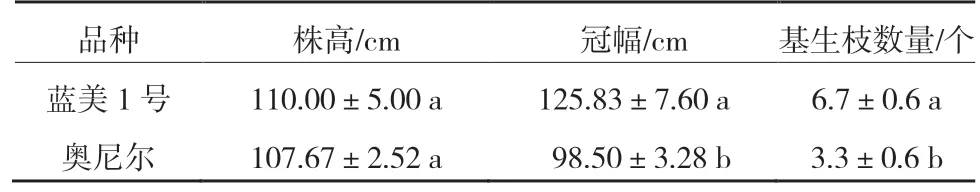
表1 2 个南高丛蓝莓品种的植株生长状况
2.2 日平均光合作用特征参数
从表2 可以看出,蓝美1 号的净光合速率(A)日均值和蒸腾速率(E)日均值均显著高于奥尼尔,而水分利用效率(WUE)日均值显著低于奥尼尔。

表2 2 个南高丛蓝莓品种的净光合速率(A)、蒸腾速率(E)、水分利用效率(WUE)日均值比较
2.3 光合有效辐射(PAR)和气温日变化动态
如图1 所示,从9:00 至17:00,PAR 呈先增加后降低的变化趋势,最高PAR 出现在13:00;气温呈波动上升而后降低的趋势,最高气温(36.9 ℃)出现在16:00,较PAR 峰值滞后3 h。

图1 试验地光合有效辐射(PAR)和气温日变化动态
如图2 所示,2 个南高丛蓝莓品种净光合速率(A)日变化曲线为双峰型,表现出明显光合“午休”现象。2 个品种A 第2 个峰值出现时间、“午休”发生的时间及“午休”光合速率的下降幅度有所差异。蓝美1 号峰值出现在10:00 和15:00,最低值出现在13:00,较峰值下降幅度为34.6%;奥尼尔峰值出现在10:00 和16:00,最低值出现在15:00,光合速率下降延续时间较长,下降幅度大,较峰值下降幅度达62.3%,且回升较慢。2 个南高丛蓝莓品种气孔导度(Gs)日变化趋势与A 日变化趋势相似,均为双峰曲线,且峰值与谷值出现时间也与A 相同。蓝美1 号蒸腾速率(E)日变化呈双峰曲线,峰值出现在10:00 和13:00,而奥尼尔呈三峰曲线变化,峰值出现在10:00、13:00和16:00。蓝美1 号胞间CO2浓度(Ci)日变化趋势与A、Gs 的日变化趋势基本相似;奥尼尔Ci 日变化呈波动上升趋势,因气孔关闭及光合消耗CO2,Ci 在12:00 出现最低值,因叶肉细胞羧化能力降低等非气孔限制因素导致光合消耗CO2减弱,Ci 在13:00 出现较大幅度升高。
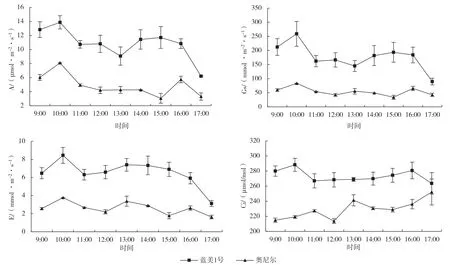
图2 2 个南高丛蓝莓品种净光合速率(A)、气孔导度(Gs)、蒸腾速率(E)、胞间CO2浓度(Ci)日变化动态
2.4 光合作用的光响应特征
由图3 可知,2 个南高丛蓝莓品种的光合-光响应曲线实测值和拟合值相似度较高。在光合有效辐射(PAR)≤200 μmol·m-2·s-1时,奥尼尔叶片净光合速率(A)随PAR 增加而迅速上升,之后A 变化曲线趋于平缓。蓝美1 号的光响应曲线则表现为在PAR≤300 μmol·m-2·s-1区间,A 随PAR 增加 近似呈线性增加,直至PAR≥900 μmol·m-2·s-1后A 增幅才逐渐趋于平缓。在弱光下(PAR≤100 μmol·m-2·s-1),2 个南高丛蓝莓品种A 差异相对较小,在150~1 800 μmol·m-2·s-1范围内,蓝美1号A 明显高于奥尼尔。
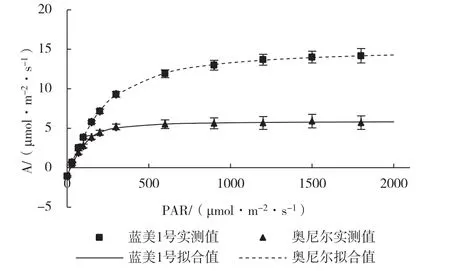
图3 2 个南高丛蓝莓品种的光合-光响应曲线
从表3 可以看出,蓝美1 号的最大净光合速率(Amax)和光饱和点(LSP)均显著高于奥尼尔,表明该品种具有较高的潜在光合能力,且对强光的适应能力较强。2 个南高丛蓝莓品种的光补偿点(LCP)、表观量子效率(AQE)、暗呼吸速率(Rd)没有显著差异。

表3 2 个南高丛蓝莓品种的光合-光响应参数比较
2.5 光合作用的CO2响应特征
如图4 所示,2 个南高丛蓝莓品种叶片净光合速率(A)对胞间CO2浓度(Ci)的响应趋势相似,均表现为A 随Ci 升高呈先迅速上升达饱和点后逐 渐趋于平缓,但相对而言,蓝美1 号的A 随Ci 上升的幅度明显大于奥尼尔。在相同Ci 条件下,蓝美1 号的A 高于奥尼尔,表明蓝美1 号叶片具有较高的核酮糖1,5-二磷酸(RuBP)羧化酶活性与较强的碳同化能力。

图4 2 个南高丛蓝莓品种的光合-胞间CO2响应曲线
由表4 可知,蓝美1 号的Rubisco 酶最大羧化速率(Vcmax)、电子传递链潜在最大电子传递速率(Jmax)、初始羧化效率(CE)、最大净光合速率(Amax)均显著高于奥尼尔,表明该品种具有较高的光合酶活性和光合碳同化能力。蓝美1 号的光呼吸速率(Rp)略高于奥尼尔,表明该品种通过光呼吸消耗过剩光能以减轻强光对光合机构伤害的能力较强。

表4 2 个南高丛蓝莓品种的光合-CO2响应参数比较
2.6 光合作用的温度响应特征
由图5 可看出,在叶片温度(Tl)为30~40 ℃时,蓝美1 号叶片净光合速率(A)随Tl升高缓慢降低,而奥尼尔叶片A 随Tl升高近似呈直线下降。在各Tl水平下,蓝美1 号叶片A 均显著高于奥尼尔。表明叶温升高对蓝美1 号光合性能的抑制作用较小,该品种具有较强的耐热性。

图5 2 个南高丛蓝莓品种的光合-叶温响应曲线
3 讨论与结论
本试验地土壤质地黏重,在试验观测当年夏季 气温≥35 ℃的高温天数为18 d。田间观测发现,与其他引进的南高丛蓝莓品种相比较,蓝美1 号植株生长势极强,枝条量多而健壮,说明其对黏重的黄棕壤和夏季高温有较强的适应性。
植物光合作用日变化是在一定天气条件下,各种生理生态因子综合效应的最终反应,其结果可作为分析生长或产量限制因素的重要依据[7]。本研究中2 个南高丛蓝莓品种净光合速率(A)日变化曲线为双峰型,即光合速率在上下午各出现1 次峰值,午间降低,表现出光合“午休”现象。这与陈薇[8]、王庆贺[9]、董丽华等[10]、古咸彬等[11]的研究结果相似。光合“午休”是植物对午间强光和高温的一种生态适应和自我调节机制的体现。造成植物叶片净光合速率下降的自身因素分为气孔限制因素和非气孔限制因素[12]。蓝美1 号气孔导度(Gs)、胞间CO2浓度(Ci)在午间也呈下降趋势,表明引起该品种叶片光合“午休”的主要因素是气孔限制因素。奥尼尔叶片A、Gs 午间下降,而Ci 升高,说明引起该品种光合“午休”的主要因素是非气孔限制因素,可能是午间强光、高温破坏了叶片光合机构,使光合磷酸化和电子传递受阻,或是影响了与光合有关的Rubisco(核酮糖-1,5-二磷酸羧化酶/加氧酶)等酶活性引起的[13-14],具体因素还需进一步研究。
植物光合作用日均值可以反映其光合能力的大小,植物水分利用效率(WUE)是评价植物生长适宜程度的综合生理生态指标,它实质上反映了植物耗水与其干物质生产之间的关系。本试验中南高丛蓝莓品种蓝美1 号叶片A 日均值和蒸腾速率(E)日均值均显著高于奥尼尔,而WUE 日均值显著低于奥尼尔,说明蓝美1 号具有高光合能力、高耗水特性,在栽培管理上必须保证充足的水分供给才能获得高产稳产。
分析研究光响应曲线可定量研究光合有效辐射与净光合速率的关系,获得最大净光合速率、光补偿点、光饱和点及暗呼吸速率等光合特征参数,有助于更好地了解光合作用过程[15]。本试验光响应拟合结果显示,蓝美1 号的光补偿点较奥尼尔略低,说明其对弱光的耐受能力稍强;蓝美1 号的表观量子效率略高于奥尼尔,说明其转化利用光能的效率略强;蓝美1 号的最大净光合速率(Amax)和光饱和点(LSP)均显著高于奥尼尔,表明该品种具有较高的潜在光合能力,且对强光的适应能力较强,在种植时可选择光照充足的地块栽培。
Rubisco 酶最大羧化速率(Vcmax)是Rubisco 酶表观羧化活性,其大小取决于Rubisco 酶的数量和活化程度,是表征植物光合能力高低的重要参数[16]。初始羧化效率(CE)与Rubisco 酶活性呈正相关[17]。本研究中,蓝美1 号的Vcmax、电子传递链潜在最大电子传递速率(Jmax)、CE、Amax均显著高于奥尼尔,表明其光合电子传递与Rubisco 活性高于奥尼尔,使其对CO2有更高的利用效率,因而具有较高的光合碳同化能力。光呼吸是耗散过剩光能,保护光合机构免于强光破坏的一个途径[18]。本研究中蓝美1号的光呼吸速率(Rp)略高于奥尼尔,这也可能是其更适应强光、高温环境的原因之一。
在蓝美1 号引种栽培中,光合温度适应性亦是重要考量指标之一[19-20]。本试验中,各叶片温度(Tl)水平下,蓝美1 号叶片A 均显著高于奥尼尔,且蓝美1 号叶片A 随Tl升高下降幅度较小,降低速度较慢,表明在高温环境下蓝美1 号光抑制程度较轻,自我调节能力强,因而其对高温的适应性较好,适宜在我国南方气候炎热地区栽培。
综上所述,与奥尼尔相比较,蓝美1 号具有较高的潜在光合能力,对强光有较强的适应能力,其对弱光的耐受能力亦稍强,在我国南方地区炎热气候条件下也具有“午休”的适应性调节机制,兼具较强的耐热性,可在我国南方地区推广种植。
