不同生境/萌发类型尖喙牻牛儿苗生物量分配特征
2022-04-15张媛媛孟欢欢周晓兵尹本丰周多奇
张媛媛, 孟欢欢, 周晓兵, 尹本丰, 周多奇, 陶 冶
(1.安庆师范大学生命科学学院,皖西南生物多样性研究与生态保护安徽省重点实验室,安徽 安庆 246133;2.中国科学院新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室,新疆 乌鲁木齐 830011)
短命植物(Ephemeral Plant),指生存于极端干旱环境且生命周期短的一类物种,在中国主要分布在新疆北部温带荒漠生态系统中,是春夏季荒漠植被主要构建者[1]。通常,4月是短命植物生长发育最主要的时期,同期沙漠降雨较多且伴着融雪水的产生,土壤表层含水量较高,因此短命植物的根系仅生长在浅土层即可获得生长发育所需的水分[2];此外,该时期也恰巧处于其他荒漠植物返青之前,大量的短命植物驻扎于裸地可保持荒漠地表稳定,维持荒漠地区物种多样性,进而促使了春夏季荒漠生态系统的稳定[1]。
生物量是植物的基本生物学特征和功能性状之一,是植物物质和能量的基本体现,而生物量分配则是指植物在生长发育过程中将同化的资源分配到各个器官的过程,是植物生存与生殖权衡产生的结果[3]。目前认为植物生物量分配特征存在种间和种内的差异,且不同物种的根、茎、叶以及繁殖器官在不同的环境条件和生长发育阶段存在显著差异。尽管如此,植物间依然存在一个相对普遍的生物量分配模式,即生长环境改变会导致生物量积累量及分配比例发生改变,而其分配关系是否变化则主要取决于植物基因型[4]。平衡生长假说认为植物通常会通过改变生物量分配来适应环境的改变,并维持自身最大生长速率;最优分配假说则认为,生物量应该被更多的分配给受资源限制的器官,以增加潜在的资源获取能力进而缓解胁迫[5]。大量研究证实了两种假说,即资源限制影响了生物量在植物各器官中的分配[6]。植物地上与地下生物量分配是生物量分配特征研究的重要组成部分[7],但目前的研究还不足以充分揭示短命植物这一特殊生活型植物在不同环境条件下的生物量分配模式和生存策略。
尖喙牻牛儿苗(Erodium oxyrhinchum)是古尔班通古特沙漠春季短命植物层片中的主要物种,于4—5 月开花、5—6 月结果,整个生命周期一般只需2—3个月,是典型一年生短营养期草本植物[8];更为特殊的是,在合适的水土和气温下,该物种也可以在其他季节萌发(即异时萌发),依据萌发时间不同可分为春萌、夏萌和秋萌3 种主要类型[9]。一般而言,夏季短期强降雨导致的夏萌株常因水资源快速枯竭而不能完成结实。除不同萌发时间类型外,尖喙牻牛儿苗特殊的尖喙状和羽毛状繁殖体组合可穿透地表坚硬的生物土壤结皮而广泛定居,其中不同类型生物结皮对尖喙牻牛儿苗生理特性具有显著的影响,并影响其生长发育[10-11]。那么,不同自然生境下尖喙牻牛儿苗地上和地下资源分配有何差异?目前已对短命植物尖喙牻牛儿苗开展了一系列研究,如繁殖体外部形态特征及其有性繁殖体生理适应性[8]、积雪和丛枝菌根真菌网络对尖喙牻牛儿苗幼苗的生长作用[12]、包含尖喙牻牛儿苗在内不同生活型草本植物生物量分配差异及其化学计量特征等[13],但对其地上和地下生物量分配策略在不同萌发类型及生物结皮类型间的差异性尚不明晰。为此,以3 种不同生境中的春萌和一种生境的夏萌尖喙牻牛儿苗为研究对象,系统阐述尖喙牻牛儿苗个体大小(生物量)在不同生境和不同萌发时间是否存在差异,地上-地下生物量分配比例是否会发生变化,是否具有一致的相关生长关系,以及个体大小是否影响其生物量分配模式。本研究将阐明尖喙牻牛儿苗在不同生境和萌发时间条件下的生物量分配特征,进一步揭示短命植物应对荒漠环境所采取的资源分配策略,为明晰短命植物的生存策略,实现荒漠生态系统的科学管理提供参考依据。
1 材料与方法
1.1 研究区概况
古尔班通古特沙漠(44°11′~46°21′N,84°31′~90°00′E,面积4.88×104km2)位于新疆北部准噶尔盆地腹心,是我国最大的固定和半固定沙漠;该沙漠年平均降水量在150 mm以下,沙漠中部年降水量约为70~120 mm;年潜在蒸发量约为2000~2800 mm。年均温6~10 ℃,极端高温超过40 ℃,极端低温在-30 ℃以下;≥10 ℃的活动年积温可达3000~3500 ℃,空气相对湿度平均50%~60%,5—8月通常在45%以下[1]。该沙漠植物群落的主要建群种有梭梭(Haloxylon ammodendron)、准噶尔沙蒿(Artemisia songarica)、膜果麻黄(Ephedra przewalskii)、白梭梭(Haloxylon persicum)等;而短命和类短命植物是该沙漠早春时节优势物种[14]。此外,该沙漠地表还生长着大量生物结皮,主要有藻结皮、地衣结皮和藓结皮,结皮生境土壤含水量与裸沙相比较低,不同类型生物结皮是荒漠生态系统重要组成部分[15-16]。
1.2 野外样品采集
于2013 年5月10 日和7 月20 日2 个时期,在古尔班通古特沙漠腹地对尖喙牻牛儿苗植株进行采样,春萌株大约在4月初萌发,5月10日左右处于花果期,此时设置裸沙春萌(Bare Sand-Spring Germination,BS)、藻结皮春萌(Algal-Spring Germination,AS)和地衣结皮春萌(Lichen-Spring Germination,LS)3种结皮/生境类型的调查样地,每个样地设置3个大小为5 m×5 m的重复样方。在每个样方内选取若干完整且健康的植株,采用全株挖掘法获取整株。当年6月下旬一场较大降雨后,尖喙牻牛儿苗迅速萌发,即为夏萌株。为与5月采集的不同生物结皮生境中的样品进行对比,以7 月20 日始花期为节点,对尖喙牻牛儿苗夏萌株开展调查取样。因夏萌株几乎仅分布于裸沙区,故选择裸沙夏萌(Summer Germination in Bare Sand,SG)样地开展研究,设置3 个5 m×5 m 的调查样方,样品采集与处理方法同上。因秋萌株通常可越冬,已不属于严格意义上的短命植物范畴,因此未在本文论述。为方便表述,下文将不同生境及萌发时间的尖喙牻牛儿苗统称为4种类型,在3种结皮类型比较时称之为“生境”,在其与夏萌SG植株比较时,称之为“类型”。4种类型尖喙牻牛儿苗的最终样本量分别为30株、72株、30株和43株。将植株鲜样放入标记好的信封,置于保温箱内带回实验室作进一步处理。
1.3 生物量的计算
对采集回的植株鲜样进行处理,将其地上、地下成熟组织分开装入信封袋,并置于烘箱内70 ℃烘24 h。用精度为0.0001 g 的天平对烘干后的样品进行称重,得到每株植物的地上生物量(AGB,g)、地下生物量(BGB,g)和总生物量(TB,g),计算根冠比(R/S)。各生物量指标均采用平均值±标准误(Mean±SE)的形式给出。
1.4 数据统计与分析
对4 种类型尖喙牻牛儿苗的地上生物量、地下生物量、总生物量、根冠比分别进行单因素方差分析。R/S 随着个体大小(地上生物量、地下生物量、总生物量)的变化趋势通过幂函数来判断,变化趋势的显著度用曲线估计进行分析。常规数据分析和作图在Excel 2016 和Origin 9.0 中完成,方差分析和曲线估计在SPSS 19.0中实现。
对4 种类型尖喙牻牛儿苗地上、地下及总生物量进行异速生长分析和等速生长检验。异速生长关系公式为Y=βXα,其中,Y是某种生物学特征或者功能,β是标准化常数,X指个体大小,α是异速生长指数。确定异速生长参数时,须将Y=βXα转换为logY=logβ+αlogX的形式,并用降主轴回归(RMA,即Model Type Ⅱ)方法计算回归模型的指数、95%置信区间(95% Confidence Interval,CI)及决定系数(R2)[3,17]。对数化之后的α即为幂函数对数化之后线性回归的斜率,logβ为线性回归的截距。上述异速生长指数(α)和截距(logβ)的计算、等速生长检验及α的方差分析均使用SMATR软件完成。
2 结果与分析
2.1 不同类型尖喙牻牛儿苗的生物量及其分配比例
通过单因素方差分析可知,4 种类型尖喙牻牛儿苗的地上生物量(AGB)、地下生物量(BGB)和总生物量(TB)均差异明显(P<0.05),生物量在4 种类型中大小关系均为:BS>AS>LS 和SG(图1),且尖喙牻牛儿苗AGB 均明显大于BGB。LS 生境中AGB(0.1203±0.0067)g、BGB(0.0164±0.0015)g、TB(0.1367±0.0095)g 和SG 类型植株AGB(0.0553±0.0031)g、BGB(0.0103±0.0131)g、TB(0.0655±0.0079)g之间的差异均不显著(P>0.05);BS 生境中BGB(0.0988±0.0135)g、TB(1.0058±0.0777)g与AS生境BGB(0.0865±0.0148)g及TB(0.0655±0.0079)g 均无显著差异(P>0.05),但BS、AS 的BGB、TB 与LS、SG 2 种类型的BGB、TB 之间存在显著差异(P<0.05)。BS 与AS 2 种生境AGB之间差异显著(P<0.05),BS、AS与LS及SG 2种类型的AGB 也存在显著差异(P<0.05)。BS 生境下单株TB 为1.0058 g,是AS 和SG 的10 倍多。4 种类型尖喙牻牛儿苗根冠比(R/S)表现为SG=AS=LS>BS的特征(图1d),其中SG 类型植株R/S 为0.1877±0.0046,BS 生境R/S 为0.1106±0.0031,表明SG 条件下植株根生物量分配比例最大而BS 生境植株的根生物量分配比例最小。

图1 4种类型尖喙牻牛儿苗地上生物量(AGB)、地下生物量(BGB)、总生物量(TB)及根冠比(R/S)特征Fig.1 Characteristics of aboveground biomass(AGB),belowground biomass(BGB),total plant biomass(TB),and root to shoot mass ratio(R/S)of Erodium oxyrhinchum of four types
2.2 不同类型尖喙牻牛儿苗地上-地下生物量间的相关生长关系
4 种类型尖喙牻牛儿苗地上-地下生物量(AGB-BGB)(指X-Y,下同)、地上-总生物量(AGBTB)、地下-总生物量(BGB-TB)均具有极显著(P<0.01)的相关生长关系(图2,表1),SG和BS 2种类型尖喙牻牛儿苗的AGB-BGB、AGB-TB、BGB-TB 以及LS 生境BGB-TB 之间均符合等速生长关系;AS 和LS 2 种生境中的AGB-BGB、AGB-TB 相关生长指数(α)显著小于1.0(P<0.01),AS 生境中BGB-TB 则呈相关生长指数(α)显著>1.0的异速生长关系。尽管如此,4 种类型AGB-BGB 间及BGB-TB 间的斜率差异并不显著(P>0.05),且都具有共同斜率,其中AGB-BGB 间共同的α为0.8843,BGB-TB 间共同的α为1.101,前者为α<1.0的异速生长关系(P<0.05),而后者则为α>1.0的异速生长关系(P<0.05)。这表明,尖喙牻牛儿苗总体上表现为AGB 分配速率高于BGB 的特征,体现出一定的保守性,但生物结皮对其生物量分配速率有一定影响。
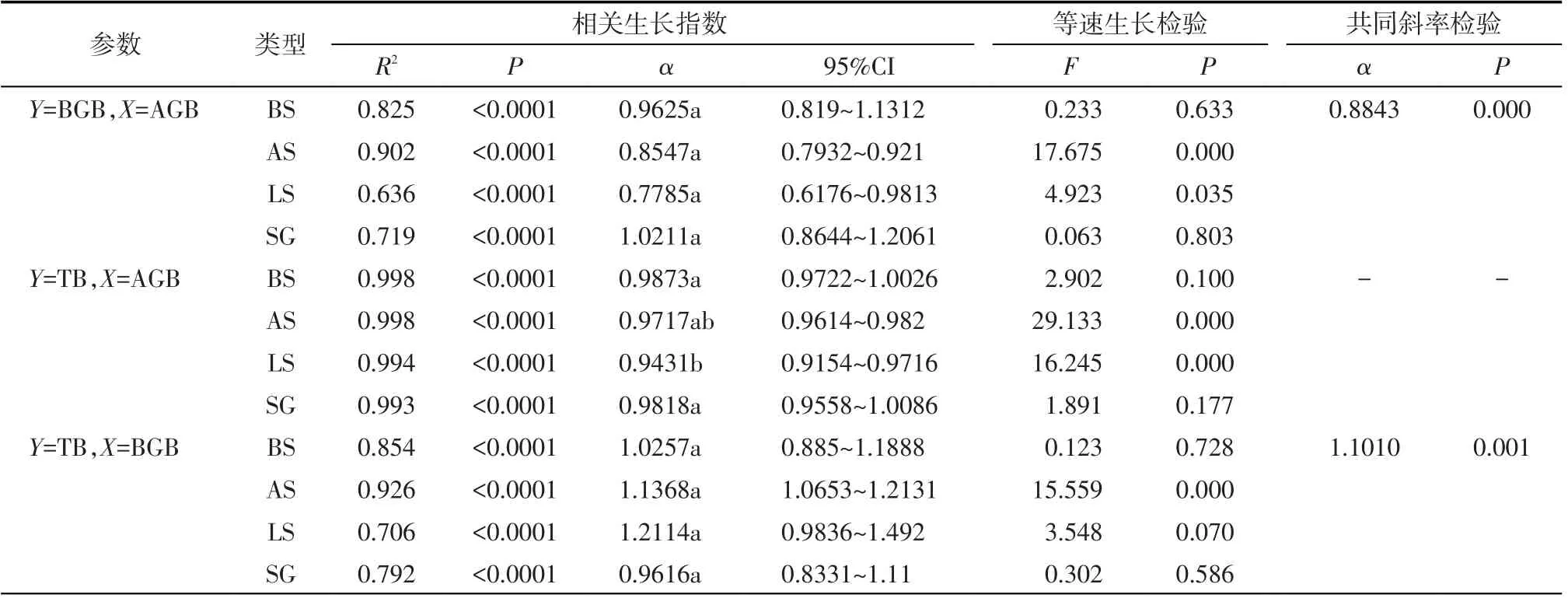
表1 4种类型尖喙牻牛儿苗生物量指标间的相关生长指数、等速生长检验及共同斜率检验Tab.1 Allometric relationships the test of allometry and the test of common slope among biomass indices of Erodium oxyrhinchum of four types

图2 4种类型尖喙牻牛儿苗单株地上生物量(AGB)、地下生物量(BGB)及总生物量(TB)间的RMA线性拟合关系Fig.2 linear regression(RMA)plots of aboveground biomass(AGB),belowground biomass(BGB)and total plant biomass(TB)of Erodium oxyrhinchum of four types
2.3 不同类型尖喙牻牛儿苗生物量分配比例与个体大小的关系
随尖喙牻牛儿苗单株AGB、BGB、TB的增加,各类型根冠比(R/S)呈现不同的增长或下降趋势(图3)。具体来看,随AGB增加,4种类型单株R/S均呈下降趋势,其中AS 和LS 生境的下降趋势达到极显著水平(P<0.01)。AS 生境植株R/S 在AGB 较小时下降趋势较快,随AGB 增大其R/S 下降趋势逐渐减缓。R/S 与地下生物量的关系在4 个生境中均不相同,其中SG类型植株R/S随BGB增大呈显著增大趋势(P<0.05),而AS 生境中R/S 则呈显著下降趋势(P<0.05)。对于TB而言,4个类型中R/S均随TB的增大而呈下降趋势,其中AS 和LS 2 种生境的下降趋势达到极显著水平(P<0.01)。可见,短命植物尖喙牻牛儿苗地上与地下生物量分配比例的变化不仅与生境有关,植株个体和器官大小也会对其产生影响。

图3 4种类型尖喙牻牛儿苗根冠比R/S随个体大小(地上生物量、地下生物量、总生物量)的变化趋势Fig.3 The change trends of root to shoot mass ratio(R/S)along with plant size(aboveground biomass,belowground biomass and total plant biomass)of Erodium oxyrhinchum of four types
3 讨论
3.1 不同类型尖喙牻牛儿苗的生物量分配特征
生境发生变化时,植物往往会产生生物量的积累以及分配的调整,体现了植物对环境变化的响应与适应性特征,也体现了植物较强的表型可塑性[4]。干旱、半干旱的温带荒漠生态系统通常具有极端温度及水分供给不足的特点,其维管植物稀少,养分周转效率差,沙面常被生物结皮所覆盖[18]。然而,作为体型较小的草本植物,尖喙牻牛儿苗却拥有特殊的繁殖体结构特征,可以在生物土壤结皮上定居[8]。目前Godínez-Alvarez[19]、Li[20]、张元明[21]等研究团队都曾研究过荒漠生物结皮与维管植物生长发育之间的关系,结果发现,在不同环境条件及生长阶段,生物结皮对植物生长发育影响不同,生物结皮可提高土壤养分,在一定程度上促进植物生长发育;但是,土壤氮素动态及植物碳累积在很大程度上取决于水资源的可获得性[22],有结皮与无结皮生境植物生物量差异主要与土壤水分含量有关[23]。Kidron等[16]研究了内盖夫沙漠不同生境中一年生植物的生长状况,发现在极端干旱条件下,结皮区土壤含水量低于裸沙区,且结皮区的植物生长状况较次,而无结皮沙丘在极端干旱年份则可作为一年生植物的汇集区。Zhuang 等[11]研究了古尔班通古特沙漠生物结皮对3 种短命植物的生长影响,发现不同植物具有不同的响应特征,尤其在植物生长后期,结皮生境土壤含水量显著低于裸沙生境,导致结皮生境尖喙牻牛儿苗与硫苞菊(Hyalea pulchella)生物量显著低于裸沙生境。
本研究中,BS生境尖喙牻牛儿苗地上、地下及总生物量均大于AS和LS生境,该结果与Kidron等[16]及Zhuang等[11]的研究结果相似。通常,水分是影响干旱区植物生长的主要限制性因素[16],而生物结皮能加快表层土壤蒸发率,阻碍土壤水分入渗[16],导致藻结皮和地衣结皮土壤含水量低于裸沙;同时,生物结皮还会与维管植物竞争水资源,反过来抑制植物生长[10]。可见,本研究中2 种结皮生境AS 和LS生境尖喙牻牛儿苗的生物量与BS 生境植株存在差异,极有可能是土壤含水量不同导致。多年观察和研究发现,尖喙牻牛儿苗种子在土壤水分充足的情况下便可萌发,夏季沙漠一场临时的降雨也可能达到尖喙牻牛儿苗种子萌发所需的水分阈值,导致大量夏萌株出现。但短暂的水资源并不能满足其整个生命周期的需求,随着后期气温升高、土壤干旱化加剧,加之多年生植物与其竞争养分与水分,导致SG类型的尖喙牻牛儿苗无法正常发育,因此其个体生物量远小于BS 和AS 生境的春萌株。可见,生物结皮(生境)或季节(萌发时间)导致的水分限制是影响尖喙牻牛儿苗生长发育的直接因素。
一般而言,植物倾向于将生物量分配给资源短缺的器官,导致不同器官之间分配比例发生变化[5]。根系是植物吸收养分和水分的主要构件,也是最先感受逆境胁迫的部位之一,干旱通常迫使植物会将更多的资源分配给根部,以增加植株吸收养分和水分的能力[23]。研究发现,随生境土壤水分含量降低,砂蓝刺头(Echinops gmelini)根冠比呈逐渐增大趋势[24]。对入侵植物胜红蓟(Ageratum conyzoides)种群构建生物量研究发现,其根冠比在不同生境条件下存在显著差异,相对干旱的生境中胜红蓟为适应干旱和水分胁迫,会加强其根部的投资,增强根系获取生长所需的水分和无机盐[25]。由于BS 生境土壤水分相对充足,尖喙牻牛儿苗无需往根部投入更多资源;而AS与LS生境下尖喙牻牛儿苗以及SG类型的尖喙牻牛儿苗由于受到的水分胁迫较大[11,26],导致植株会投入更多的资源到根部,进而造成不同生境间植物根冠比出现差异。本研究结果还有一点较为特别,即AS、LS 2 种生物结皮生境中尖喙牻牛儿苗生物量虽然低于BS 生境,但是AS 生境的总生物量及地下生物量却与BS生境无显著差异,这可能是藻结皮的固氮作用抵消了一部分水分胁迫带来的生长抑制作用[27]。
3.2 不同类型尖喙牻牛儿苗构件生物量间的相关生长关系
相关生长关系是指生物体的某些生物学特征与个体大小之间的幂函数关系,且能够揭示生物界各物种器官生物量分配的基本速率问题[28]。相关生长关系与研究尺度无关,在生物界中普遍存在[3,13,28]。大量研究表明物种间存在一个相对普遍的生物量分配模式,如全球木本植物地上、地下生物量均为等速生长关系,而其他生物学特征与构件生物量间多呈固定的异速生长关系;其中等速生长关系体现了植物构件间具有恒定的分配速率,而异速生长关系则体现植物体资源分配的不均匀性[29]。但也有学者认为植物在受到自身基因限制以及响应自然选择压力下,会进化出诸多异速生长模式[29]。
目前关于荒漠植物地上地下生物量分配对环境的响应已经从个体、种群、群落、生活型等多个方面进行了研究[13,21,25]。总结发现,不同物种、不同生活型或同一物种不同生长期的植株,其地上-地下生物量分配存在相似之处但无统一规律。研究表明,除了植物本身遗传因素外,植物各器官间的生物量分配也是由环境条件驱动,植物通常以改变资源分配策略来响应环境变化[17,30]。本研究中,4个类型尖喙牻牛儿苗地上与地下生物量均具有极显著的相关生长关系,其中BS 与SG 类型的尖喙牻牛儿苗植株地上-地下生物量的相关生长指数95%置信区间都包括1.0,即呈等速生长关系,符合理论预测;而AS与LS生境的尖喙牻牛儿苗则均呈地上资源分配速率大于地下的异速生长关系。有研究表明,水分差异虽然导致荒漠植物地上与地下生物量比值存在差异,但并没有改变其地上与地下的生物量分配模式或分配速率[31],如红砂(Reaumuria soongorica)地上、地下生物量在不同水分处理间存在差异,但地上-地下生物量的分配模式并未改变[32]。这与本文AS 和LS 生境的研究结果相似,这2 种结皮生境尖喙牻牛儿苗地上、地下或总生物量虽差异显著,但其根冠比及相关生长指数却无显著差异。而且,尖喙牻牛儿苗属短命植物,短命植物在种子萌发当年就可以完成生长发育的全部环节;在有限的生命周期里,短命植物往往会将大部分同化产物分配给地上组织,用于营养生长和繁殖,因而一般不会分配更多生物量到地下,不必将能量保存于类似多年生植物的根系中[30,33]。可见,AS和LS 2种结皮生境下尖喙牻牛儿苗的生物量分配模式与其生活史策略是相符的。但同为春萌株,裸沙BS生境尖喙牻牛儿苗地上-地下生物量间为等速关系,而藻结皮AS 与地衣结皮LS 为异速关系的原因,可能由于水是干旱区植物生长的主要限制性资源,BS生境含水量较后者更为充足[11],资源限制缓和导致各器官间的竞争也削弱,因此BS 生境植株表现出等速生长,这反映了植物在适应环境过程中资源之间的平衡[34]。
SG类型的植株地上-地下生物量间相关生长关系虽同BS生境一样呈等速生长关系,但在种间竞争与水分的双重胁迫下,其生长状态远次于后者。有学者认为,当环境不利于植物生存时,植株的生长发育易受限制,并产生个体变小、无法正常结实、种子数量减少等现象,而植株在遭受过度胁迫(伤害)后的种种表现,很难被认为是对环境的一种适应性反应[35]。从SG类型的尖喙牻牛儿苗个体最小、无法结实等特点来看,其受到的生存压力远大于前3 种生境下的植株。最优分配理论认为植物倾向于将生物量分配到资源最受限制的器官[5]。SG 类型的植株为在恶劣生境中生存,故而改变了原有的生活史策略,将资源更多地分配给根系,以提升其竞争水分和养分的能力来保持生存;也正因如此,SG 尖喙牻牛儿苗把资源更多地分配给地下而表现出地上-地下等速生长格局。尽管4 种类型尖喙牻牛儿苗地上-地下生物量相关生长关系存在差异,但却拥有相同的异速生长指数0.8843,表明尽管生存条件不同,生物量之间也有差异,但其具有相同资源分配的内在共性,这可能与其物种固有特性有关[29]。
3.3 尖喙牻牛儿苗生物量分配与个体大小的关系
个体发育影响植株不同构件间的资源分配比例,即生物量分配存在显著的个体发育漂变现象[36]。有学者指出荒漠植物生物量分配不仅与物种相关,与植物个体大小也息息相关[32]。植物根冠比与繁殖比通常是植物生物量分配研究中常选的功能性状,其中根冠比是最能直接反应植物生物量的分配特征[37]。研究表明,大部分陆生草本植物分配给根部的生物量随个体增大而减小[38]。对荒漠短命植物、类短命植物及一年生长营养期植物根冠比与个体大小的关系进行分析表明,其根冠比均随个体增大而呈异速减小趋势,且这种趋势在个体较小时更为明显[29]。
在本研究中,4 种类型尖喙牻牛儿苗根冠比与个体大小之间的关系存在差异,该结果与上述结果部分相似。BS 与SG 2 种类型植株的根冠比随植株个体增大而下降,但趋势不显著,这与其地上-地下生物量为等速生长关系相符;AS 与LS 生境下的植株根冠比则随个体增大呈显著或极显著下降趋势,这表明,在生物结皮存在的条件下,尖喙牻牛儿苗地上-地下生物量分配存在个体发育漂变现象。植物可以通过调节生物量分配来达到生长最大化,以应对变化的环境[37]。本研究中,4 种类型尖喙牻牛儿苗个体大小和分配比例存在差异,其原因可能是植株受到了不同程度的水分胁迫所致。BS与AS生境相比,尖喙牻牛儿苗的生物量差别不大,但是2种生境下的根冠比随个体增大的变化趋势却有明显不同;这可能是由于AS 与LS 生境中土壤含水量均较裸沙生境低,故相对于裸沙生境而言,2种结皮生境个体较小的尖喙牻牛儿苗会将更多的生物量分配到根系,以促进根系生长、增强植株获取水分的能力。在这个过程中,个体较大的植株为保证有性繁殖和种群扩散,其生物量将更多地分配到地上部分。对于SG类型的尖喙牻牛儿苗而言,其生境条件是4种类型中最为恶劣的,严酷的环境可能改变了其生活史策略,表现出根冠比不受个体大小的影响。可见,本研究结果证实了Hutchings和John的观点,即个体发育以及环境条件对生物量分配均有影响[39]。
综上可知,尖喙牻牛儿苗在不同生境及萌发时间条件下均受到了不同程度的水分胁迫,导致其个体生物量及分配比例存在差异,其地上-地下相关生长关系也并不遵从单一的、规范的生物量分配模式。尖喙牻牛儿苗在环境胁迫下可通过对生物量的调节分配来维持其最优生长,其根冠比与个体大小的关系也不是固定不变的,环境对其不同个体水平地上-地下部分生物量分配比例具有一定的影响作用。因此,尖喙牻牛儿苗在适应不同荒漠环境过程中形成了可塑的资源分配策略,研究结果对深入揭示荒漠短命植物生活史策略具有一定科学意义。
4 结论
通过对生长在不同生境和萌发时间条件下4个类型的短命植物尖喙牻牛儿苗地上地下生物量分配特征进行分析,得出以下结论:
(1)尖喙牻牛儿苗单株地上、地下及总生物量总体表现为BS>AS>LS=SG的格局,水分胁迫是导致不同类型植株生物量产生差异的主要因素;尖喙牻牛儿苗通过调节根冠比以适应干旱和水分胁迫,其根冠比呈SG=LS=AS>BS的特征。
(2)BS 与SG 类型的尖喙牻牛儿苗地上地下等速生长关系,AS 与LS 生境的尖喙牻牛儿苗呈地上资源分配速率大于地下的异速生长关系,水分胁迫及短命植物生活史策略共同制约着尖喙牻牛儿苗地上地下资源分配速率,但不同生境及萌发时间条件下尖喙牻牛儿苗生物量分配存在一定的内在共性。
(3)个体发育及环境条件对生物量分配均有影响,AS 与LS 生境尖喙牻牛儿苗根冠比随个体增大均呈异速减小趋势,而BS 与SG 类型植株根冠比则保持相对恒定。
