放牧强度对羊草草甸草原植物器官及群落氮磷化学计量的影响
2022-04-14王淼张宇李瑞强辛晓平朱晓昱曹娟周忠义闫瑞瑞
王淼,张宇,李瑞强,辛晓平,朱晓昱,曹娟,周忠义,闫瑞瑞
放牧强度对羊草草甸草原植物器官及群落氮磷化学计量的影响
王淼1,张宇1,李瑞强2,辛晓平1,朱晓昱3,曹娟1,周忠义4,闫瑞瑞1
1中国农业科学院农业资源与农业区划研究所/呼伦贝尔草原生态系统国家野外科学观测研究站,北京 100081;2内蒙古自治区环境在线监控中心,呼和浩特 010055;3农业农村部环境保护科研监测所,天津 300191;4牙克石市林业和草原局事业发展中心,内蒙古牙克石 022150
【】以地上植物茎、叶以及群落为研究对象,探究不同放牧强度对植物器官以及群落碳、氮、磷含量及化学计量特征的影响,为深入解析放牧干扰下草地生态系统退化过程,促进草地放牧适应性管理提供科学依据。基于呼伦贝尔羊草草甸草原长期控制放牧试验平台,在6种不同放牧强度(不放牧G0.00、较轻度放牧G0.23、轻度放牧G0.34、中度放牧G0.46、重度放牧G0.69、极重度放牧G0.92)下采集植物群落、植物不同器官的样品,采用元素分析法测定碳、氮、磷含量,并分析其碳、氮、磷含量及计量比在不同放牧强度下的变化。随着放牧强度的增加,植物茎、叶和群落的碳含量呈现显著的线性下降,重度-极重度放牧显著低于不放牧(<0.05);群落氮、磷含量呈现显著的线性增加,重度-极重度放牧显著高于不放牧(<0.05);碳含量茎叶比以中度放牧G0.46处理最低、氮含量茎叶比以重度放牧G0.69处理最高、磷含量茎叶比以较轻度放牧G0.34处理最高。植物不同器官碳含量及氮磷比的变异系数较小,氮、磷含量的变异系数较大, 不同器官的碳含量及氮磷比变异系数变化范围分别为4.01%—5.74%和2.84%—8.54%,植物氮、磷含量变异系数范围为11.47%—14.96%和11.13%—22.88%,碳氮比和碳磷比的变异系数范围分别为10.64%—16.00%和8.88%—13.57%,其中,植物叶片氮磷比的变异系数最小,植物茎磷含量的变异系数最大。轻度放牧G0.34和极重度放牧G0.92时叶片的氮磷比介于14—16,受到氮、磷元素的共同限制,其余在不同放牧强度下均低于14,表现为受到氮的限制。放牧强度与植物茎、叶、群落的碳、碳氮比以及茎和群落的氮磷比呈现出极显著负相关,与植物茎、叶、群落的氮含量,叶和群落的磷含量以及茎的氮磷比呈现极显著正相关。土壤温度与植物茎中磷含量呈现显著正相关。土壤水分与植物叶、群落的碳含量,茎、叶、群落的碳氮比,茎和群落的碳磷比呈显著正相关;与茎、叶、群落的氮含量和叶、群落的磷含量呈现显著负相关。放牧使得植物碳、氮、磷含量,生态化学计量比及群落碳氮磷总含量发生了明显的变化,过度放牧不仅降低了植物茎、叶、群落的碳含量、碳氮比和碳磷比,还降低了群落碳氮磷总含量,增加了植物茎、叶、群落的氮、磷含量,以及植物茎、叶、群落的氮磷比,适度放牧降低了茎叶碳氮比,增加了茎叶碳磷比和氮磷比,适度放牧有利于促进草地生态功能可持续稳定发展。
草甸草原;放牧强度;碳、氮、磷含量;生态化学计量特征;茎叶;群落;呼伦贝尔
0 引言
【研究意义】我国的草原生态系统是欧亚大陆温带草原生态系统的重要组成部分,它既是重要的生态屏障,又是重要的畜牧业生产基地[1]。但是近年来,由于多年过度气候变暖等,全球草原生态系统遭受退化,其中我国90%左右的天然草原发生不同程度的退化。放牧作为草原最主要的利用方式,通过家畜的采食、践踏以及排泄影响着生态系统养分的循环与利用,植物通过调整自身的养分利用策略来适应不同放牧程度所产生的威胁。【前人研究进展】植物最重要的器官叶和茎对其生长、发育和生活史的完成都有极其重要的意义,在草原生态系统中,动物通过对植物的叶片和茎的采食和践踏对其产生影响。牛得草[2]对黄土高原天然草地建群种植物长芒草地上组织的研究中发现,放牧草地地上氮、磷含量增加的原因是家畜采食,其不仅能刺激植物茎、叶再生长,还可以促进地上植物元素循环,导致氮、磷转向幼嫩器官,且氮的转移量大于磷。许雪贇等[3]在西藏那曲进行的放牧与围封研究表明,围封显著降低植物叶片磷含量,显著增加植物叶片碳含量,对氮含量影响不显著。郑伟[4]在荒漠草地上研究围栏封育对其资源分配格局的影响,认为封育后生殖枝与叶相反,氮含量显著增加,而对碳含量影响不显著。丁小慧等[5]对草甸草原的研究表明,放牧对植物叶碳氮含量无显著影响。FRANK等[6]对美国黄石公园近半个世纪的草地连续放牧研究结果显示,放牧增加了氮含量而对磷含量无明显影响,从而提高了枝条氮磷比。HAN等[7]研究全国753种植物叶片的碳氮磷元素含量后认为,叶片氮含量与叶片磷含量呈显著正相关,植物体中的氮、磷元素是协同元素,具有相似的功能,一般呈正相关关系。而李香真等[8]和张婷等[9]对不同放牧强度下植物个体和小叶章养分及化学计量比的研究发现,随着放牧强度的增加,碳氮比增加先降低后升高,在中度放牧区碳氮比达到最低值。2001年李香真等[10]对旱黄梅衣的研究结果同样表明,8月份植物茎的碳氮比的最低值出现在中度放牧区,且叶和植物全株的碳氮比变化相类似。【本研究切入点】草原生态系统植物-土壤生态的化学计量特征受自然因素经纬度、区域尺度、草原类型、温湿度和人为干扰放牧利用方式、利用强度的影响而有所不同。BARDGETT等[11]提出了关于放牧对群落水平碳、氮、磷含量及其化学计量特征的假说,然而,目前国内外在不同放牧干扰下,植物不同器官水平间生态化学计量特征及其内在联系方面仍然知之甚少。【拟解决的关键问题】基于此,本研究依托呼伦贝尔草原生态系统国家野外科学观测研究站大型控制放牧实验平台,基于11年连续放牧活动,开展不同放牧强度下植物茎、叶、群落的碳、氮、磷含量以及生态化学计量特征的变化研究,探讨草甸草原生态系统不同器官对放牧的响应过程及适应策略,旨在为草原生态系统的恢复和放牧管理提供基础依据。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古呼伦贝尔市谢尔塔拉牧场场部东3 km,试验基地依托呼伦贝尔草原生态系统国家野外科学观测研究站长期控制放牧实验平台(49°32′—49°34′N,119°94′—119°96′E),海拔约670 m,属温带半干旱大陆性气候,年平均降水量约400 mm,年均气温-5—-2℃,≥10℃年积温1 580— 1 800℃,无霜期约110 d[12]。试验区土壤为黑钙土或栗钙土;植被类型为羊草+杂类草草甸草原,主要物种有羊草()、贝加尔针茅()、羽茅()等,伴生种有柴胡()、糙隐子草()等,主要退化指示种包括冷蒿()、二裂委陵菜()等。
1.2 研究方法
1.2.1 试验设计及样品采集 放牧试验设6个水平的放牧强度处理,载畜率分别为0.00、0.23、0.34、0.46、0.69和0.92 Cow.Au·hm-2(其中Au以500 kg肉牛为一个标准家畜肉牛单位),分别表示为不放牧G0.00、较轻度放牧G0.23、轻度放牧G0.34、中度放牧G0.46、重度放牧G0.69、极重度放牧G0.92,每个处理3次重复(图1),试验区围成18个面积为5 hm2的试验小区,试验区总面积90 hm2(图1)。在试验区面积一定,放牧时间相同条件下,用250—300 kg的放牧肉牛头数来控制不同放牧强度的实施,6个放牧强度肉牛头数分别为0、2、3、4、6、8头,肉牛数量共69头。试验于2009年开始,每年6月初开始放牧,10月初终止放牧,为期4个月。整个放牧期间牛全天在试验样地,不补充饲料,保证充足的饮水和盐分供给。2020年不同放牧强度土壤理化性状情况见表1。
2019年和2020年的生长盛期8月份,在不同的放牧处理中随机选取5个1 m×1 m样方开展取样,每个样方之间最少间隔20 m,共计选用样方90个。在样方内采用齐地面剪割法,将每个样方内的植物样品带回室内,新鲜样品105℃杀青 15 min,然后降温至65℃烘干12 h后称重,烘干后的样品用球磨仪充分磨碎后一部分测定茎叶元素含量,另一部分用于测定群落元素含量,采用化学方法测定植物有机碳、全氮、全磷。采用土钻法采集0—10 cm土层土壤样品用烘干法测定水分、采用地温计测定土壤温度,重复3次。
1.2.2 养分的测定及数据处理 土壤、植物有机碳采用重铬酸钾氧化-外加热法进行测定;土壤、植物全氮采用半微量凯氏定氮法进行测定;土壤全磷采用氢氧化钠熔融法、植物全磷采用硫酸-过氧化氢消煮-钼锑抗吸光光度法测定[13]。

上行字母为“小区编号”,其中 W—西面,M—中间,E—东面;下行数字和字母为“家畜放牧强度”(肉牛当量)
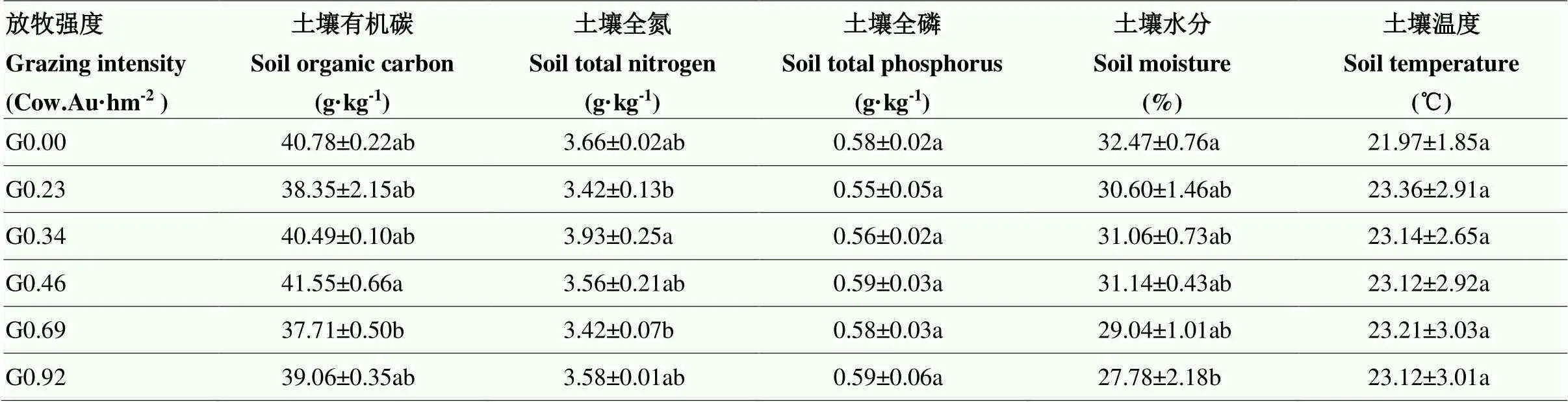
表1 2020年不同放牧强度下土壤理化性状
不同字母表示显著差异(<0.05),相同字母表示差异不显著(>0.05)
Different letters indicate significant difference (<0.05), while same letters indicate no significant difference (>0.05)
采用Excel 2017对数据进行整理,运用SPSS21软件进行统计分析,采用单因素方差分析(one-way ANOVA)法对不同放牧强度下的植物茎、叶、群落有机碳、全氮、全磷含量及其化学计量特征进行方差分析,并用LDS和Duncan法进行平均值之间的多重比较,显著性水平设为<0.05,极显著性水平设为<0.001,利用Origin2017进行绘图。变异系数(CV)(%)=标准差(SD)/均值×100。
2 结果
2.1 不同放牧强度下植物茎叶碳氮磷含量及其计量比的变异特征
由表2可知,两年均值植物各器官碳含量的变异系数整体较小,介于3%—6%之间;不同放牧强度间植物茎的碳含量变化范围为345.14—425.42 g·kg-1。植物各器官氮含量两年均值的变异系数均大于10%,介于11%—15%之间,植物叶片的氮含量变化范围为18.56—32.68 g·kg-1,其变异系数最高,茎的变异系数较低,群落最低为11.47%。磷含量的变异系数表现为植物茎最高,达到22.88%,且远高于叶(11.13%)和群落(12.57%)的变异系数,不同放牧强度间植物叶中磷含量最大,均值为1.6 g·kg-1。总体上,茎中磷含量的变异系数最大,叶片中氮含量的变异系数最大,群落磷含量的变异系数最大。
不同放牧强度植物器官之间计量比呈现出明显的变化(图2),植物茎、叶、群落的C﹕N、C﹕P均值随着放牧强度的增加显著降低(<0.05)。其中,与不放牧相比,放牧强度G0.46—G0.92茎和群落的C﹕N分别显著降低了28.22%—37.24%、12.92%—27.80%,极重度放牧G0.92叶中C﹕N显著降低了30.04%。植物叶C﹕P均值G0.92处理显著低于G0.34(<0.05),降低了24.7%,群落C﹕P均值G0.92显著低于G0.00(<0.05),降低了28.36%。植物茎、叶、群落的N﹕P均值随着放牧强度的增加无显著差异(>0.05),分别在放牧强度为0.69、0.92和0.92 Cow.Au·hm-2时达到最大值,为11.67、15.62和13.80。
2.2 不同放牧强度下植物茎叶碳氮磷元素含量的响应特征
2019年和2020年植物茎、叶、群落的碳含量以及茎叶比呈现不同的变化趋势(图3)。不同放牧强度下植物碳含量变化范围茎为347.25—415.69 g·kg-1,叶为345.48—380.87 g·kg-1,群落为359.04—401.81 g·kg-1,茎叶比为0.94—1.12。2020年不同放牧强度植物茎碳含量均高于2019年,2019年植物茎碳含量随着放牧强度的增加呈现显著的线性下降(2=0.97,<0.001);2020年植物茎碳含量(2=0.61,>0.05)在各放牧强度间无显著差异(>0.05)。2019年植物叶碳含量在各放牧强度间呈现显著线性回归(2=0.81,<0.05);2020年植物叶的碳含量随着放牧强度的增加呈现显著的线性下降(2=0.83,<0.05)。2019年(2=0.74,<0.05)和2020年(2=0.80,<0.05)植物群落的碳含量均随着放牧强度的增加呈现显著的线性下降。碳含量茎叶比在各放牧强度间呈现显著二项式回归(2=0.96,<0.05),在中度放牧G0.46最低。
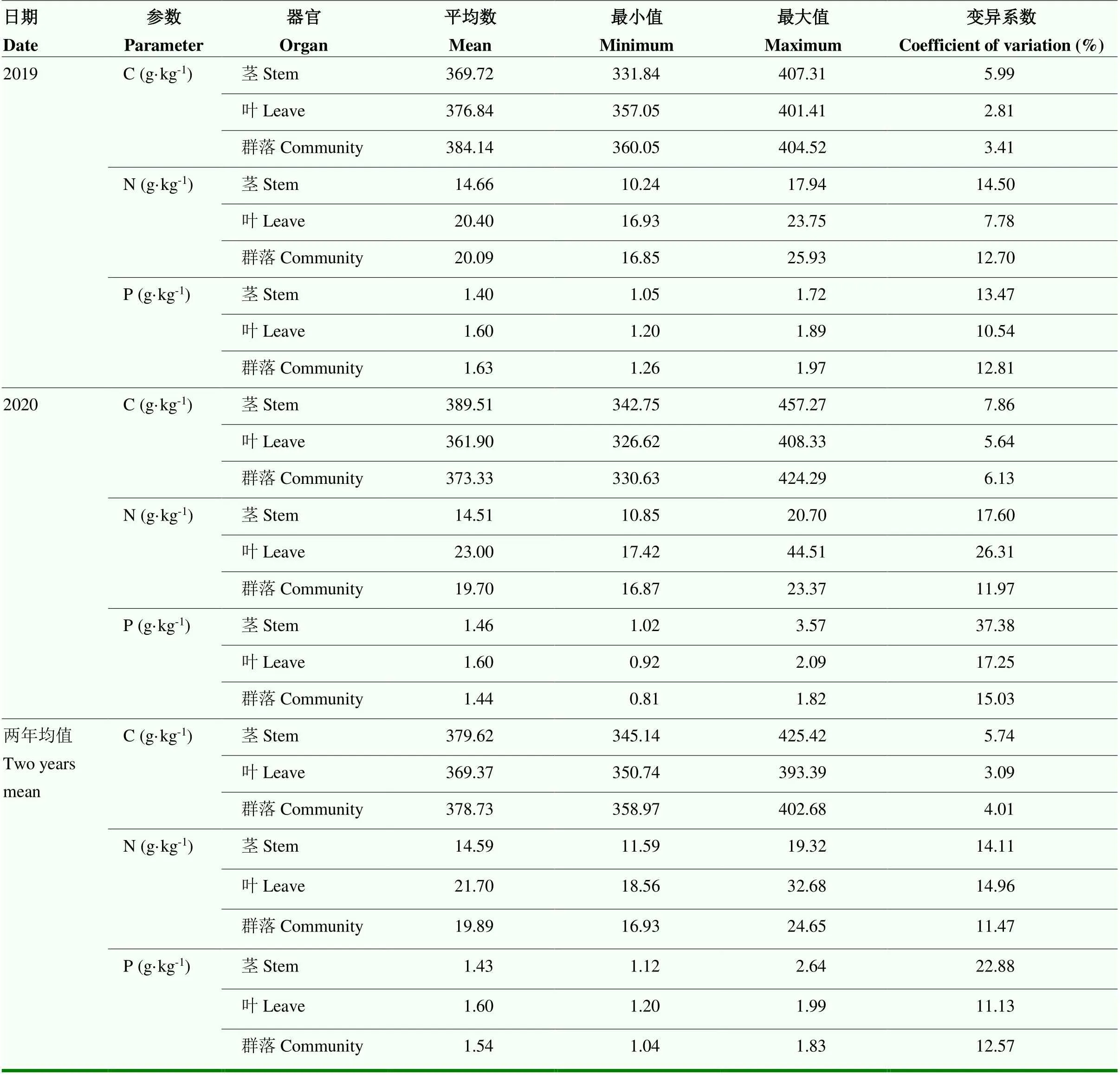
表2 不同放牧强度下不同器官C、N、P含量的变异特征
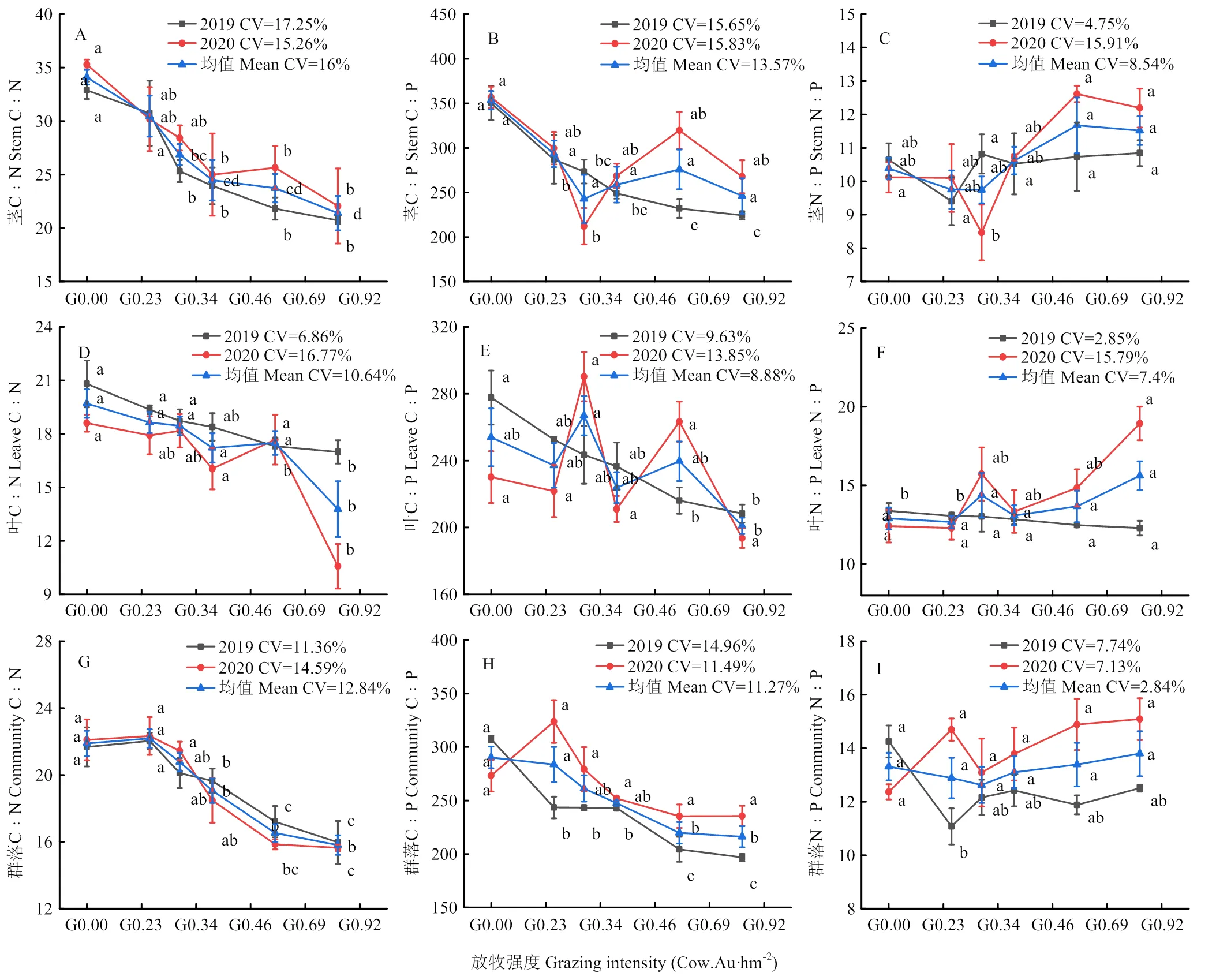
不同字母表示显著差异(P<0.05),相同字母表示差异不显著(P>0.05)。下同
2019年和2020年植物茎、叶、群落的氮含量以及茎叶比呈现出与碳含量相反的趋势(图4)。不同放牧强度下植物氮含量变化范围茎为11.78—17.83 g·kg-1,叶为18.44—33.94 g·kg-1,群落为18.19—22.97 g·kg-1,茎叶比为0.54—0.78。2019年(2=0.96,<0.001)和2020年(2=0.83,<0.05)植物茎氮含量、2019年叶氮含量(2=0.93,<0.05)均随着放牧强度的增加呈现显著的线性增加;2020年极重度放牧显著高于其他放牧强度(2=0.50,>0.05)。群落氮含量2019年(2=0.86,<0.05)与2020年(2=0.86,<0.05)均随着放牧强度的增加呈现显著的线性增加。氮含量的茎叶比(2=0.80,>0.05)随着放牧强度的增加呈现二项式变化,在重度放牧G0.69最高。
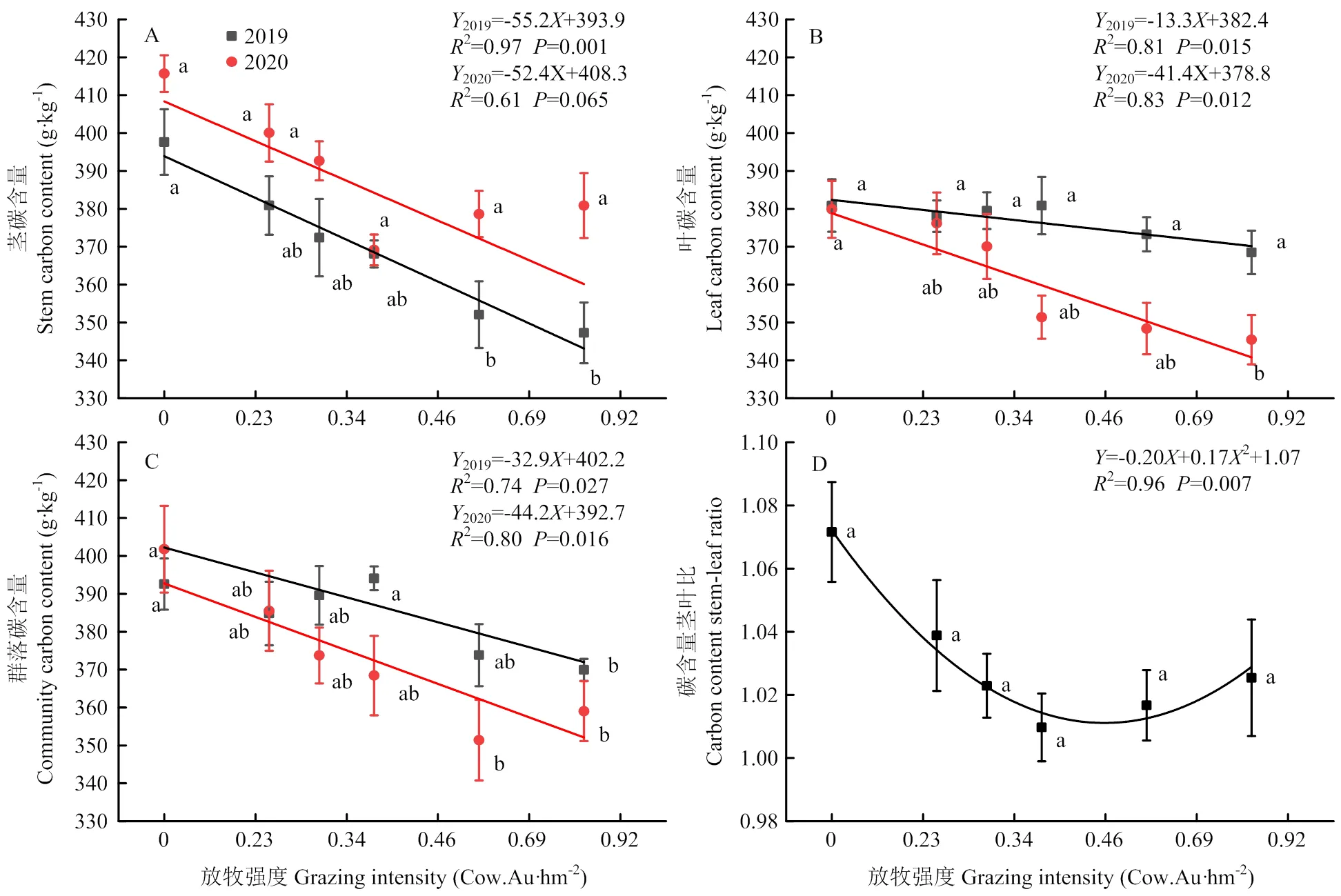
图3 不同放牧强度下茎、叶、群落碳含量及茎叶比
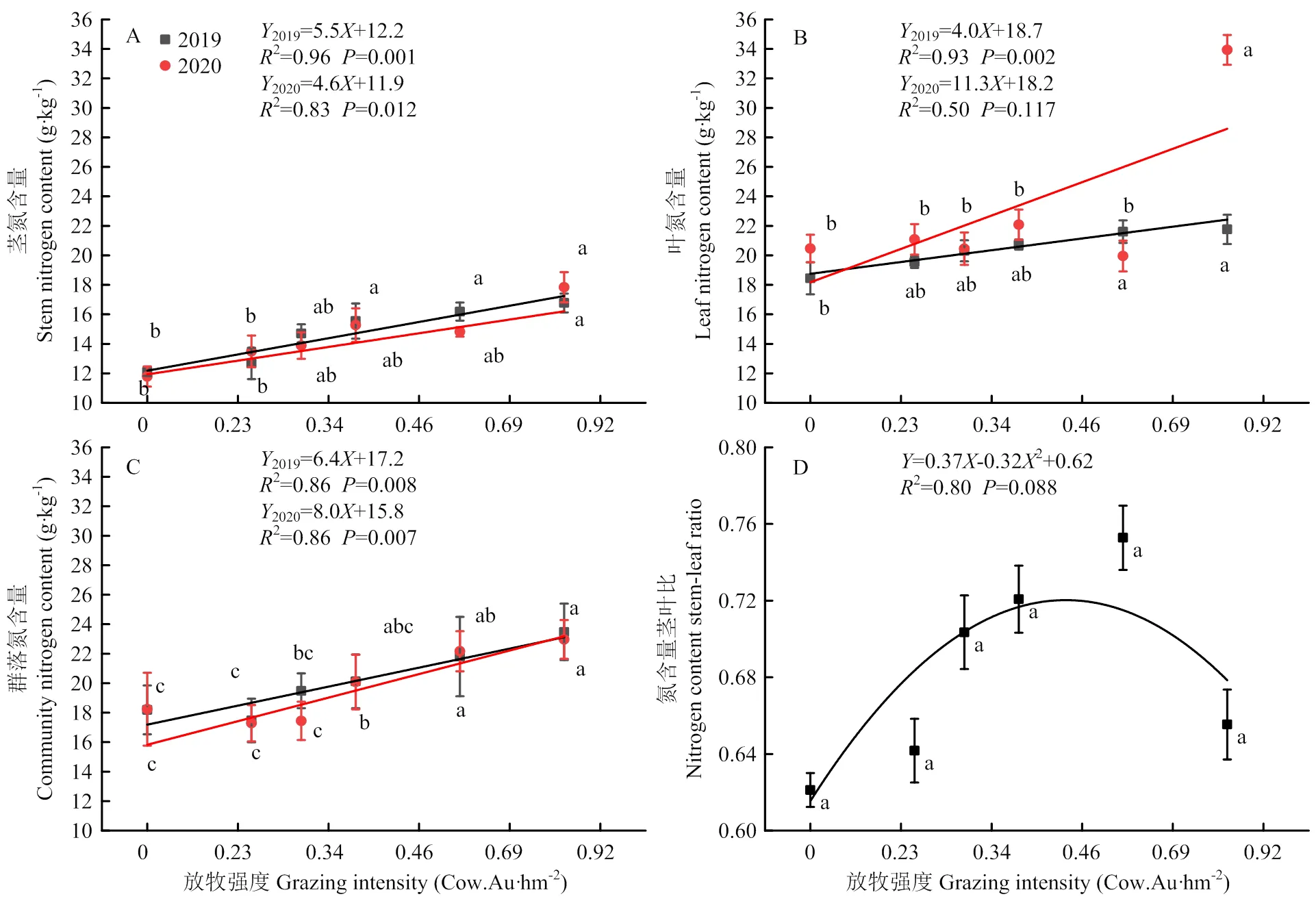
图4 不同放牧强度下茎、叶、群落氮含量及茎叶比
不同放牧强度下植物磷含量变化范围茎为1.14—2.05 g·kg-1,叶为1.39—1.77 g·kg-1,群落为1.18—1.88 g·kg-1,茎叶比为0.71—0.82(图5)。植物茎磷2019年含量G0.46、G0.69和G0.92显著高于不放牧(2=0.81,<0.05);植物叶磷含量2019年(2=0.89,<0.001)G0.69和G0.92显著高于不放牧(<0.05),2020年在各放牧强度间无显著差异(2=0.10,>0.05),峰值在G0.46处,达到1.76 g·kg-1。群落磷含量2019年(2=0.92,<0.05)与2020年(2=0.58>0.05)均随着放牧强度的增加呈现显著的线性下降。磷含量的茎叶比(2=0.23,>0.05)随着放牧强度的增加呈现二项式变化,G0.34处理最高。
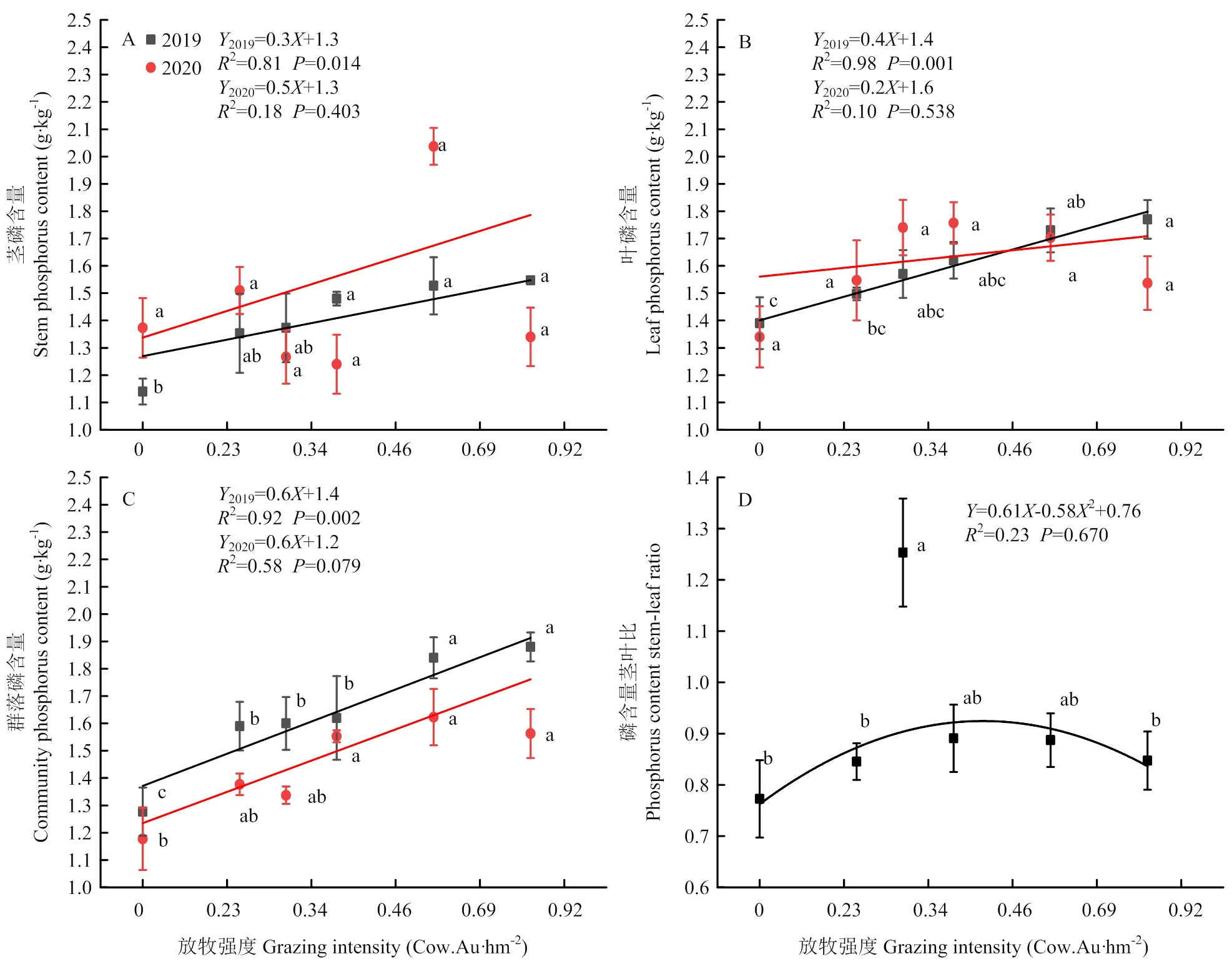
图5 不同放牧强度下茎、叶、群落磷含量及茎叶比
2.3 植物茎叶碳氮磷含量计量比与放牧强度和土壤理化性质相关性
通过对土壤理化性质与植物不同器官元素含量及计量比之间的相关性分析表明(表3):放牧强度与植物茎、叶、群落的碳、碳氮比以及茎和群落的氮磷比呈现出极显著负相关(<0.01),与茎、叶、群落的氮含量和群落的磷含量呈现极显著正相关(<0.01),与叶的磷含量和茎的氮磷比呈现显著正相关(<0.05)。温度与茎的磷含量呈现显著正相关(<0.05),水分与茎、叶、群落的碳氮比、茎和群落的碳磷比以及叶的碳含量呈现出极显著正相关,与茎、群落的氮含量和群落的磷含量呈现极显著负相关(<0.01),与叶的氮、磷含量呈现显著负相关,和群落的碳含量呈现显著正相关(<0.05)。土壤碳含量与植物叶片碳含量呈现极显著正相关,土壤氮含量与植物茎、叶、群落均呈现显著负相关。
2.4 植物群落碳氮磷总吸收量对放牧强度的响应
放牧降低了群落的总生物量、碳总吸收量和氮总吸收量,磷总吸收量呈现单峰曲线,G0.23时高于其他放牧强度(图6)。两年中,群落总生物量随着放牧强度的增加显著降低(<0.05),数值在102.76—235.83 g·m-2之间变化,重度G0.69和G0.92显著低于不放牧和轻度放牧(<0.05),与G0.00相比,G0.92两年群落总生物量分别降低了49.91%和26.66%;群落碳总吸收量与生物量变化趋势相同,重度G0.69和极重度G0.92放牧显著低于不放牧和轻度放牧(<0.05),两年中G0.92较G0.00分别降低了52.82%和34.51%;群落氮、磷总吸收量两年略有差异,2019年氮总吸收量随着放牧强度的增加显著降低(<0.05),中-重度放牧较不放牧降低了35.60%—41.46%,而群落磷总吸收量先增加再降低,两年均呈现轻度放牧G0.23时达到最大,2019年轻度放牧G0.23显著高于中-重度放牧(<0.05)。
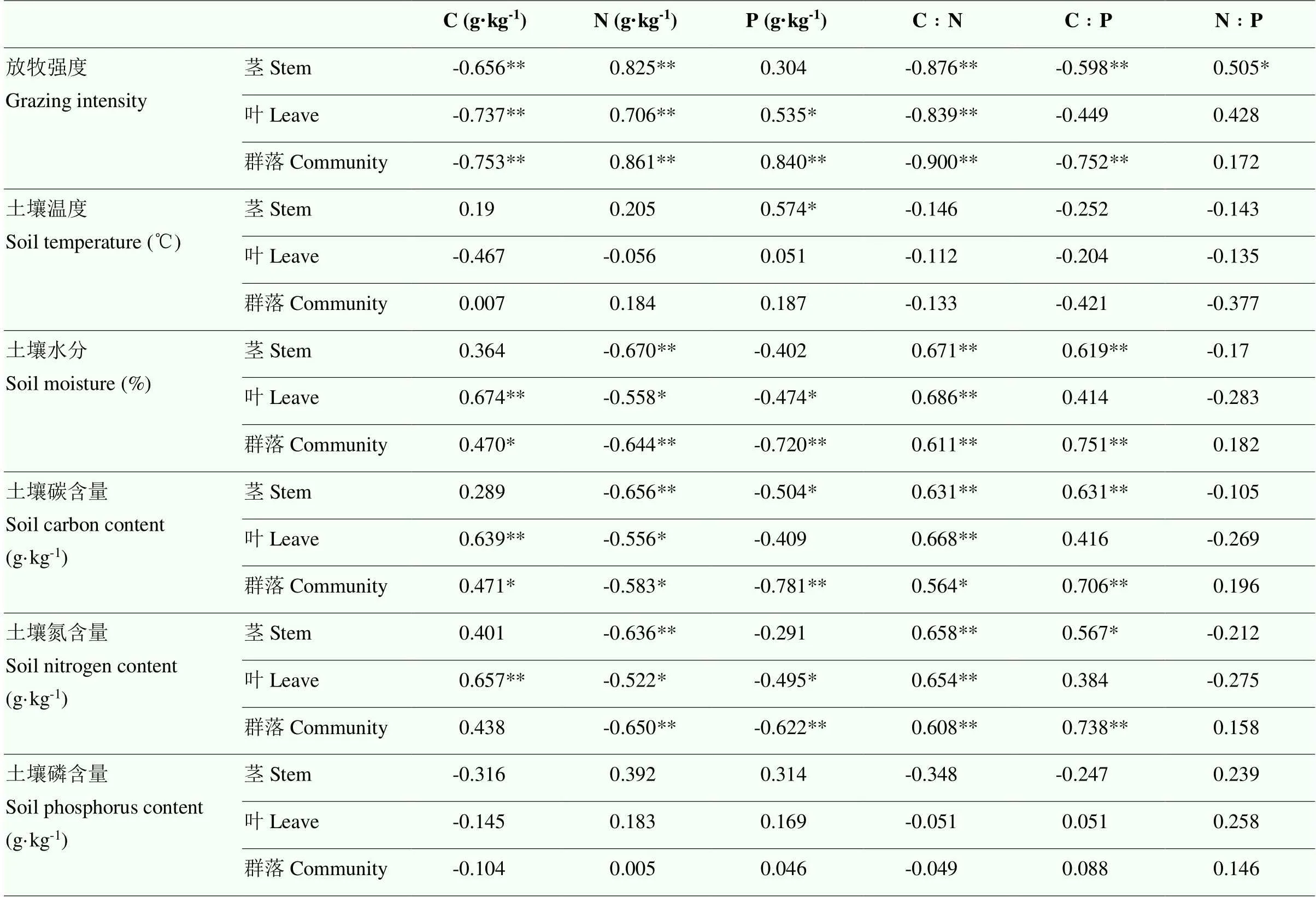
表3 土壤理化性质与植物不同器官元素含量及计量比之间的相关性分析
* 和 ** 分别表示在<0.05 和<0.01 水平下显著相关 *and ** indicate significant correlation at<0.05 and<0.01, respectively
3 讨论
3.1 放牧对植物器官及群落碳氮磷元素含量的影响
植物能主动调节对养分的需求,在外界的环境发生变化时,对自身的养分重新分配以抵御逆境,所以植物各组织器官的碳、氮、磷元素含量随外界环境不断改变[14]。本研究中,不同放牧强度下植物茎、叶、群落的碳含量均低于呼伦贝尔草地叶片碳含量(448.2 g·kg-1)和全球尺度上陆生植物叶片碳含量(461.6 g·kg-1)[15],说明在各放牧强度下,植物的碳同化能力均较低。除极重度放牧条件下的叶,其余不同放牧强度下茎叶、群落的氮含量均低于青藏高原草地(24.86 g·kg-1)和内蒙古草原(26.8 g·kg-1)[16],各放牧强度下茎、叶、群落的磷含量均高于韩文轩[17](2.0 g·kg-1)和任书杰[18](1.56 g·kg-1)的研究结果,这是由于放牧活动所导致的践踏与采食,消除老叶及其残叶的同时促进了牧草的生长,刺激氮元素向幼嫩部位的重新分配,提高植物叶氮含量。
本研究中,放牧显著降低了植物茎、叶、群落的碳含量,这与范月君等[19]、许雪贇等[3]的研究结果一致,但丁小慧等[5]和鄢燕等[20]认为放牧对植物叶片及群落的碳含量无显著影响,LI等[21]认为放牧使叶片和茎器官中的碳含量显著升高。放牧牲畜的种类和放牧强度的设置不同是造成植物碳含量变化不一致的主要原因,而碳含量降低的主要原因是随放牧强度的增加,动物的采食和践踏导致衰老部位存在的比重逐渐下降,再生幼叶所占比例相对较多,幼叶不能进行固碳,导致整体碳含量降低[22]。土壤碳含量与植物叶片、群落有显著正相关关系,表明其碳含量主要来源是土壤,而非光合作用,这也与前文幼叶无法固碳导致叶碳含量逐渐降低一致。研究结果表明,放牧会增加茎、叶、群落的氮、磷含量,不同放牧强度下茎的磷含量变异系数最大,这一结果的原因是家畜的采食和践踏,消除衰老部位,促进了幼嫩组织的光合作用,进而提高了叶片中的叶绿素含量,氮元素是叶绿素的主要组成成分,这是植物补偿生长的一种表现[23],而植物体中的氮、磷是协同元素,一般呈正相关关系[7],即放牧会使得茎、叶、群落的氮、磷含量均上升。这与仲延凯等[24]在内蒙古典型草原研究所得,割草促进草地植物氮含量增加以及张婷等[9]、BAi等[25]研究放牧会显著增加植物茎、叶中氮、磷含量结果类似。然而,与这些结果相反的是,有研究认为[26-27]放牧会减少地上部分氮素分配,即放牧使得茎、叶中氮含量呈降低趋势。这种结果是植物与土壤相互作用有关的养分利用策略的复杂机制造成的,重度放牧动物的采食和践踏会导致土壤有机氮的流失,植物吸收氮元素的量随之降低,但本研究结果中,土壤氮含量与植物氮含量呈现显著负相关,这是因为夏季为植物的快速生长季,随着放牧强度的增加,植物为抵抗胁迫增加对氮元素的吸收,土壤氮含量却随着养分流失而降低,从而形成了植物氮含量与土壤氮含量成反比的关系,这与上述研究结果不同。本试验样地进行了连续11年的放牧处理,在草地生态系统中土壤特性的响应具有滞后作用[28]。因此,本研究中,茎、叶、群落的碳含量随着放牧强度的增加而降低,氮、磷元素含量增加的现象表明,适度放牧可以提高植物碳氮磷元素的循环速率和可利用性,因此要合理控制草原的放牧强度。

图6 不同放牧强度下群落生物量及碳氮磷总吸收量
3.2 放牧对植物器官及群落化学计量学的影响
植物叶片碳氮比和碳磷比意味着植物吸收氮、磷元素时所能同化碳的能力,反映了植物的养分利用效率和生长速率[29]。白永飞等[30]、范月君等[19]、丁莉君等[31]的研究结果表明,随着放牧强度的增加,植物碳氮比降低。有观点认为放牧通过家畜采食减少了植物部分衰老组织,幼嫩组织生长速率较快,使叶片碳氮比降低[23],且李香真等[10]的结果表明,叶的碳氮比的变化趋势与全株的相类似。这与本试验结果相同:随着放牧强度的增加,茎、叶、群落的碳氮比均降低。分析原因是载畜率高时,家畜的排泄物提供的速效氮增多,满足植物对氮素的需求,由于植物碳氮比较低时,植物生长速率较快,对氮元素的需求量增大,使得养分周转的周期变短,最终导致放牧强度越高,碳氮比越低[9]。本研究结果显示,茎与群落的碳磷比变化均随着放牧强度的增加而降低,这与LI等[21]的研究结果一致:碳磷比与放牧压力呈显著负相关,主要是由于随放牧强度的增加,动物的采食践踏导致再生幼嫩器官所占比例相对较多,但其碳含量较低,磷含量较高,进而导致碳磷比降低,这是植物为了提高对逆境的适应能力。不同放牧强度下只有茎在不放牧、轻度及中度放牧时的碳氮比和不放牧时的碳磷比高于全球水平(23.80、300.90)[32],说明牲畜的采食会减弱植物固碳潜能和同化碳的能力。长期以来,在植物与草食动物的相互作用系统中限制草原植物的养分一直是一个有争议的话题[33],而氮磷比可作为判断环境对植物生长养分供应状况和植物生长速率的重要指标[34]。Koerselman等[35]的试验研究结果表明,当植物N﹕P<14时,表现为受氮的限制;当N﹕P>16时,表现为受磷的限制,介于两者中间表明受到氮、磷元素的共同限制作用。He等[36]研究指出叶片作为植物的主要光合器官,其氮磷比的限制关系会随着外界环境的改变而改变。本研究中,叶的氮磷比最大,其次是群落,茎的氮磷比最低,叶在轻度放牧G0.34和极重度放牧G0.92时氮磷比介于14与16之间,受到氮、磷元素的共同限制,其余在不同放牧强度下均低于14,表现为受到氮的限制。仔细研究后发现,是家畜采食刺激氮、磷向地上幼嫩器官重新分配,氮的转移量大于磷,而叶作为最主要的采食对象,再生长能力更强[2]。综上所述,中度放牧时植物既可以保持较高的生长速率,又可以高效利用养分。
3.3 放牧对植物群落碳氮磷总吸收量的影响
董晓玉等[37-38]的研究表明,草地植物元素总含量由元素含量和地上生物量共同决定,植物碳、氮、磷元素总含量均与生物量呈显著正相关关系。但碳、氮、磷的总含量受元素含量影响不大,这些变化都与地上生物量的变化密切相关。放牧对草地的影响首先体现在地上生物量[39-40],家畜觅食过程中适口性较好的物种优先被采食,会打破原有草地的平衡,随着放牧强度的增加,地上生物量逐渐降低[41]。本研究结果与之相同,碳、氮、磷总含量与群落生物量的变化趋势相似,均随着放牧强度的增加而降低。因此,如若想获得较大的碳、氮、磷含量则需要控制放牧强度,适度放牧来提高地上部总生物量,进而提高元素总含量。
4 结论
放牧使得植物碳、氮、磷含量及其生态化学计量比发生了明显的变化,降低了植物茎、叶、群落的碳含量、碳氮比以及碳磷比;增加了植物茎、叶、群落的氮磷含量;植物各器官中,仅叶在轻度放牧和极重度放牧时受到氮、磷元素的共同限制,其余器官在不同放牧强度下均受到氮的限制。
放牧强度与植物茎、叶、群落的碳含量呈现极显著负相关,与植物茎、叶、群落的氮含量和叶、群落的磷含量呈现显著正相关。土壤水分与植物叶、群落碳含量呈显著正相关;与茎、叶、群落的氮含量和叶、群落的磷含量呈现显著负相关。土壤碳含量与植物叶片碳含量呈现极显著正相关,土壤氮含量与植物茎、叶、群落氮含量均呈现显著负相关。
重度放牧显著降低了群落的总生物量及对碳氮磷元素的总吸收量,适度放牧G0.34有利于维持并促进草地生态功能可持续稳定发展。
[1] ELDRIDGE D J, DELGADO-BAQUERIZO M, TRAVERS S K, VAL J, OLIVER I, HAMONTS K, SINGH B K. Competition drives the response of soil microbial diversity to increased grazing by vertebrate herbivores. Ecology, 2017, 98(7): 1922-1931.
[2] 牛得草, 董晓玉, 傅华. 长芒草不同季节碳氮磷生态化学计量特征. 草业科学, 2011, 28(6): 915-920.
NIU D C, DONG X Y, FU H. Seasonal dynamics of carbon, nitrogen and phosphorus stoichiometry in. Pratacultural Science, 2011, 28(6): 915-920. (in Chinese)
[3] 许雪贇, 曹建军, 杨淋, 杨书荣, 龚毅帆, 李梦天. 放牧与围封对青藏高原草地土壤和植物叶片化学计量学特征的影响. 生态学杂志, 2018, 37(5): 1349-1355.
XU X Y, CAO J J, YANG L, YANG S R, GONG Y F, LI M T. Effects of grazing and enclosure on foliar and soil stoichiometry of grassland on the Qinghai-Tibetan Plateau. Chinese Journal of Ecology, 2018, 37(5): 1349-1355. (in Chinese)
[4] 郑伟. 封育对伊犁绢蒿种群营养元素分配格局及贮量的影响. 草业科学, 2012, 29(10): 1503-1511.
ZHENG W. Effects of enclosure on storage and allocation of carbon and nutrient elements inpopulation. Pratacultural Science, 2012, 29(10): 1503-1511. (in Chinese)
[5] 丁小慧, 宫立, 王东波, 伍星, 刘国华. 放牧对呼伦贝尔草地植物和土壤生态化学计量学特征的影响. 生态学报, 2012, 32(15): 4722-4730.
DING X H, GONG L, WANG D B, WU X, LIU G H. Grazing effects on eco-stoichiometry of plant and soil in Hulunbeir, Inner Mogolia. Acta Ecologica Sinica, 2012, 32(15): 4722-4730. (in Chinese)
[6] FRANK D A. Ungulate and topographic control of nitrogen: phosphorus stoichiometry in a temperate grassland; soils, plants and mineralization rates. Oikos, 2008, 117(4): 591-601.
[7] HAN W X, FANG J Y, GUO D L, ZHANG Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377-385.
[8] 李香真, 陈佐忠. 不同放牧率对草原植物与土壤C、N、P含量的影响. 草地学报, 1998, 6(2): 90-98.
LI X Z, CHEN Z Z. Influences of stocking rates on C, N, P contents in plant-soil system. Acta Agrestia Sinica, 1998, 6(2): 90-98. (in Chinese)
[9] 张婷, 翁月, 姚凤娇, 史印涛, 崔国文, 胡国富. 放牧强度对草甸植物小叶章及土壤化学计量比的影响. 草业学报, 2014, 23(2): 20-28.
ZHANG T, WENG Y E, YAO F J, SHI Y T, CUI G W, HU G F. Effect of grazing intensity on ecological stoichiometry ofand meadow soil. Acta Prataculturae Sinica, 2014, 23(2): 20-28. (in Chinese)
[10] 李香真, 邢雪荣, 陈佐忠. 不同放牧率对旱黄梅衣生物量和化学元素组成的影响. 应用生态学报, 2001, 12(3): 369-373.
LI X Z, XING X R, CHEN Z Z. Effect of grazing rate on biomass and element composition of. Chinese Journal of Applied Ecology, 2001, 12(3): 369-373. (in Chinese)
[11] BARDGETT R D, WARDLE D A. Herbivore-mediated linkages between aboveground and belowground communities. Ecology, 2003, 84(9): 2258-2268.
[12] 张宇, 侯路路, 闫瑞瑞, 辛晓平. 放牧强度对草甸草原植物群落特征及营养品质的影响. 中国农业科学, 2020, 53(13): 2550-2561.
ZHANG Y, HOU L L, YAN R R, XIN X P. Effects of grazing intensity on plant community characteristics and nutrient quality of herbage in a meadow steppe. Scientia Agricultura Sinica, 2020, 53(13): 2550-2561. (in Chinese)
[13] 鲍士旦. 土壤农化分析. 3版. 北京: 中国农业出版社, 2000.
BAO S D. Soil and Agricultural Chemistry Analysis.3rd ed. Beijing: Chinese Agriculture Press, 2000. (in Chinese)
[14] 李红林, 贡璐, 洪毅. 克里雅绿洲旱生芦苇根茎叶C、N、P化学计量特征的季节变化. 生态学报, 2016, 36(20): 6547-6555.
LI H L, GONG L, HONG Y. Seasonal variations in C, N, and P stoichiometry of roots, stems, and leaves ofin the Keriya Oasis, Xinjiang, China. Acta Ecologica Sinica, 2016, 36(20): 6547-6555. (in Chinese)
[15] 贺合亮, 阳小成, 李丹丹, 尹春英, 黎云祥, 周国英, 张林, 刘庆. 青藏高原东部窄叶鲜卑花碳、氮、磷化学计量特征. 植物生态学报, 2017(1): 126-135.
HE H L, YANG X C, LI D D, YIN C Y, LI Y X, ZHOU G Y, ZHANG L, LIU Q. Stoichiometric characteristics of carbon, nitrogen and phosphorus ofshrub on the eastern Qinghai- Xizang Plateau. Chinese Journal of Plant Ecology, 2017(1): 126-135. (in Chinese)
[16] HE J S, FANG J Y, WANG Z H, GUO D L, FLYNN D F B, GENG Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia, 2006, 149(1): 115-122.
[17] 韩文轩, 吴漪, 汤璐瑛, 陈雅涵, 李利平, 贺金生, 方精云. 北京及周边地区植物叶的碳氮磷元素计量特征. 北京大学学报(自然科学版), 2009, 45(5): 855-860.
HAN W X, WU Y, TANG L Y, CHEN Y H, LI L P, HE J S, FANG J Y. Leaf carbon, nitrogen and phosphorus stoichiometry across plant species in Beijing and its periphery. Acta Scientiarum Naturalium Universitatis Pekinensis, 2009, 45(5): 855-860. (in Chinese)
[18] 任书杰, 于贵瑞, 陶波, 王绍强. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究. 环境科学, 2007, 28(12): 2665-2673.
REN S J, YU G R, TAO B, WANG S Q. Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC. Environmental Science, 2007, 28(12): 2665-2673. (in Chinese)
[19] 范月君, 侯向阳, 石红霄, 师尚礼. 封育与放牧对三江源区高寒草甸植物和土壤碳储量的影响. 草原与草坪, 2012, 32(5): 41-46, 52.
FAN Y J, HOU X Y, SHI H X, SHI S L. The response of carbon reserves of plants and soils to different grassland managements on alpine meadow of three headwater source regions. Grassland and Turf, 2012, 32(5): 41-46, 52. (in Chinese)
[20] 鄢燕, 马星星, 鲁旭阳. 人为干扰对藏北高寒草原群落生物量及其碳氮磷含量特征的影响. 山地学报, 2014, 32(4): 460-466.
YAN Y, MA X X, LU X Y. Effect of human disturbance on plant biomass and CNP contents of the alpine steppe in northern Tibet. Mountain Research, 2014, 32(4): 460-466. (in Chinese)
[21] LI X L, LIU Z Y, REN W B, DING Y, JI L, GUO F H, HOU X Y. Linking nutrient strategies with plant size along a grazing gradient: Evidence fromin a natural pasture. Journal of Integrative Agriculture, 2016, 15(5): 1132-1144.
[22] 孙宗玖, 李培英. 放牧对伊犁绢蒿叶片碳氮磷化学计量特征的影响. 中国草学会第九次全国会员代表大会暨学术讨论会论文集. 2016: 7.
SUN Z J, LI P Y. Effects of grazing on carbon, nitrogen and phosphorus stoichiometry in leaves of Seriphidium iliense//The Ninth National Member Congress of China Grass Society and Academic Symposium Proceedings. China Grass Society, 2016: 7. (in Chinese)
[23] 翟夏杰, 黄顶, 王堃. 围封与放牧对典型草原植被和土壤的影响. 中国草地学报, 2015, 37(6): 73-78.
ZHAI X J, HUANG D, WANG K. Effects of fencing and grazing on vegetation and soil in typical grassland. Chinese Journal of Grassland, 2015, 37(6): 73-78. (in Chinese)
[24] 仲延凯, 孙维, 孙卫国, 包青海. 割草对典型草原植物营养元素贮量及分配的影响Ⅳ.土壤—植物营养元素含量的比较. 干旱区资源与环境, 2000, 14(1): 55-63.
ZHONG Y K, SUN W, SUN W G, BAO Q H. Influence of mowing on storing amount and distribution of nutrient elements in typical steppe ⅣA comparative study on nutrient element contents of soil and plant. Journal of Arid Land Resources and Environment, 2000, 14(1): 55-63. (in Chinese)
[25] BAI Y F, WU J G, CLARK C M, PAN Q M, ZHANG L X, CHEN S P, WANG Q B, HAN X G. Grazing alters ecosystem functioning and C: N:P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology, 2012, 49(6): 1204-1215.
[26] 宋智芳, 安沙舟, 孙宗玖. 放牧地伊犁绢蒿营养元素分配特点. 草业科学, 2014, 31(1): 132-138.
SONG Z F, AN S Z, SUN Z J. Characteristics of nutrition allocation ofunder grazing condition. Pratacultural Science, 2014, 31(1): 132-138. (in Chinese)
[27] CUI X Y, WANG Y F, NIU H S, WU J, WANG S P, SCHNUG E, ROGASIK J, FLECKENSTEIN J, TANG Y H. Effect of long-term grazing on soil organic carbon content in semiarid steppes in Inner Mongolia. Ecological Research, 2005, 20(5): 519-527.
[28] JIA B R, ZHOU G S, WANG F Y, WANG Y H, WENG E S. Effects of grazing on soil respiration ofsteppe. Climatic Change, 2007, 82(1): 211-223.
[29] 黄小波, 刘万德, 苏建荣, 李帅锋, 郎学东. 云南普洱季风常绿阔叶林152种木本植物叶片C、N、P化学计量特征. 生态学杂志, 2016, 35(3): 567-575.
HUANG X B, LIU W D, SU J R, LI S F, LANG X D. Stoichiometry of leaf C, N and P across 152 woody species of a monsoon broad-leaved evergreen forest in Pu'er, Yunnan Province. Chinese Journal of Ecology, 2016, 35(3): 567-575. (in Chinese)
[30] 白永飞, 黄建辉, 郑淑霞, 潘庆民, 张丽霞, 周华坤, 徐海量, 李玉霖, 马健. 草地和荒漠生态系统服务功能的形成与调控机制. 植物生态学报, 2014, 38(2): 93-102.
BAI Y F, HUANG J H, ZHENG S X, PAN Q M, ZHANG L X, ZHOU H K, XU H L, LI Y L, MA J. Drivers and regulating mechanisms of grassland and desert ecosystem services. Chinese Journal of Plant Ecology, 2014, 38(2): 93-102. (in Chinese)
[31] 丁莉君, 卫智军, 张爽, 刘佳, 吕世杰. 放牧强度季节调控对荒漠草原土壤和植物养分的影响. 草原与草业, 2019, 31(1): 53-59.
DING L J, WEI Z J, ZHANG S, LIU J, LÜ S J. Effects of seasonal regulation of grazing intensity on soil and plant nutrients in desert steppe. Grassland and Prataculture, 2019, 31(1): 53-59. (in Chinese)
[32] 李从娟, 雷加强, 徐新文, 唐清亮, 高培, 王永东. 塔克拉玛干沙漠腹地人工植被及土壤CNP的化学计量特征. 生态学报, 2013, 33(18): 5760-5767.
LI C J, LEI J Q, XU X W, TANG Q L, GAO P, WANG Y D. The stoichiometric characteristics of C, N, P for artificial plants and soil in the hinterland of Taklimakan Desert. Acta Ecologica Sinica, 2013, 33(18): 5760-5767. (in Chinese)
[33] MAMOLOS A P, VASILIKOS C V, VERESOGLOU D S. Vegetation in contrasting soil water sites of upland herbaceous grasslands and N: P ratios as indicators of nutrient limitation. Plant and Soil, 2005, 270(1): 355-369.
[34] 吴统贵, 吴明, 刘丽, 萧江华. 杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化. 植物生态学报, 2010, 34(1): 23-28.
WU T G, WU M, LIU L, XIAO J H. Seasonal variations of leaf nitrogen and phosphorus stoichiometry of three herbaceous species in Hangzhou Bay coastal wetlands, China. Chinese Journal of Plant Ecology, 2010, 34(1): 23-28. (in Chinese)
[35] KOERSELMAN W, MEULEMAN A F M. The vegetation N: P ratio: A new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology, 1996, 33(6): 1441.
[36] HE J S, WANG L A, FLYNN D F B, WANG X P, MA W H, FANG J Y. Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes. Oecologia, 2008, 155(2): 301-310.
[37] 董晓玉. 放牧与围封对黄土高原典型草原植物碳、氮、磷生态计量特征及其贮量的影响[D]. 兰州: 兰州大学, 2009.
DONG X Y. The effect of grazing and fencing on ecological stoichiometry of carbon, nitrogen, phosphorus and their storages in steppe grasslands of loess plateau[D]. Lanzhou: Lanzhou University, 2009. (in Chinese)
[38] 董晓玉, 傅华, 李旭东, 牛得草, 郭丁, 李晓东. 放牧与围封对黄土高原典型草原植物生物量及其碳氮磷贮量的影响. 草业学报, 2010, 19(2): 175-182.
DONG X Y, FU H, LI X D, NIU D C, GUO D, LI X D. Effects on plant biomass and CNP contents of plants in grazed and fenced steppe grasslands of the Loess Plateau. Acta Prataculturae Sinica, 2010, 19(2): 175-182. (in Chinese)
[39] ELSER J J, STERNER R W, GOROKHOVA E, FAGAN W F, MARKOW T A, COTNER J B, HARRISON J F, HOBBIE S E, ODELL G M, WEIDER L W. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540-550.
[40] HEDIN L O. Global organization of terrestrial plant-nutrient interactions. PNAS, 2004, 101(30): 10849-10850.
[41] 侯路路, 闫瑞瑞, 张宇, 辛晓平. 放牧强度对草甸草原羊草功能性状的影响. 中国农业科学, 2020, 53(13): 2562-2572.
HOU L L, YAN R R, ZHANG Y, XIN X P. Effects of grazing intensity on functional traits ofin meadow steppe. Scientia Agricultura Sinica, 2020, 53(13): 2562-2572. (in Chinese)
Effects of Grazing Disturbance on the Stoichiometry of Nitrogen and Phosphorus in Plant Organs ofMeadow Steppe
WANG Miao1, ZHANG Yu1, LI RuiQiang2, XIN XiaoPing1, ZHU XiaoYu3, CAO Juan1, ZHOU ZhongYi4, YAN RuiRui1
1Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences/National Field Scientific Observation and Research Station of Hulunbuir Grassland Ecosystem, Beijing 100081;2Inner Mongolia Autonomous Region Environmental Online Monitoring Centre, Hohhot 010055;3Environmental Protection Research and Monitoring Institute, Ministry of Agriculture and Rural Affairs, Tianjin 300191;4Career Development Center of Yakeshi Forestry and Grassland Administration, Yakeshi 022150, Inner Mongolia
【】Taking above-ground communities, stems and leaves as research objects, the effects of different grazing intensities on the contents of carbon (C), nitrogen (N) and phosphorus (P) elements and stoichiometric characteristics of plant components and plant communities were investigated, which provided a scientific basis for in-depth analysis of grassland ecosystem degradation process under grazing disturbance and promotion of adaptive management of grassland grazing.【】Based on the long-term controlled grazing test platform ofmeadow grassland in Hulunbuir, the plant communities and plant samples of different components were collected. The contents of C, N and P were determined by elemental analysis method, and then the changes of C, N and P contents and the measurement ratio under different grazing intensities were analyzed.【】With the increase of grazing intensity, the C content of plant stem, leaf and community showed a significant linear decrease, and the C content of heavy to extremely heavy grazing was significantly lower than that of no grazing (<0.05). The N and P contents of the community were linearly increased, and the heavy to extremely grazing was significantly higher than that of no grazing (<0.05). The stem-leaf ratio of C content was the lowest in moderate grazing (G0.46), while the stem-leaf ratio of N content was the highest in heavy grazing (G0.69), and the highest in P content (G0.34). The coefficients of variation of C content and N/P ratio in different plant organs were small, but the coefficients of variation of N and P contents were large. The variation coefficients of C content and N/P ratio of different components ranged from 4.01% to 5.74% and 2.84% to 8.54%, respectively. The variation coefficients of N and P contents of plants ranged from 11.47% to 14.96% and 11.13% to 22.88%, respectively, and the variation coefficients of C/N and C/P ratios ranged from 10.64% to 16.00% and 8.88% to 13.57%, respectively; among them, the coefficient of variation of the N/P ratio of plant leaves is the smallest, and the coefficient of variation of plant stem P content is the largest. The N/P ratio of leaves was between 14 and 16 under light grazing G0.34 and extremely heavy grazing G0.92, which was restricted by both N and P elements; while the other components were all lower than 14 under different grazing intensities, which was restricted by N. There was a significant negative correlation between grazing intensity and C content and C/N ratio of stem, leaf and community, and N/P ratio of stem and community, whilst they were significantly positively related to N content of stem, leaf and community, P content of plant leaf and community, and the N/P ratio of plant stem. The soil moisture was significantly positively correlated with the C content of plant leaves and community, the C/N ratio of stem, leaf and community, and the C/P ratio of stem and community, however, which were significantly negatively correlated the N content of stem, leaf and community and the P content of leaf and community.【】Grazing caused significant changes in plant C, N, P content, ecological stoichiometric ratio, and total community C, N, and P content. Overgrazing not only decreased the C content, C/N ratio and C/P ratio of plant stem, leaf and community, but also reduced the total C, N, and P content of the community, increased the N and P content of plant stem, leaf and community, and N/P ratio of plant stem, leaf and community. Moderate grazing decreased the C/N ratio of stem and leaf, and increased the C/P ratio and N/P ratio of stem and leaf, which was beneficial to promote the sustainable and stable development of grassland ecological functions.
meadow steppe; grazing intensity; carbon, nitrogen and phosphorus content; ecological stoichiometry characteristics; stem and leaf; community; Hulunbuir

10.3864/j.issn.0578-1752.2022.07.009
2021-03-01;
2021-07-22
国家自然科学基金面上项目(31971769)、国家重点研发计划(2016YFC0500601,2017YFE0104500)、中央级公益性科研院所基本科研业务费专项(Y2019YJ13,Y2020YJ19,1610132021016)、呼伦贝尔市科技计划项目(YYYFHZ201903)、财政部和农业农村部:国家现代农业产业技术体系、农业科技创新联盟建设-农业基础性长期性科技工作(NAES037SQ18)
王淼,E-mail:wangmiao2021163@163.com。通信作者闫瑞瑞,E-mail:yanruirui@caas.cn
(责任编辑 李云霞)
