不同天然基质对条斑紫菜丝状体培养的影响*
2022-04-12尚浩江胡传明朱朋昌
尚浩江,周 伟,胡传明**,申 欣,朱朋昌,李 帅
(1.江苏海洋大学,江苏省海洋生物技术重点实验室,江苏连云港 222000;2.江苏省海洋水产研究所,江苏南通 226007;3.上海海洋大学,上海 201306)
条斑紫菜(Neopyropiayezoensis)是我国重要的经济红藻,主产于江苏海区,此外山东与辽宁海区也有少量分布。2020-2021生产年度,条斑紫菜育苗面积达8×105m2,海上栽培面积4.1×104hm2,干紫菜产量5.6×109枚(数据由江苏省紫菜协会提供),条斑紫菜产业是我国长江以北人工栽培海藻中最具经济价值的产业[1]。长期以来条斑紫菜种苗培育效率滞后于栽培面积的快速发展,每年均会出现部分地区种苗供应紧张的情况,导致种苗培育面积无序发展,育苗面积与栽培面积比逐年增加,育苗企业亏损面扩大,不利于紫菜产业的稳定。紫菜的种苗培育技术急需向机械化、智能化方向发展以提高育苗效率,而其中种苗培养基质的筛选,以及突破贝壳基质的制约因素而开发新型基质,是解决工厂化育苗的关键环节。
1953-1954年,日本藻类学家黑木宗尚和我国藻类学家曾呈奎几乎同时完成了甘紫菜(Neopyropiatenera,曾用名Porphyratenera)生活史的研究,为紫菜的人工育苗打下理论基础[2],此后曾有自由丝状体(Free-living conchocelis)采苗[3]、单孢子采苗[4]、紫菜叶状体细胞酶法采苗[5]等无基质育苗技术出现,均因采苗量达不到生产要求而未被市场接受。目前紫菜种苗产业仍采用基质培养技术,且贝壳是唯一的培养基质,天然贝壳由95%-99%的碳酸钙(方解石、文石、球霰石及非晶态)和1%-5%的蛋白质(糖蛋白、多糖、几丁质和脂质等)构成[6],种苗以贝壳丝状体(Shell conchocelis[7])形式生长于贝壳内层。这与贝壳丝状体的特殊生长方式有关,紫菜果孢子附着于贝壳内层表面,萌发形成丝状体后钻入贝壳生长[8],而硅藻、蓝藻等杂藻不能钻入贝壳,因此生产培养中可通过简单的清洗贝壳表面加以纯化,从而可以大规模培养贝壳丝状体。相反,悬浮培养的自由丝状体常因杂藻而导致污染难以去除[9]。此外,贝壳丝状体生长于贝壳内层表面一定深度,受光较为均匀,可以大量同步形成“双分”,保障了短期内大规模采苗的要求。
文蛤(Meretrixmeretrix)、牡蛎、三角帆蚌(Hyriopsiscumingii)和扇贝的贝壳是4种广泛应用的紫菜种苗培养基质,适宜紫菜丝状体生长且较易获取[10]。Heo等[11]研究认为牡蛎壳的紫菜丝状体接种效率高于扇贝壳和文蛤壳。除贝壳外,也有研究表明蛋壳可作为紫菜种苗培养的基质[12]。但目前对紫菜种苗培养技术研究主要聚焦于培养的环境因子上,如光照强度、光周期与培养水温等[13-15],以及黄斑病[16,17]等病害防治方面,未见对贝壳培育性能的系统性研究。因此,本研究选取不同基质材料,如文蛤、近江牡蛎(Ostrearivularis)壳、三角帆蚌壳、虾夷扇贝壳(Patinopectenyessoensis)壳、海月蛤(Placunaplacenta)壳、鸡蛋壳、珍珠等生物合成基质,以及方解石原石、霰石原石、几丁质碎片与壳聚糖薄膜等与贝壳成分相关的非生物基质,拟通过比较分析,研究多种基质对条斑紫菜贝壳丝状体着生、生长与壳孢子放散的影响,筛选最适培养基质,为贝壳替代基质的研究打下科学基础。
1 材料与方法
1.1 材料
条斑紫菜自由丝状体取自国家级紫菜种质库Y-9101品系,文蛤壳、近江牡蛎壳、三角帆蚌壳、虾夷扇贝壳由如东华通水产品有限公司提供,海月蛤壳、鸡蛋壳、珍珠、方解石原石、霰石原石从市场购买,几丁质(Lot.G7TDKRJ)由东京化成工业株式会社生产,高黏度壳聚糖(Lot.C11029612)由上海麦克林生化科技有限公司生产。
1.2 仪器
光照培养箱(MGZ-120B-3,上海丙林电子科技有限公司);生物显微镜[E800,尼康仪器(上海)有限公司];光谱闪烁照度计(SFIM-300,杭州远方光电信息股份有限公司);扫描电子显微镜(JSM-6510,日本电子株式会社)。
1.3 方法
1.3.1 丝状体接种与培养
将文蛤壳、近江牡蛎壳、三角帆蚌壳、虾夷扇贝壳、海月蛤壳、鸡蛋壳、珍珠、方解石原石和霰石原石清洗干净,其中鸡蛋壳放入有效氯含量5%(m∶V)的84消毒液中浸泡12 h,使鸡蛋壳乳突层与蛋壳内侧的壳膜层纤维分离[18],用镊子将壳膜层清除干净。壳聚糖加入2%(V∶V)乙酸配置成5%(V∶V)壳聚糖溶液,将5%壳聚糖溶液倒入培养皿自然晾干形成壳聚糖薄膜,冲洗至pH中性。将条斑紫菜自由丝状体打碎成40-60 μm小段,分别接种于上述培养基质上,接种密度为150 ind./cm2,将每种贝壳各3枚、鸡蛋壳碎片3枚(每枚约10 cm2)、珍珠5粒放入培养盒培养,共设置4组平行。将方解石原石、霰石原石各3块、壳聚糖薄膜3张、碎片状几丁质1 g放入培养盒中共同培养,设置培养条件为光照强度(12.5±1.0) μmol/(m2·s),培养温度16℃,海水盐度27‰,光照周期12D∶12L。
1.3.2 着生密度与着生效率的测量
接种后每周用生物显微镜观察丝状体着生情况,接种6周后贝壳等基质表面藻斑肉眼清晰可见,统计藻斑数量,计算着生密度(着生密度=藻斑数/贝壳面积),从而计算出着生效率(着生效率=着生密度/接种密度×100%)。
1.3.3 贝壳丝状体长度的测量
接种2周起通过测量藻斑直径计算水平生长面积(藻斑近似圆形),至藻斑生长交连到一起时停止测量。分别于第55天与第100天将贝壳等基质沿着藻斑中心处纵切,采用扫描电镜测量丝状体纵向生长长度。
1.3.4 贝壳丝状体壳孢子放散测量
为了研究不同培养基质中贝壳丝状体壳孢子的放散水平,接种120 d后,分别取每种贝壳基质各1枚、珍珠5粒、鸡蛋壳碎片1枚(约10 cm2),置于50 mL海水中培养,设置4组平行。培养温度16℃,光照条件为7:00-12:30光强(31.3±1) μmol/(m2·s),12:30-18:30光强(6.3±1.0) μmol/(m2·s),18:30至次日7:00黑暗。连续20 d测定单位面积培养基质的日放散量[19],每天中午镜检壳孢子放散量后,清洗贝壳并更换海水。通过测量各种基质的接种面积(珍珠接种面积近似半球形),计算不同基质中壳孢子的日放散效率[壳孢子的日放散效率=壳孢子的日放散量/基质接种面积,spores/(cm2·d)]。
1.3.5 实验数据处理
各组实验数据采用GraphPad Prism 7 (GraphPad Software Inc.,美国)进行处理。数据显著性分析采用SPSS 22 (IBM,美国)进行单因素方差分析,设置显著水平P<0.05。
2 结果与分析
2.1 条斑紫菜丝状体着生情况及其不同培养基质微观结构
未发现条斑紫菜丝状体在方解石原石、霰石原石上着生。几丁质碎片面积为1-12 mm2,浸入海水后逐渐软化,表面未发现丝状体附着。条斑紫菜丝状体一端附着于壳聚糖薄膜表面,呈自由丝状体样向外生长,并未钻入薄膜内部,藻丝有少量单侧分枝,且分枝处无明显隆起[图1(a)]。而条斑紫菜丝状体着生于文蛤壳等5种贝壳基质以及鸡蛋壳和珍珠基质上,并向外生长与向内钻入生长,其中向外生长呈自由丝状体样,与壳聚糖薄膜表面生长的丝状体无异;向内钻入基质中,主分枝多于2条,向四周呈辐射生长,侧枝多呈对生,且多出现不定形隆起。在贝壳基质和珍珠基质中,条斑紫菜丝状体着生后先于基质表层生长后钻入内部[图1:(b)-(g)],而在鸡蛋壳中,丝状体着生后即钻入内部,呈弥散状[图1(h)]。

图1 条斑紫菜贝壳丝状体着生于不同培养基质的显微图
通过扫描电镜对基质纵切面结构和丝状体在基质中生长的痕迹进行观测,三角帆蚌壳、文蛤壳和珍珠外层由排列密集且整齐的细小文石多边形板片相互叠加构成[图2:(a)-(d)]。这3种基质剪切时断裂面整齐。近江牡蛎壳、虾夷扇贝壳和海月蛤壳由许多细长的菱形方解石板条层层叠加构成,切面在水平方向呈细条状纹路[图2:(e)-(f)],剪切时易产生多层滑动断裂。条斑紫菜贝壳丝状体在这6种基质内大多集中在基质的表层,且更容易发育为孢子囊枝,下层的贝壳丝状体数量较少,且呈营养藻丝形态纵向生长。鸡蛋壳主要分为两层结构,分别是蛋壳内侧的乳突层和蛋壳外侧的栅栏层,乳突层厚度约为100 μm,蛋壳厚度约为300-400 μm [图2(g)]。两层结构的表层均分布大量贝壳丝状体,且藻斑内外发生重叠,显示丝状体穿透蛋壳生长。

图2 不同条斑紫菜贝壳丝状体培养基质切面扫描电镜图
2.2 不同培养基质中条斑紫菜贝壳丝状体的着生效率
丝状体在近江牡蛎壳与鸡蛋壳上着生效率最大,分别为(4.5±0.6)%、(4.2±0.6)%(P≥0.05),其次为虾夷扇贝壳与三角帆蚌壳,在珍珠基质上的着生效率最低,为(1.7±0.6)%(图3)。

S:虾夷扇贝壳;H:三角帆蚌壳;M:近江牡蛎壳;D:鸡蛋壳;Z:珍珠;W:文蛤壳;J:海月蛤壳。相同小写字母表示不同组间差异未达显著水平(P≥0.05)
2.3 不同培养基质中条斑紫菜贝壳丝状体的分布
接种两周后丝状体开始钻入基质中并形成贝壳丝状体。在三角帆蚌、海月蛤、近江牡蛎、虾夷扇贝和文蛤等贝壳基质中贝壳丝状体于第4周水平生长速率最快,生长速率均超过1 000 μm/周,第6至第10周水平生长速率较慢,平均生长速率为321-429 μm/周。蛋壳基质中,贝壳丝状体在第3至第9周的平均水平生长速率为297 μm/周,第10周生长速率最快,达768 μm/周。珍珠基质中,贝壳丝状体在第3至第8周水平生长速率较慢,平均为317 μm/周,第9周生长速率最快,达736 μm/周。在三角帆蚌壳、近江牡蛎壳和文蛤壳中,贝壳丝状体前10周的平均水平生长速率为63.5 μm/d (P≥0.05);其次为虾夷扇贝壳,为(59.5±10.6) μm/d;鸡蛋壳中贝壳丝状体的水平生长速率最慢,为(39.9±6.9) μm/d(图4)。
营养藻丝阶段,贝壳丝状体在鸡蛋壳中钻入的深度最深,达(267.4±60.1) μm,其次为三角帆蚌壳与海月蛤壳,分别为(82.1±35.1) μm、(75.9±37.3) μm,而在近江牡蛎壳、虾夷扇贝壳、文蛤壳以及珍珠中的钻入深度基本相同(P≥0.05)(图4)。
第13周,贝壳丝状体由营养藻丝发育为孢子囊枝后继续生长,孢子囊枝在鸡蛋壳中的钻入深度最深,穿透蛋壳生长,达(334.9±15.0) μm;其次为虾夷扇贝壳,钻入深度为(168.5±44.8) μm;孢子囊枝在三角帆蚌壳、海月蛤壳、近江牡蛎壳、文蛤壳和珍珠中的钻入深度基本一致,为(113.9±27.9) μm(P≥0.05)(图4)。

S:虾夷扇贝壳;H:三角帆蚌壳;M:近江牡蛎壳;D:鸡蛋壳;Z:珍珠;W:文蛤壳;J:海月蛤壳。相同小写字母表示不同组间差异未达显著水平(P≥0.05)
2.4 不同培养基质对条斑紫菜壳孢子放散的影响
贝壳丝状体壳孢子前6 d平均日放散量均较低,后14 d平均日放散量差异较大,其中文蛤壳基质壳孢子日放散量最大,达624 spores/(cm2·d),其次为鸡蛋壳和海月蛤壳,分别达360 spores/(cm2·d)、223 spores/(cm2·d),三角帆蚌壳基质壳孢子日放散量最小,为41 spores/(cm2·d) (图5)。
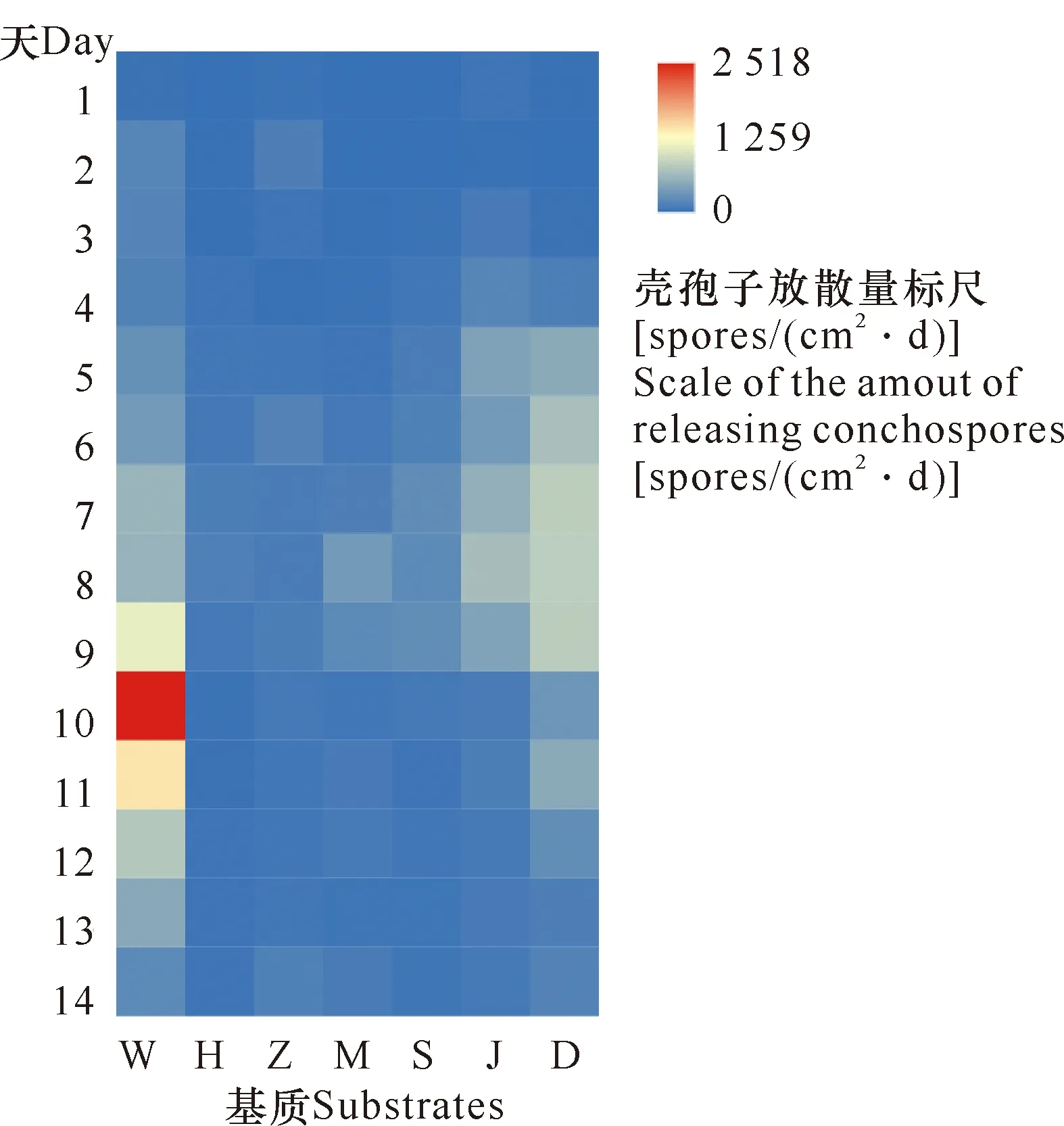
S:虾夷扇贝壳;H:三角帆蚌壳;M:近江牡蛎壳;D:鸡蛋壳;Z:珍珠;W:文蛤壳;J:海月蛤壳
3 讨论
紫菜种苗可采用平铺或吊挂的方式培养,由于不同贝壳的成分、结构以及质地有差别,导致贝壳丝状体的生长与壳孢子放散量存在较大的差异。本研究中文蛤壳、近江牡蛎壳、三角帆蚌壳、虾夷扇贝壳、海月蛤壳、鸡蛋壳和珍珠等7种丝状体培养基质均为常见的天然材料,主要成分均为碳酸钙,以及少量几丁质和蛋白质,贝壳丝状体在7种基质中的生长及发育水平差异明显,可能与培养基质的结构、碳酸钙结晶方式、晶体排列方式和有机成分等因素有关。Oaki等[20]对鸡蛋壳、贝壳珍珠层、海绵骨骼、珊瑚和有孔虫等5种不同生物矿物材料的纳米结构进行研究,发现其均由纳米晶粒聚合形成。从基质的结构及结晶方式来看,文蛤壳和三角帆蚌壳内侧的珍珠层均由文石晶体组成[21];珍珠外侧的珍珠层也是由文石晶体组成;虾夷扇贝壳的内层由文石晶体组成,厚度为250-300 μm[22];牡蛎壳的内层由方解石晶体形态构成[23];鸡蛋壳主要分为乳突层和栅栏层,其中的碳酸钙主要以方解石形态存在[24];而海月蛤壳为99%细长的菱形方解石条板构成的单叶微结构。从晶体排列方式来看,Cain等[25]测得鸡蛋壳中栅栏层的方解石晶体有着一致的取向性,晶体轴和蛋壳表面的垂直方向有16°-28°的倾斜角度,但是乳突层的晶体取向则是随机的;虾夷扇贝壳内层的方解石晶体具有相同的倾斜角度;海月蛤壳具有独特的透光性,可见光总透射率高达80%,这是因为其每个方解石板条衍射为单晶[26]。本研究中鸡蛋壳、虾夷扇贝壳、近江牡蛎壳和海月蛤壳的培养层为方解石结晶形式,文蛤壳、珍珠和三角帆蚌壳属于文石结晶形式。丝状体在这两种结晶形式的天然基质中均能发育为贝壳丝状体。
三角帆蚌、文蛤、牡蛎与虾夷扇贝的贝壳常用于紫菜育苗产业,这4种贝壳基质的结构及硬度均有所差异(表1)。本研究显示,在上述4种贝壳中,贝壳丝状体在三角帆蚌、文蛤与近江牡蛎的贝壳中水平生长速率高于虾夷扇贝壳,生长深度低于虾夷扇贝(P≥0.05)。文蛤壳中孢子放散水平显著高于其他3种贝壳,三角帆蚌壳最低,因此条斑紫菜壳孢子放散水平不能由水平生长速率和生长深度直接反映,还可能与培养基质的硬度等物理量有关。

表1 紫菜育苗产业常用贝壳结构及硬度
贝壳因其面积小、内表面不平整的特点,导致紫菜种苗培育过程中清洗、铺摆工序耗费大量人力与时间,且其易翻覆,降低了采苗效率,因此具有较高平整度与组装水平的贝壳替代基质是未来紫菜种苗基质发展的方向。本研究选取了与贝壳内层相同晶体结构的方解石原石、霰石原石,以及与贝壳内层具有共同成分的壳聚糖薄膜、几丁质碎片进行同步培养,未发现条斑紫菜丝状体生长于其中,仅在壳聚糖薄膜表面着生,并向外以自由丝状体形态生长,与钻入生物合成基质的贝壳丝状体形态有明显差异。从生物合成基质的有机成分分析,文蛤壳的有机质成分主要为几丁质、富含丙氨酸的不可溶蛋白质、富含甘氨酸和天冬氨酸等酸性氨基酸的可溶性蛋白等;牡蛎壳中含有几丁质、糖蛋白、珍珠蛋白、天冬氨酸、丝氨酸、谷氨酸和甘氨酸等[34-37];三角帆蚌壳和珍珠的珍珠层文石结晶间的有机质主要是几丁质纤维和类丝素蛋白,还含有甘氨酸和丙氨酸等[37];鸡蛋壳中的有机质主要为蛋白质、糖蛋白、蛋白多糖[38],这些天然生物基质中均含有1%-5%的有机成分[39-43],因此紫菜丝状体的着生与贝壳丝状体的形成可能与基质中的有机成分有关。
4 结论
条斑紫菜丝状体可在生物合成基质与壳聚糖薄膜表面着生并向外呈自由丝状体样生长,向内只在生物合成基质中形成贝壳丝状体,并发育至放散壳孢子,未发现生长于壳聚糖薄膜内,且未在方解石原石、霰石原石与几丁质碎片内部着生。贝壳丝状体在近江牡蛎壳与鸡蛋壳中着生效率最高,分别达(4.5±0.6)%、(4.2±0.6)%;在三角帆蚌、近江牡蛎和文蛤贝壳中水平生长速率最大,平均为63.5 μm/d(P≥0.05);在鸡蛋壳中纵向生长最深,达(334.9±15.0) μm;在文蛤贝壳中的壳孢子放散量最高,其次为鸡蛋壳。因此,除了常用于紫菜育苗生产的文蛤壳、近江牡蛎壳、三角帆蚌壳与扇贝壳外,鸡蛋壳较为适宜贝壳丝状体的着生、生长与壳孢子放散。
