岩溶石漠化不同修复林分林下植被和植物多样性的变化*
2022-04-12周晓果温远光孙冬婧明安刚贾宏炎朱宏光赵岩岩李锦良黄勇杰
周晓果,温远光,**,王 磊,孙冬婧,明安刚,贾宏炎,朱宏光,赵岩岩,李锦良,黄勇杰,梁 佳
(1.广西科学院生态环境研究所,广西南宁 530007;2.广西大学林学院,广西森林生态与保育重点实验室,广西南宁 530004;3.中国林业科学研究院热带林业实验中心,广西凭祥 532600;4.南宁市武鸣区城厢镇农林水利综合服务中心,广西南宁 530199)
林下植被是人工林生态系统的重要组成部分,它既受环境条件的深刻影响,同时也对环境产生持续性的改造作用[1]。林下植被的发育不仅能有效提高土壤有机质、促进养分循环,还能增加生物多样性、加速生态修复、维持生态系统功能稳定性[2,3]。物种组成和多样性是植被最基本的特征和群落结构的基础,是判断生物群落结构变化或生态系统稳定性的指标,成为群落生态学研究的核心问题[4]。对长期固定样地的监测研究已成为生物多样性研究领域的重点方向之一[5-8]。通过对林下植被的长期固定样地进行生物多样性动态变化监测,了解石漠化修复过程中人工林林下植被和多样性的变化规律,可为森林保护修复、经营管理和研究提供理论依据[9,10]。
西南岩溶地区石漠化综合治理是国家长期生态治理目标需求[11],涌现多种多样的治理模式,形成了封山育林、人工造林等修复模式和石山火龙果(Hylocereusundatus)、食用菌、赤苍藤(Erythropalumscandens)、金银花(Lonicerajaponica)等特色产业[12,13],为脱贫攻坚发挥了积极作用[14]。近年来,学者对岩溶地区不同恢复阶段植物群落结构和多样性变化进行了大量研究[5,15-17],Zeng等[18]研究了不同干扰因素对喀斯特自然植被恢复的影响;宋同清等[16]研究了喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征;温远光等[19]研究了广西马山岩溶植被年龄序列的群落特征;盛茂银等[17]研究了贵州喀斯特石漠化地区植物多样性与土壤理化性质的关系;陆志成等[20]研究了封育年限对植被组成和土壤肥力修复的影响。但是,这些研究绝大多数为一次性的短期调查,忽视了对植物群落演替规律的长期监测[5],也很少开展人工修复林分植物多样性的长期演替规律研究。最近,张芳等[5]基于2007年、2012年、2017年的3次植被调查,探讨了岩溶天然灌木林、次生林、原生林群落结构及多样性随时间的动态变化特征,一定程度上揭示了自然植被的动态变化规律。目前尚缺乏对岩溶石漠化不同修复林分植物多样性长期动态变化研究,对于岩溶石漠化地区人工修复林分是否能持续增加多样性、加速生物多样性修复以及动态变化规律尚不清楚。由于缺乏对石漠化植被修复群落的长期监测和评估,导致一些尚未经科学评估的技术、模式大规模应用,造成治理效果不佳甚至失败[14,15]。因此,开展石漠化修复林分的长期监测研究,并与治理前的灌丛群落植物多样性进行比较,将更好地揭示和了解石漠化地区人工修复林分植物多样性的演变机制。本研究基于2011年建立的顶果木(Acrocarpusfraxinifolius)纯林、降香黄檀(Dalbergiaodorifera)纯林和顶果木×降香黄檀混交林固定监测样地于2013年、2015年、2019年、2021年的4次植被调查,并以当地原有的石山灌丛为对照,探讨了岩溶石漠化不同修复林分林下植被物种组成及多样性随时间的变化特征,为进一步探索3种人工修复林分林下植被物种组成和物种多样性特征、森林更新及其长期动态变化提供基础数据和资料,为石漠化综合治理提供科学依据。
1 材料与方法
1.1 研究区域的自然环境概况
研究区域位于广西南宁市马山县(23°24′-24°2′N,107°41′-108°29′ E),境内岩溶地貌发育,是广西石漠化最严重的地区之一。地处南亚热带季风气候区,年均气温21.3℃,最高气温38.9℃,最低气温-0.7℃;年均降水量1 667.1 mm,最高2 063 mm,最低1 245.8 mm;雨量充沛,但降雨分布不均,夏季多暴雨,春秋两季少雨,易造成干旱;相对湿度76%;年平均无霜期343 d。土壤主要以碳酸盐岩发育的石灰土为主,石粒含量高,质地黏重,土层浅薄,岩基裸露度大,通常为40%-50%,高的达80%以上,土壤pH呈弱碱性。属南亚热带季雨林化常绿阔叶林地带、桂中石山青冈仪花青檀林区[21],原生顶极植被为以青冈栎(Cyclobalanopsisglauca)、海红豆(Adenantheramicrosperma)、青檀(Pteroceltistatarinowii)、小果化香(Platycaryaglandulosa)、仪花(Lysidicerhodostegia)等为优势的常绿落叶阔叶混交林[16]。这类原生森林几乎荡然无存,已退化为各种次生植被或人工植被[19]。
1.2 样方设置与群落调查
于2013年在马山县民族村2011年营造的顶果木纯林(PureA.fraxinifoliusPlantation,PAFP)、降香黄檀纯林(PureD.odoriferaPlantation,PDOP)和顶果木×降香黄檀混交林(MixedA.fraxinifoliusandD.odoriferaPlantation,MADP),分别选择坡度、坡向、土壤母质、土壤类型(棕色石灰土)等立地条件基本一致的坡面,建立长期固定监测样地,样地面积20 m×20 m,每种林分设3个重复,在每个400 m2样地随机设置3个5 m×5 m小样方,记录物种名称、个体数、高度、盖度和生长状况等。同时,在原有的灌丛(Shrub Land,SL)中设置灌丛群落样地,每个样地面积为5 m×5 m,共15个。分别于2015年、2019年和2021年进行样地复查。
1.3 重要值计算
采用每种林下植物物种的重要值(Importance Value,IV)来表征其在相应群落中的地位和优势大小,计算方法如下[22]:
IV=100×(Ra+Rf+Rc)/3,
(1)
式(1)中,Ra为相对多度,为样方中某个种的个体数占全部种的个体总数的百分比;Rf为相对频度,为样方中某个种出现的频度占全部种的频度的百分比;Rc为相对盖度,为样方中某个种的盖度占全部种的盖度的百分比。
1.4 物种多样性的计算
1.4.1α多样性计算
根据4次调查数据,对表征林下植被物种α多样性基本特征的各多样性指数按下列公式计算[23]。
物种丰富度指数(S):S=样地内出现的物种数,
(2)
(3)
(4)
(5)
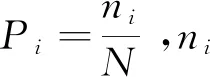
1.4.2β多样性计算
β多样性指数反映群落间物种组成差异及其物种多样性对环境梯度变化的响应。本研究采用Cody指数和Sφrensen指数来度量。计算公式如下[24,25]:
Cody指数(βc):βc=(b+c)/2,
(6)
Sφrensen指数(CS):CS=2a/(2a+b+c),
(7)
式(6)、(7)中,a是两个样地内共同拥有的物种数目,b是样地A独有的物种数目,c是样地B独有的物种数目。
1.5 数据统计分析
在SPSS 24.0 for Windows中,采用单因素方差分析(One-way ANOVA)检验不同林分林下植被群落物种多样性的差异显著性,采用最小显著性差异法(Least Significant Difference,LSD)进行多重比较,显著性水平设置为P<0.05。采用Bray-Curtis距离的非限制性主坐标轴分析(Principal Co-ordinates Analysis,PCoA)揭示不同修复模式物种组成的差异及变化趋势。
2 结果与分析
2.1 灌丛和3种不同修复林分林下植被科、属、种及个体数的变化
3种不同修复林分林下灌木层植物科、属、种数目呈递增趋势,其中以PDOP的数值较大,由2013年的18科25属27种增至2021年的24科30属31种;其次是MADP,由2013年的14科25属26种增至2021年的15科27属29种;PAFP数值最小,由2013年12科15属17种增至2021年的15科20属23种。相反,SL群落灌木层植物的科、属、种数目呈递减趋势,由2013年的13科21属21种减至10科11属11种(表1)。
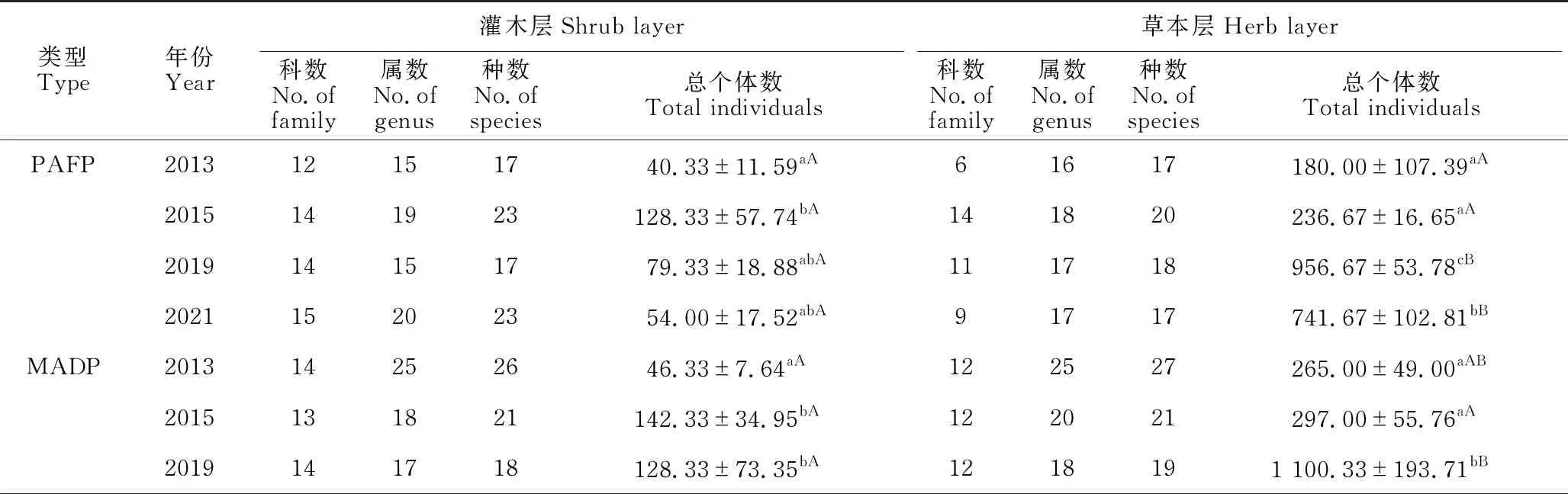
表1 灌丛和3种不同修复林分林下植被灌木层和草本层植物科、属、种及总个体数的变化
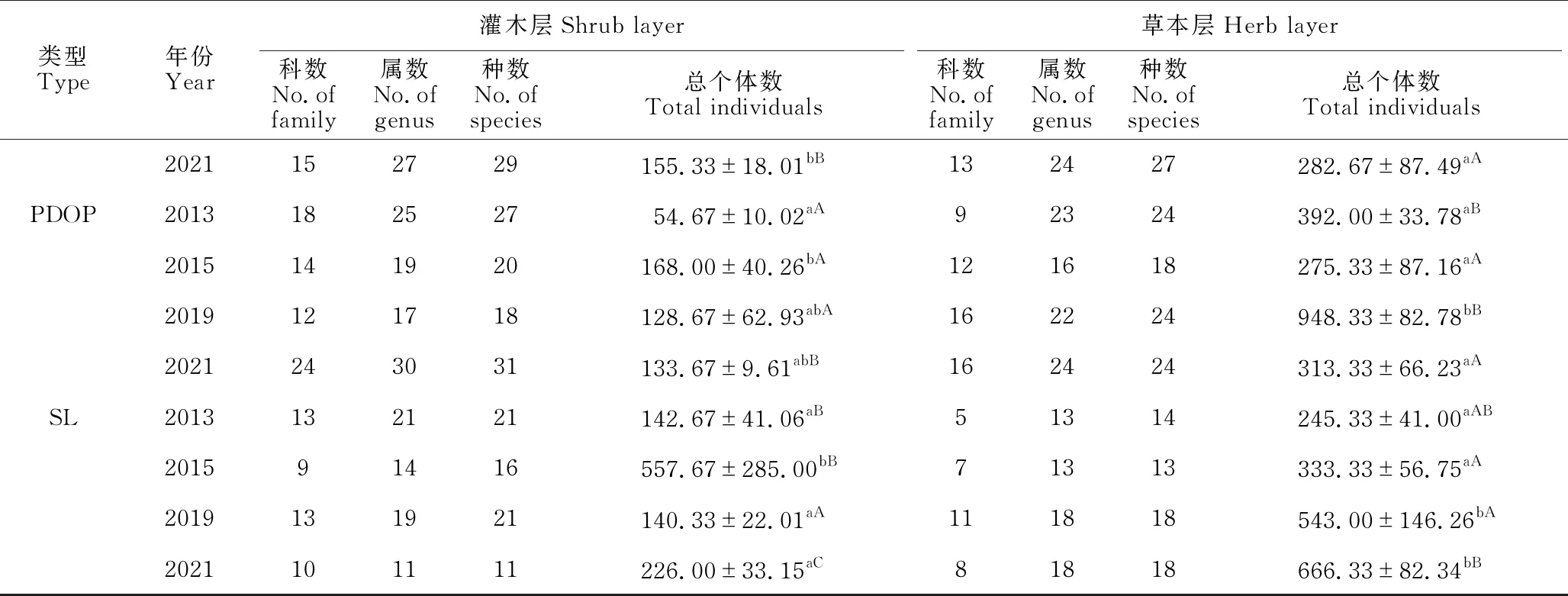
续表
草本层植物科、属、种的变化与灌木层的明显不同(表1)。与灌木层相反,3种不同修复林分林下草本层植物科、属、种数目的变化并不一致,除PAFP和PDOP草本层植物科数有所增加外,所有林分林下草本层植物属、种数目的变化极少。而SL草本层植物科、属、种数目均呈现明显的增加,由2013年的5科13属14种增至2021年的8科18属18种(表1)。
不同修复类型灌木层和草本层总个体数的年变化表现为PAFP和PDOP灌木层2015年的总个体数显著高于2013年,其他年份间差异不显著;MADP灌木层2013年总个体数显著低于其他年份,其他年份间差异不显著;而SL灌木层2015年的总个体数显著高于其他年份,其他年份间差异不显著。不同修复类型草本层总个体数的变化与灌木层不同,PAFP和SL为2019年和2021年草本层的总个体数显著高于其他年份,其他年份间差异不显著(在PAFP中,2019年还显著高于2021年);而MADP和PDOP相似,2019年草本层总个体数显著高于其他年份,其他年份间差异不显著(表1)。
同一年份不同修复类型灌木层和草本层总个体数的变化表现为2013年和2015年SL灌木层的总个体数显著高于3种修复林分,3种人工修复林分间差异不显著;2019年各类型间没有显著差异;2021年SL灌木层显著高于3种人工修复林分,MADP和PDOP显著高于PAFP,前两者差异不显著。2013年PDOP草本层总个体数显著高于PAFP,其他两两间差异不显著;2015年各类型间差异不显著;2019年3种人工修复林分草本层总个体数显著高于SL草本层,3种人工修复林分间差异不显著;2021年则表现为PAFP和SL草本层总个体数显著高于MADP和PDOP,其余差异不显著(表1)。
2.2 灌丛和3种不同修复林分林下植被组成物种重要值的变化
(1)灌木层。
PAFP 2013年灌木层植物的重要值排前5的物种是苎麻(Boehmerianivea)、黄荆(Vitexnegundo)、构树(Broussonetiapapyrifera)、地桃花(Urenalobata)和菜豆树(Radermacherasinica),前4种的重要值均大于10。2021年灌木层植物的重要值排前5的物种是黄荆、金花茶(Camelliapetelotii)、鸡矢藤(Paederiafoetida)、菜豆树和粗叶悬钩子(Rubusalceifolius),除了黄荆重要值超过20外,其他物种的重要值均小于10。2013年和2021年重要值≥1的木本植物分别有16种和22种,分别占总重要值的99.00%和99.02%。2013-2021年,有9个物种退出群落(或重要值<1),新出现15个物种,由于2019年在林下种植金花茶,因此金花茶增加最多,苎麻减少最多(表2)。
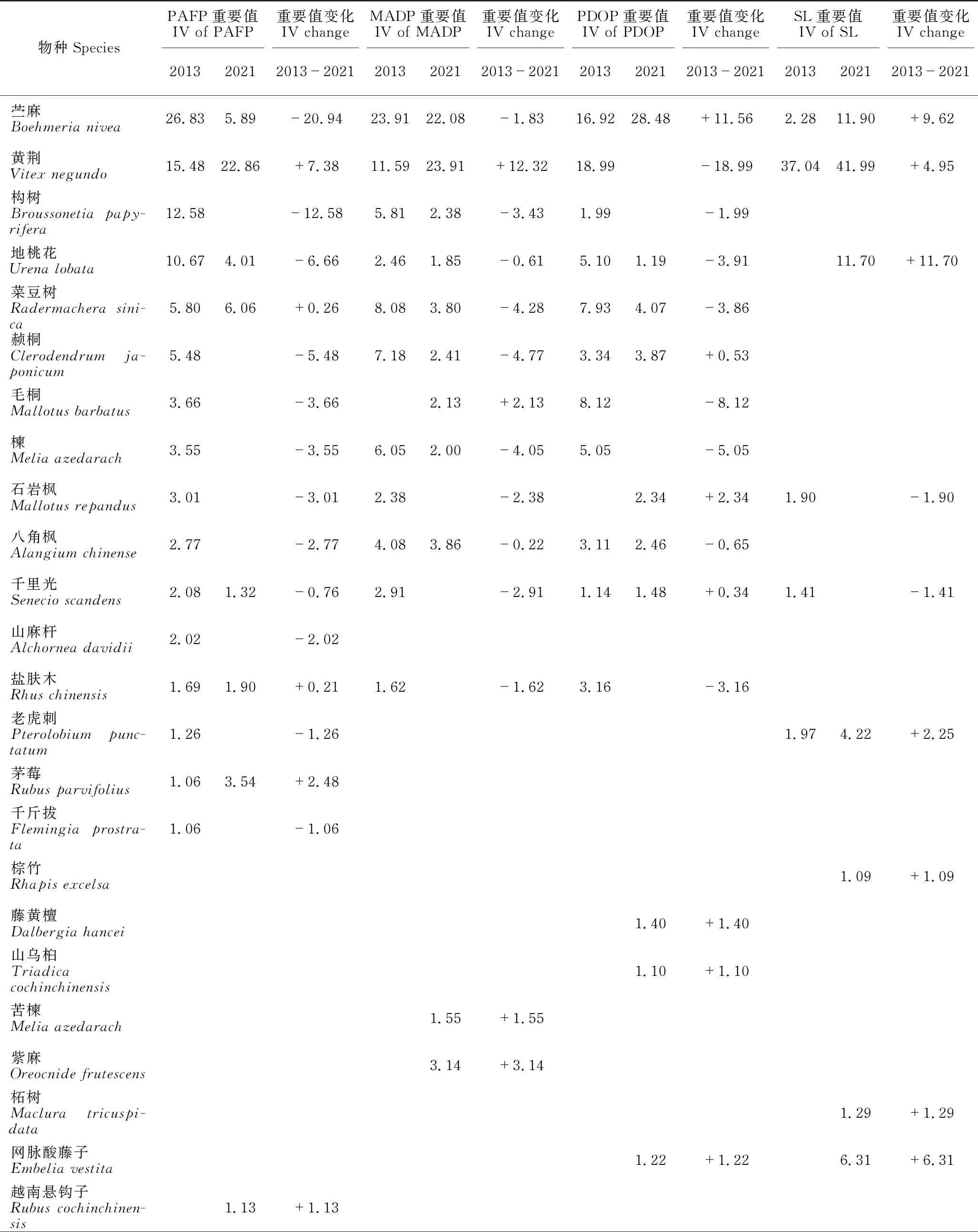
表2 灌丛和3种不同修复林分林下植被灌木层物种重要值的变化(重要值≥1)
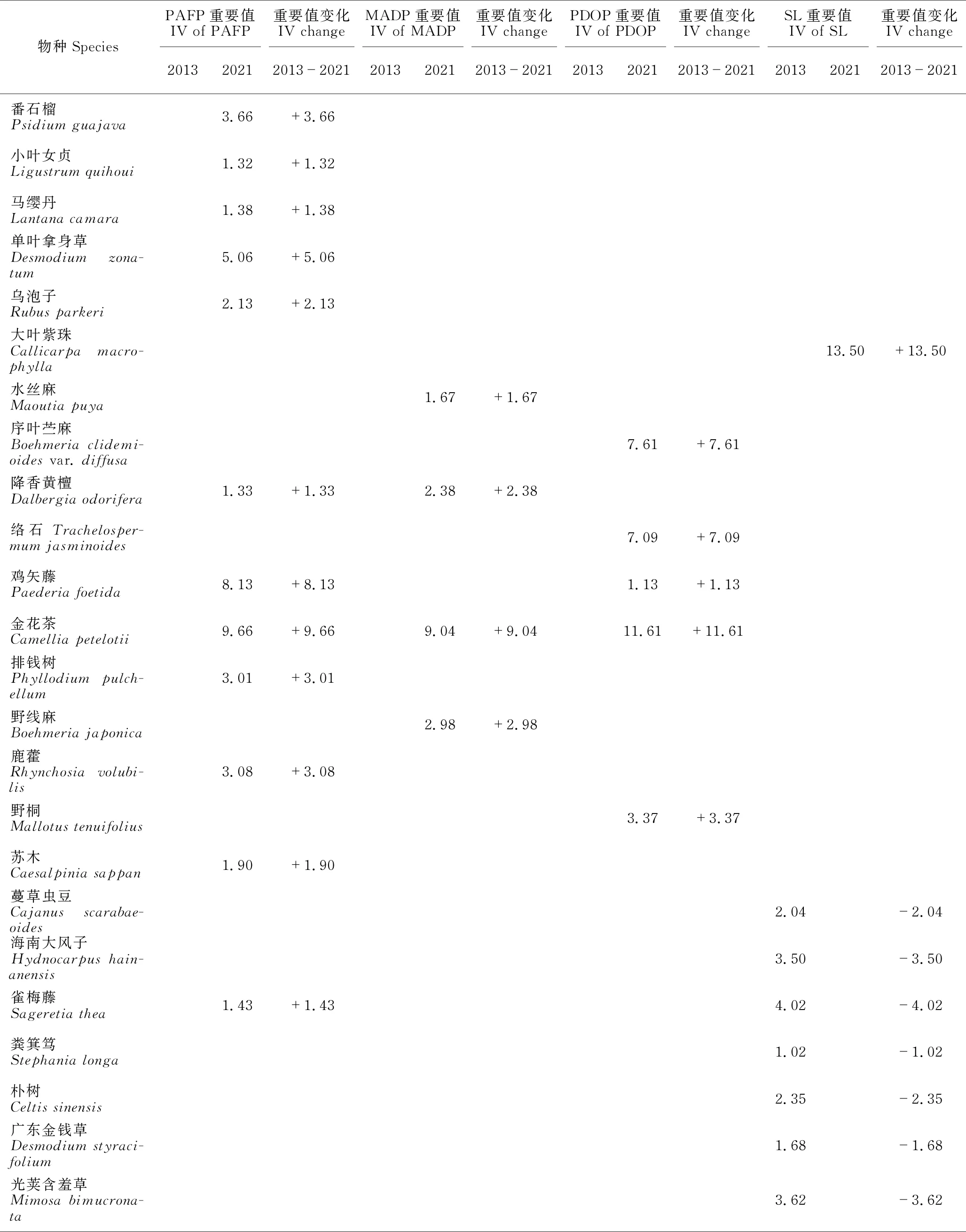
续表

续表
MADP 2013年灌木层植物的重要值排前5的物种是苎麻、黄荆、菜豆树、赪桐(Clerodendrumjaponicum)和楝(Meliaazedarach),前2种的重要值均大于10。2021年灌木层植物的重要值排前5的物种是黄荆、苎麻、金花茶、八角枫(Alangiumchinense)和菜豆树,除了苎麻和黄荆重要值超过20外,其他物种的重要值均小于10。2013年和2021年重要值≥1的木本植物分别有20种和21种,分别占总重要值的95.41%和94.93%。2013-2021年,有8个物种退出群落(或重要值<1),新出现9个物种,黄荆增加最多,赪桐减少最多(表2)。
PDOP 2013年灌木层植物的重要值排前5的物种是黄荆、苎麻、红背山麻杆(Alchorneatrewioides)、毛桐(Mallotusbarbatus)和菜豆树,除了前2种的重要值大于15,其余均小于10。2021年灌木层植物的重要值排前5的物种是苎麻、乌蔹莓(Cayratiajaponica)、金花茶、序叶苎麻(Boehmeriaclidemioidesvar.diffusa)和络石(Trachelospermumjasminoides),苎麻重要值超过20,乌蔹莓和金花茶重要值超过10,其他物种的重要值均小于10。2013年和2021年重要值≥1的木本植物分别有20种和16种,分别占总重要值的94.17%和90.35%。2013-2021年,有14个物种退出群落(或重要值<1),新出现10个物种,乌蔹莓增加最多,黄荆减少最多(表2)。
SL 2013年灌木层植物的重要值排前5的物种是黄荆、红背山麻杆、假马鞭(Stachytarphetajamaicensis)、雀梅藤(Sageretiathea)和光荚含羞草(Mimosabimucronata),黄荆和红背山麻杆重要值占绝对优势,分别为37.04和27.09,其余均小于5。2021年灌丛群落灌木层植物的重要值排前5的物种是黄荆、大叶紫珠(Callicarpamacrophylla)、苎麻、地桃花和粗叶悬钩子,除了粗叶悬钩子的重要值小于10外,其他均超过10,黄荆占绝对优势,重要值达41.99。2013年和2021年重要值≥1的木本植物分别有14种和9种,分别占总重要值的94.59%和98.37%。2013-2021年,有11个物种退出群落(或重要值<1),新出现6个物种,大叶紫珠增加最多,红背山麻杆减少最多(表2)。
(2)草本层。
PAFP 2013年草本植物的重要值排前5的物种是蒮香蓟(Ageratumconyzoides)、白茅(Imperatacylindrica)、水蔗草(Apludamutica)、蔓生莠竹(Microstegiumfasciculatum)和渐尖毛蕨(Cyclosorusacuminatus),前4种的重要值均大于10。2021年草本层植物的重要值排前5的物种是五节芒(Miscanthusfloridulus)、肾蕨(Nephrolepiscordifolia)、鬼针草(Bidenspilosa)、白茅和华南毛蕨(Cyclosorusparasiticus),前4种的重要值超过10,以五节芒的重要值最高,达28.24。2013年和2021年重要值≥1的草本植物分别有17种和11种,分别占总重要值的100.01%和95.36%。2013-2021年,有12个物种退出群落(或重要值<1),新出现6个物种,五节芒增加最多,蒮香蓟减少最多,基本退出群落(表3)。
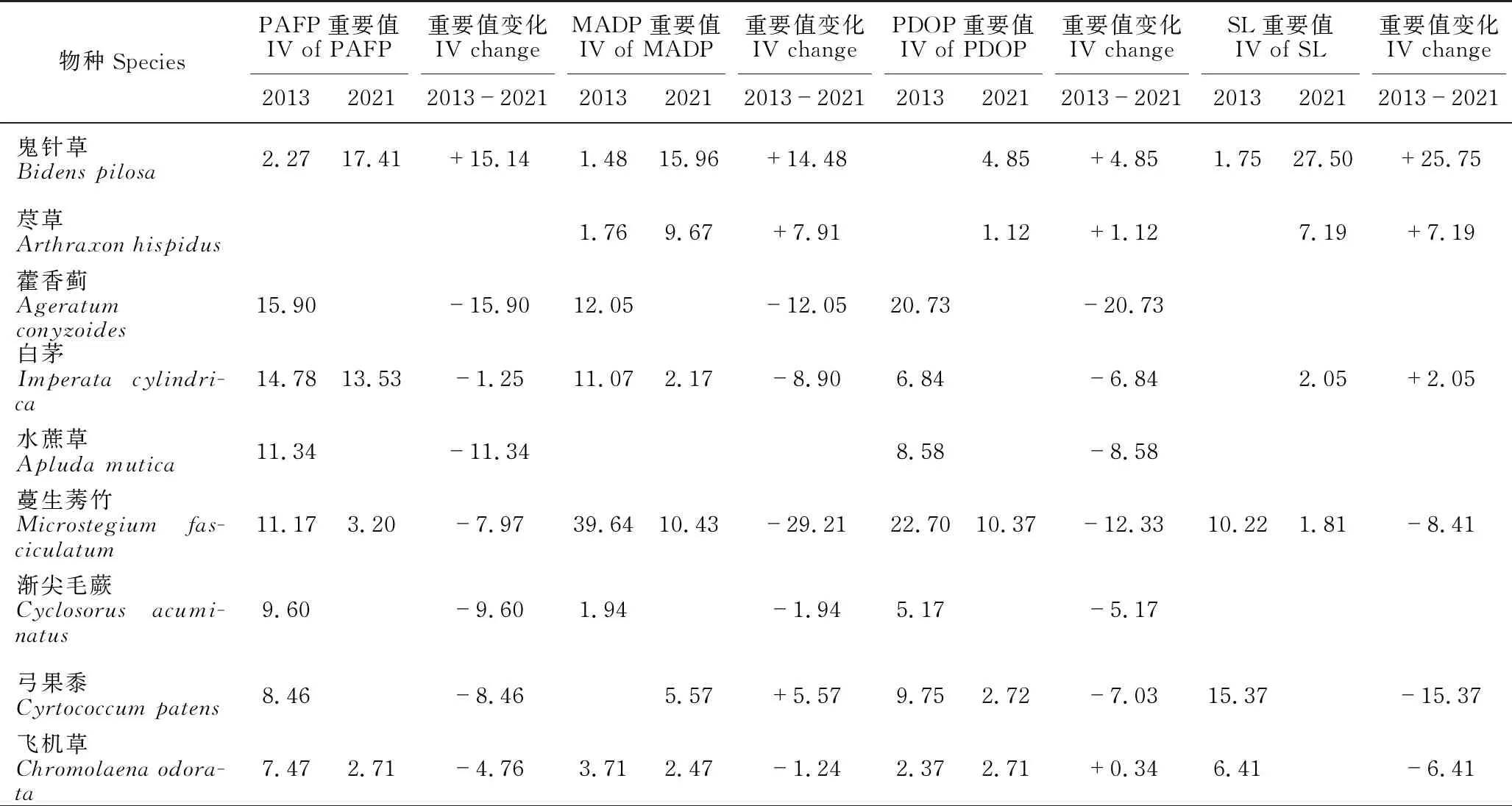
表3 灌丛和3种不同修复林分林下草本植物重要值的变化(重要值≥1)
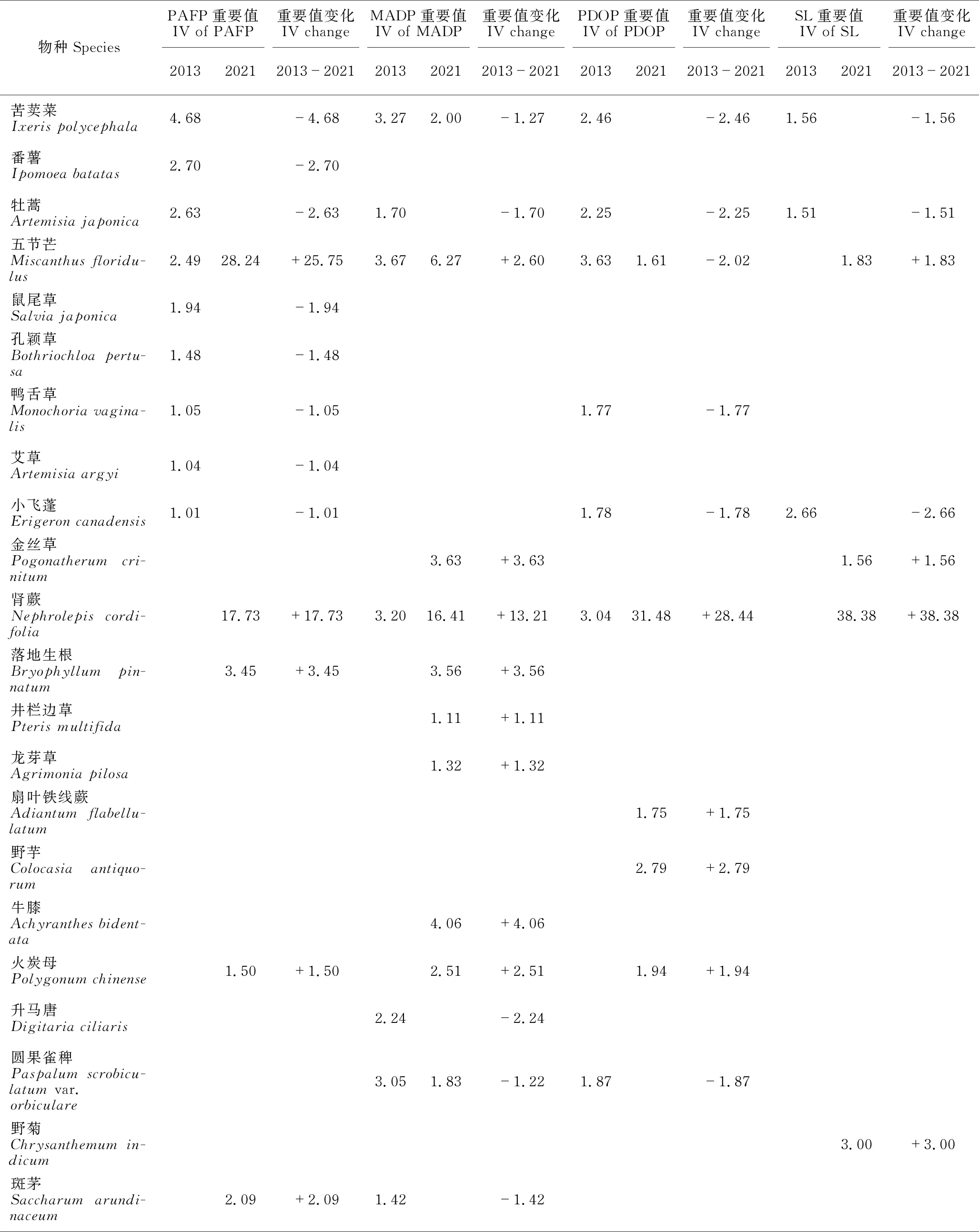
续表
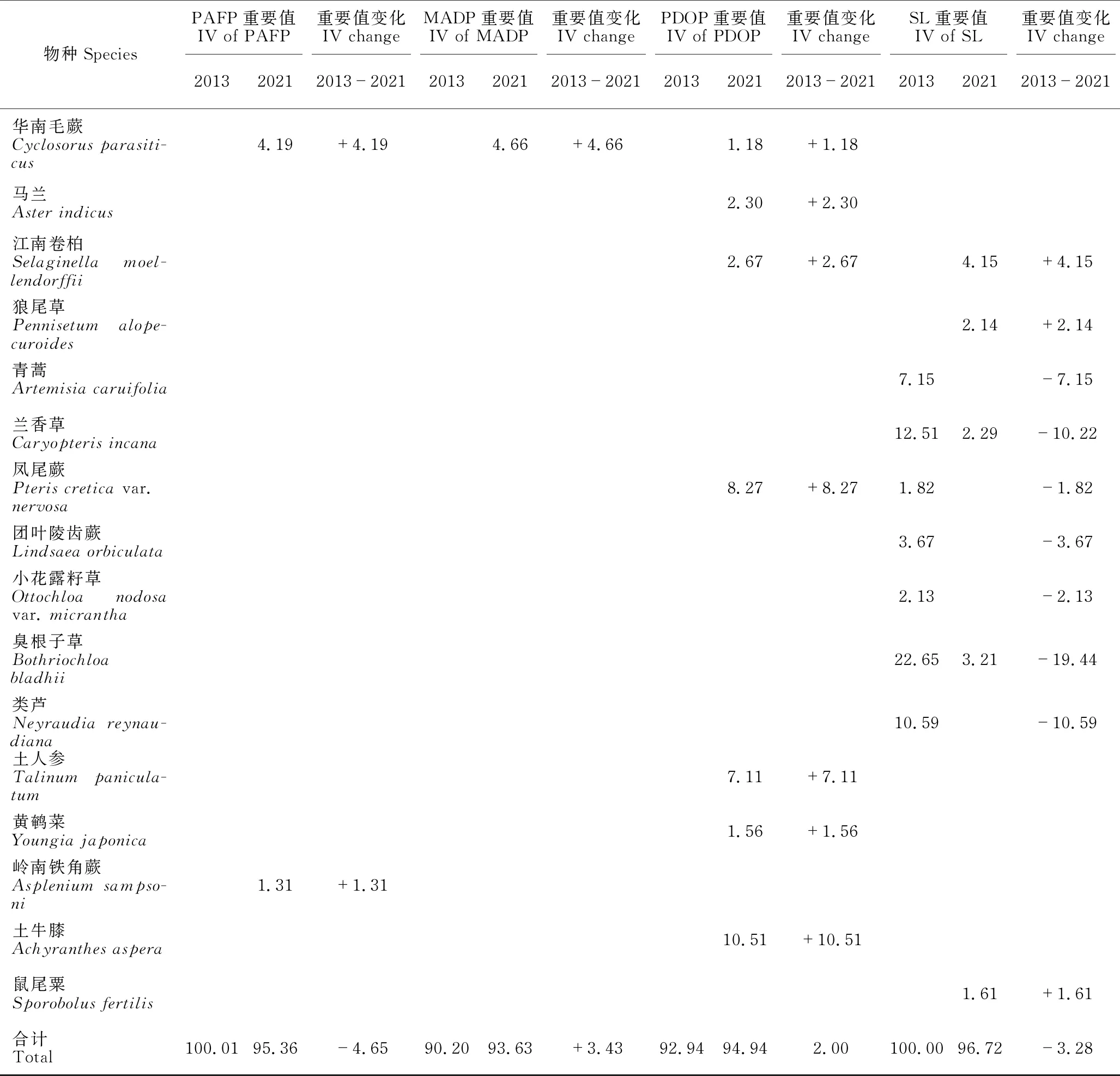
续表
MADP 2013年草本层植物的重要值排前5的物种是蔓生莠竹、蒮香蓟、白茅、飞机草(Chromolaenaodorata)和五节芒,前3种的重要值均大于10。2021年草本层植物的重要值排前5的物种是肾蕨、鬼针草、蔓生莠竹、荩草(Arthraxonhispidus)和五节芒,除了肾蕨、鬼针草、蔓生莠竹的重要值超过10外,其他物种的重要值均小于10。2013年和2021年重要值≥1的草本植物分别有14种和17种,分别占总重要值的90.20%和93.63%。2013-2021年,有5个物种退出群落(或重要值<1),新出现8个物种,鬼针草增加最多,蔓生莠竹减少最多(表3)。
PDOP 2013年草本层植物的重要值排前5的物种是蔓生莠竹、蒮香蓟、弓果黍(Cyrtococcumpatens)、水蔗草和白茅,除了蔓生莠竹和蒮香蓟的重要值大于20,其余均小于10。2021年草本层植物的重要值排前5的物种是肾蕨、蔓生莠竹、土牛膝(Achyranthesaspera)、凤尾蕨(Pteriscreticavar.nervosa)和土人参(Talinumpaniculatum),肾蕨占明显优势,重要值超过30,蔓生莠竹和土牛膝重要值超过10,其他物种的重要值均小于10。2013年和2021年重要值≥1的草本植物分别有14种和17种,分别占总重要值的92.94%和94.94%。2013-2021年,有9个物种退出群落(或重要值<1),新出现12个物种,肾蕨增加最多,蒮香蓟减少最多,基本退出群落(表3)。
SL 2013年草本层植物的重要值排前5的物种是臭根子草(Bothriochloabladhii)、弓果黍、兰香草(Caryopterisincana)、类芦(Neyraudiareynaudiana)和蔓生莠竹,5个物种的重要值均大于10。2021年草本层植物的重要值排前5的物种是肾蕨、鬼针草、荩草、江南卷柏(Selaginellamoellendorffii)和臭根子草,除了肾蕨和鬼针草的重要值大于20外,其他均小于10。2013年和2021年重要值≥1的草本植物分别有14种和13种,分别占总重要值的100.00%和96.72%。2013-2021年,有10个物种退出群落(或重要值<1),新出现9个物种,肾蕨增加最多,臭根子草减少最多(表3)。
2.3 灌丛和3种不同修复林分林下植被物种多样性的变化
(1)α多样性。
双因素方差分析结果显示,不同修复林分对灌木层和草本层物种丰富度、Shannon-Wiener指数、Simpson指数和Pielou均匀度指数均具有显著影响(草本层植物Pielou均匀度指数除外)(P<0.05)。年份对灌木层Pielou均匀度指数以及对草本层物种丰富度、Shannon-Wiener指数、Simpson指数和Pielou均匀度指数均具有显著影响(P<0.05)。修复类型和年份的交互作用除了对灌木层和草本层的Pielou均匀度指数存在显著影响(P<0.05)外,对其他α多样性指数没有显著影响(P>0.05)(表4)。
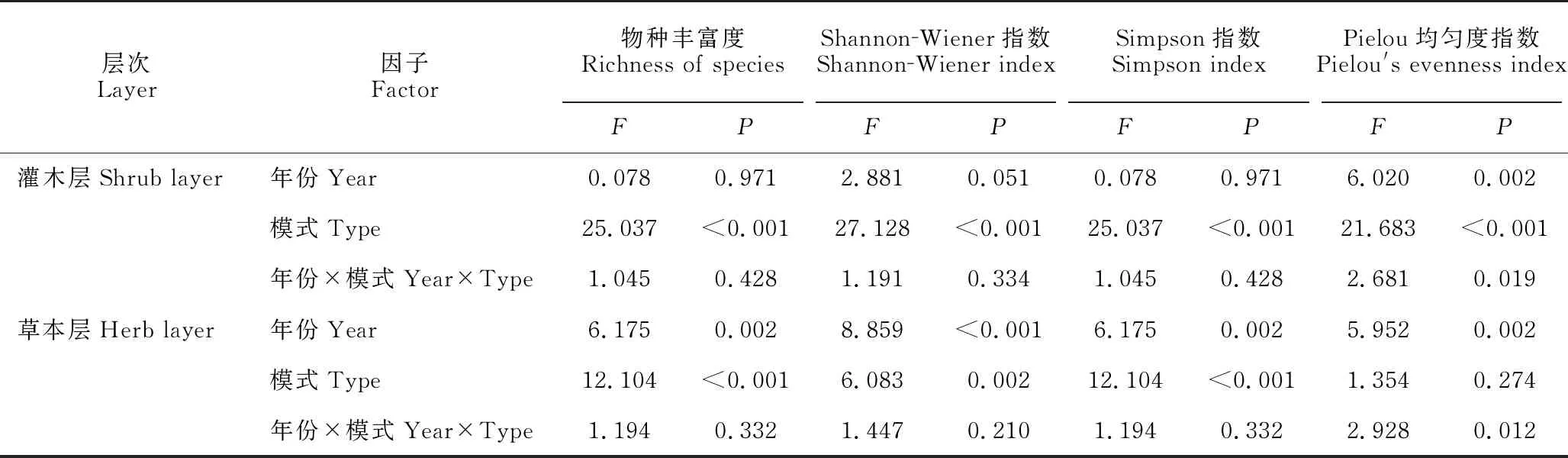
表4 灌丛和3种不同修复林分林下植物群落α多样性指数双因素方差分析
由图1可知,随着植被恢复时间的增加,3种人工修复林分林下灌木层的α多样性呈现相似的变化规律,Shannon-Wiener指数、Simpson指数和Pielou均匀度指数的变化相似,呈先减少后增加再减少的趋势;而灌丛群落灌木层的Shannon-Wiener指数、Simpson指数和Pielou均匀度指数的变化相似,呈先陡然减少后急速增加的趋势[图1:(a)、(b)、(c)]。
随着植被恢复时间的增加,灌丛和3种人工修复林分林下草本层的α多样性的变化有所不同。MADP的Shannon-Wiener指数、Simpson指数和Pielou均匀度指数的变化相似,呈先增加后减少再增加的“波浪形”变化趋势,PAFP表现为先增后降的“抛物线”变化格局,而PDOP草本层的Shannon-Wiener指数、Simpson指数则呈先增加(2019年达到峰值)后下降的趋势;灌丛草本层3种指数变化相似,呈先增加后急速减少的趋势[图1:(d)、(e)、(f)]。
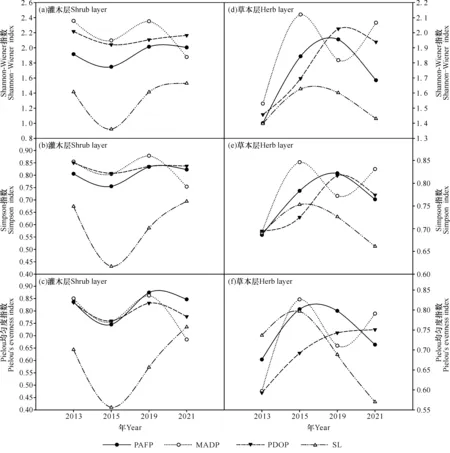
图1 灌丛和3种不同修复林分林下植被灌木层和草本层α多样性的变化
(2)β多样性。
不同植被修复类型灌木层、草本层物种组成均存在显著差异,并随调查时间变化(图2)。基于Bray-Curtis距离的非限制性主坐标轴分析(PCoA)表明,灌木层物种组成明显分为两大类,一类是灌丛群落,另一类是3种人工林修复类型,两大类在第2轴分开,见图2(a)。对于草本层,在2013年,MADP物种组成与其他3种植被类型差异显著,并在第2轴分开,见图2(b);在2015年,两种人工纯林(PAFP和PDOP)物种组成差异不大,但在第1轴与混交林(MADP)和灌丛群落分开,混交林与灌丛群落的物种组成则在第2轴分开;在2019年,3种人工林修复类型的物种组成差异不大,并在第1轴与灌丛群落分开;在2021年,3种人工林修复类型的物种组成则趋向一致(图2)。
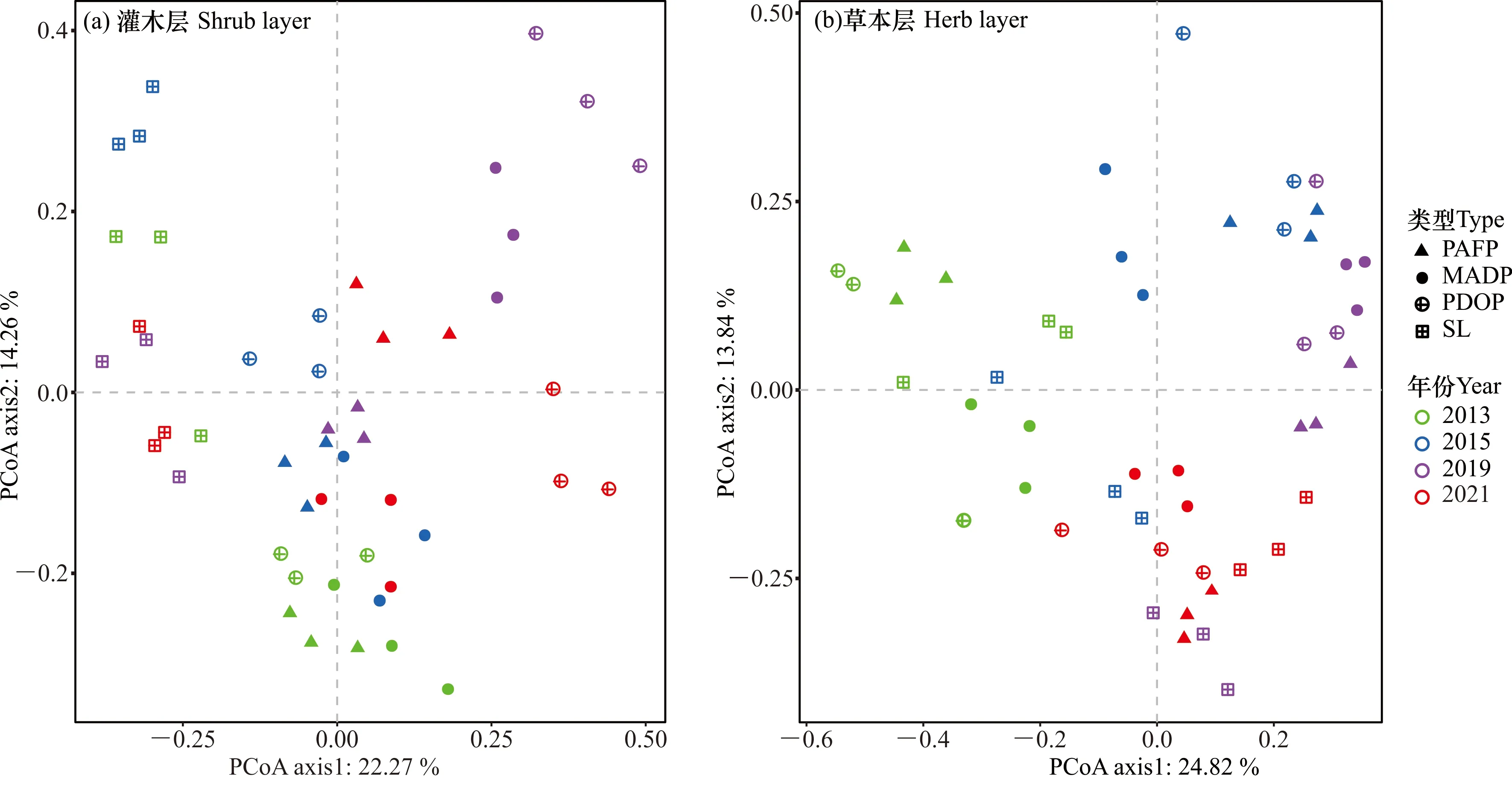
图2 灌丛和3种不同修复林分林下灌木层、草本层物种组成随时间的变化
不同植被修复类型灌木层、草本层物种β多样性指数随调查时间变化(表5),3种修复林分与灌丛群落之间灌木层和草本层的Sφrensen指数和Cody指数均随林龄增加而增加。在2013年和2015年,3种修复林分之间灌木层和草本层Sφrensen指数和Cody指数差异并不大;而2019年和2021年Sφrensen指数和Cody指数均增加,3种修复林分之间灌木层和草本层共有物种数持续减少。
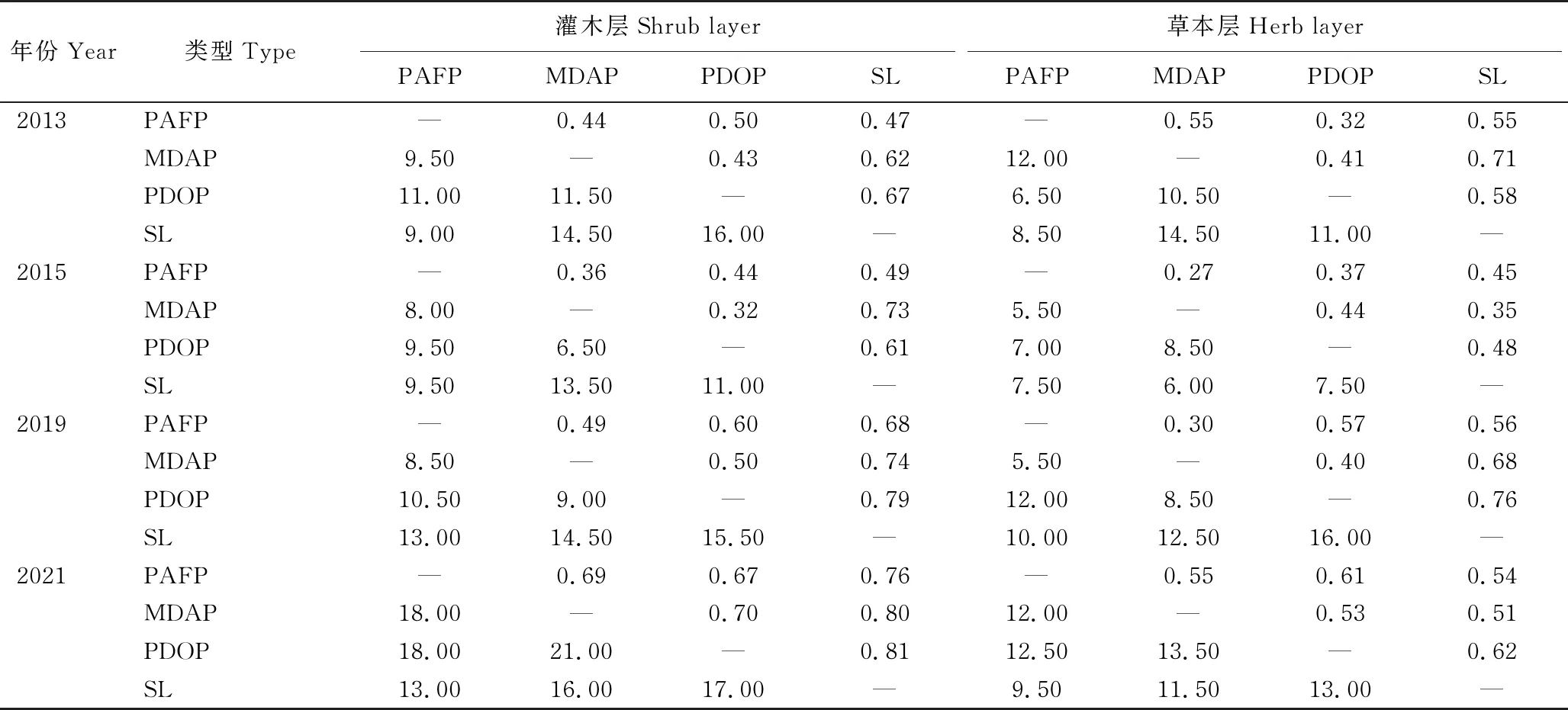
表5 灌丛和3种不同修复林分林下灌木层和草本层物种β多样性
3 讨论
本研究发现,灌丛和3种不同修复林分林下植被的总个体数存在明显波动,且草本植物比木本植物波动更大。这与岩溶地区生态环境恶劣,环境胁迫导致群落个体数量出现较大波动的结论相符[5]。张芳等[5]研究发现,2007-2017年,岩溶地区天然灌木林、次生林、原生林总个体数存在明显波动,以减少为主,认为可能是2009年的干旱和2015年的暴雨洪涝等导致植物大量死亡,有限时间内个体补员数量不足,导致样地内木本植物个体数减少[5]。本研究还发现,灌丛群落和两种人工纯林修复林分灌木层总个体数的最高值均出现在2015年,而混交林(MADP)灌木层总体个数在2015年时为次高水平;3种人工修复林分草本层总个体数的最高值均出现在2019年,而灌丛群落草本层总个体数在2019年时为次高水平。这与2015年和2019年研究区域(马山县)的降水量较稳定,分布比较均匀,没有明显干旱有关。据马山县气象观测数据,2014年有8个月的干旱期(月降水量<100 mm),2015年和2019年只有3个月,其余年份的干旱期也均在6-7个月,干旱期的长短或周期性波动对缺水严重的岩溶石漠化地区影响很大,从而导致群落不同层次植物总个体数的波动[5,26]。
植被的物种组成与多样性变化受到诸多因素的影响,岩溶石漠化区域气候干旱,缺土少水,种群发育受到更强烈的环境胁迫[5]。研究表明,岩溶天然灌木林和次生林群落受到一定程度的人为干扰,优势种群单一,而原生林群落环境相对稳定,优势种群不明显,形成多优势种群落[5,26]。本研究中,无论是灌丛还是人工修复林分林下灌木层,优势种主要是苎麻和黄荆,群落物种数量出现波浪式的变化,反映出岩溶石漠化地区生境恶劣的共同特征。然而,研究发现3种人工修复林分林下草本层物种组成和丰富度变化规律与灌丛明显不同,3种人工修复林分呈现出较高的物种组成和丰富度,并且随着林分的生长和环境的改善,群落物种数量表现为波浪式递增,而灌丛群落则表现出波浪式递减。其中又以MADP和PDOP的物种数量较高。这可能与MADP和PDOP两种修复类型的林分郁闭度较高,林内环境比较湿润,一些喜阴湿环境的物种增多有关,而PAFP修复林分开始出现衰退迹象,林分枯死木增多,郁闭度下降,林下环境比较干燥,使得物种多样性的恢复减缓。
与2019年相比,2021年PAFP、MADP和PDOP的物种数分别比2019年提高35.29%、61.11%和72.22%,而灌丛的物种数比2019年减少47.62%。这与人工修复林分生态环境的快速、持续改善密切相关,也与灌丛区域大量放养黑山羊对生态环境和植物群落的持续、破坏性干扰有关。此外,2019年在林下引入金花茶,金花茶的保存率在90%以上,成为林下植被的优势种之一,表明在岩溶石漠化植被恢复后,在林下种植高价值的金花茶是可行的。研究表明,天然灌木林、次生林和原生林群落中新增和减少的物种均为偶见种或稀有种[27],群落中偶见种或稀有种的增加和减少引起群落物种数量的波动[28]。本研究发现,在人工修复林分中,不仅是偶见种或稀有种的新增和减少导致群落组成的波动,林下植被优势种个体数量的增加和减少,同样引起群落灌木层和草本层组成和结构的波动,而且对物种重要值分配格局产生重要影响。
前人研究表明,天然次生林和原生林Shannon-Wiener指数总体呈增加趋势,而灌木林的α多样性指数随恢复时间增加而降低[5]。本研究表明,不同岩溶石漠化修复类型、不同群落层次物种α多样性的变化存在差异,PAFP灌木层的Shannon-Wiener指数、Simpson指数和Pielou均匀度指数随恢复时间的增加,总体呈微弱增加趋势,MADP和PDOP则相应为总体微弱递减趋势;而灌丛则表现为2015年急速降低后又快速回升,总体呈微弱增加趋势。进一步分析发现,MADP和PDOP林下灌木层群落优势种重要值增加,均匀度下降,导致α多样性指数微弱降低。PAFP草本层的3种指数总体呈2015年急速上升后2019年又快速下降,MADP和PDOP则相应为总体增加趋势;灌丛草本层3种指数均于2015年上升后又持续下降,且总体呈减少趋势。3种人工修复林分新增物种数高于死亡物种数,导致物种丰富度和多样性指数增加,而灌丛新增物种数小于死亡物种数,导致物种丰富度显著下降和多样性指数降低。本研究表明,新增或死亡的物种通常是偶见种或稀有种,与前人研究结果一致[5]。
β多样性又称生境间或群落间的多样性,是指沿某一环境梯度不同生境群落之间物种组成的相异性或物种沿环境梯度的更替速率[29,30],是度量群落演替过程中物种组成和生境变化的重要指标[5,31,32]。有研究表明,群落β多样性随着时间推移逐渐减少[33],随着人工林生长发育阶段越接近老龄林,其林下植物β多样性指数越小,低地位指数的样地林下植物β多样性指数值明显低于高地位指数的样地[32]。本研究表明,随着时间的推移,3种人工修复林分与灌丛之间灌木层和草本层的Sφrensen指数和Cody指数均随林龄增加而增加,表明人工林修复类型与灌丛共有物种数持续减少,群落演替速率持续升高。这说明3种人工修复林分与灌丛群落之间的林下植物种类组成和群落生境存在较大差异。主要原因是人工造林和植被保护增加了物种多样性,森林小气候的形成和凋落物的分解改善了群落生态环境[17,34],而灌丛地持续强度放牧和人为干扰,导致植被盖度减少、植物多样性丧失和生态环境退化[11,17]。这与环境异质性增加导致群落相似性降低的生态位理论一致[35]。本研究中,3种人工修复林分之间灌木层和草本层的Sφrensen指数和Cody指数差异不大,以及2019年和2021年Sφrensen指数和Cody指数增加,说明3种人工修复林分之间灌木层和草本层共有物种数持续减少、群落演替速率升高,加速植被恢复进程。
