城市景观格局视角下空气微生物研究进展
2022-04-11吴志丰蒋梧州
吴志丰,邱 月,2,任 引,蒋梧州,杨 磊
1 中国科学院城市环境研究所,城市环境与健康重点实验室, 厦门 361021
2 中国科学院大学, 北京 100049
3 福建省厦门第六中学, 厦门 361012
4 中国科学院生态环境研究中心,城市与区域生态国家重点实验室, 北京 100085
微生物与人类关系极为密切。研究证实微生物能调控人体免疫机能、参与新陈代谢、甚至影响个体的情绪和认知行为[1]。随着工业化和城市化的不断推进,大量人口从农村向城市迁移,人们的生活环境和生活方式都发生了明显变化,人体体表和体内的微生物群落结构和多样性也随着外界环境的转变而发生着一系列相应的改变。其中,城市居民由于自身免疫系统失调而引发的多种疾病在很大程度上就是由于其体内微生物群落的变化而引起的[2]。过敏、哮喘、糖尿病、炎症性肠病、以及肥胖和抑郁症等传统上认为与免疫系统无关的疾病在近年来都逐渐被证实与人体内的微生物群落特征有着密切的关系[3]。除了饮食结构、抗生素使用等原因外,生活环境的变化也会改变人群携带的微生物群落。外界环境也因此以微生物为沟通渠道与人群健康建立起间接但关键的联系[4]。
细菌、真菌和病毒等微生物粒子广泛存在于大气中,它们依附在悬浮于空气中的颗粒物上,随着空气流动在水平或垂直方向上运移,对沿途诸多环境过程和人群健康均会造成影响。例如某些微生物会在其代谢过程中产生N2O、NO等污染物,也有些微生物有助于大气污染物的降解[5]。空气微生物对人群健康的威胁主要来源于病原体与人体接触所造成的感染和继发性传播[4]。研究表明,人每天吸入的细菌数量在百万量级[6],快速城市化过程对空气微生物群落的影响必将通过直接或者间接的途径影响人居环境质量及人群健康状况。揭示城市化进程所导致的空气微生物群落的变化及其对人群健康可能造成的影响,首先需要了解空气微生物群落在城市及周边范围内的组成、时空分布特征、变化规律、主要影响因素等,并在此基础上提出可行的景观优化方案,如合理地规划绿地或水体等景观组分来改善城市空气微生物的群落特征,实现对人群健康的保护。
近20年来,城市空气微生物研究呈现快速发展态势。以空气微生物和城市等为关键词在Web of Science上进行检索,有关城市空气微生物的研究论文的数量已经由20世纪80年代末的个位数增加至2019年的234篇。我国对于相关领域的研究始于20世纪70年代,但直到近10年来,国内学者发表的城市空气微生物相关文章数量才迅速增加。虽然目前国内外城市空气微生物研究已经有了一定的进展,但在很多方面的认识仍然非常有限,亟待进一步深入。本文从城市环境中空气微生物的来源、时空分布特征、影响因素、对环境及人群健康的影响等方面对以往研究进行了系统的梳理和总结,明确了当前城市空气微生物研究中的不足以及未来需要开展的方向,以期为构建健康的城市人居环境提供科学参考。
1 空气微生物的来源及其分布特征
空气微生物主要包括细菌、真菌、病毒和少量古生菌,不同环境中的空气微生物组成具有明显差异,但都以细菌和真菌为主[7]。空气中所有生物成分均来自于地表。植被、土壤、水体、动物体内或体表等生物或非生物表面均为空气微生物的来源。微生物以不同方式进入近地表大气,并随着空气运动向周围扩散[4]。Jones和Harrison[8]将微生物释放分为主动和被动两种方式。动物咳嗽或打喷嚏时形成的飞沫中携带有大量细菌(有时还会有部分病毒),这种微生物释放方式被称为主动释放,而通过风力扰动或摩擦触碰等形成的释放则被称为被动释放。释放后的微生物会附着在灰尘粒子、植物孢子上,又或者以细胞群的方式漂浮在大气中[9]。植物表面,尤其是叶片上的微生物的主要来源可以分为两类。第一类是种子和土壤中的微生物随种子萌发和植株生长过程与植物组织形成共生关系,并发展成为依赖于植物种类的特殊微生物群落。如Knief 等[10]发现不同年份收集的一年生草本植物拟南芥(ArabidopsisThaliana)叶片上的甲基杆菌属细菌(Methylobacterium)的群落组成相似。固定的本地微生物来源是形成这种微生物种群特征连续性的重要原因。第二类是大气中的微生物通过植物表面对颗粒物的滞留等方式在叶片等部位进行生存和繁衍。Smets等[11]对位于城市和乡村的常春藤属植物叶片上的微生物进行采集和分析发现,两个区域所采集的叶际微生物种群特征差别明显。城市区域采集的叶际微生物以Hymenobacter和Sphingomonadaceae为主,而乡村区域采集的叶际微生物以Beijerinckia和Methylocystaceae为主。以上研究虽然存在矛盾之处,但也说明植物既是空气微生物的源,也是空气微生物的汇。植物、空气以及土壤等周围环境的共同作用导致了大气微生物的空间异质性。
大气是一种贫营养介质,紫外线辐射强度高,温度、湿度和酸度等属性的变化范围较其他介质也更为剧烈,这导致空气中无法形成稳定的微生物群落,但仍然栖息着丰富的原核和真核生物[12]。微生物对外界环境具有很强的适应能力。Tringe等[13]研究表明空气微生物会通过基因修复和调整新陈代谢等机制来适应大气中恶劣的生存环境。空气微生物是大气生物气溶胶的主要组成成分,因此生物气溶胶有时又被称为微生物气溶胶[12]。微生物气溶胶种类繁多,粒径大小差异显著,其中真菌孢子粒径为1—30 μm,细菌粒径范围为0.25—8 μm,病毒粒径一般小于0.3 μm[8]。微生物粒子的形状、密度以及大小在一定程度上决定了它们在大气中的运动特征。由于微生物粒子个体微小,进入大气后可随空气运动进行长时间、长距离的移动,甚至实现跨大洲运移[14],并对下风向大气微生物群落的构成产生影响[7]。在垂直方向上,微生物主要存在于近地表大气层,但在高达80 km的平流层也有微生物生存[15]。悬浮在大气中的微生物粒子最终会以干、湿沉降的方式回归生物或非生物表面(图1),其中干沉降指大气中的微生物粒子以其自身末速度下降并附着于各类表面的过程,其沉降速度与颗粒直径的平方成正比[16],而湿沉降则是随雨、雪、冰雹等迅速降落至地表的过程[17]。空气微生物随着气流向周围的扩散在相当程度上降低了源景观造成的异质性,也在一定程度上增加了确定空气微生物来源的难度[18—19]。
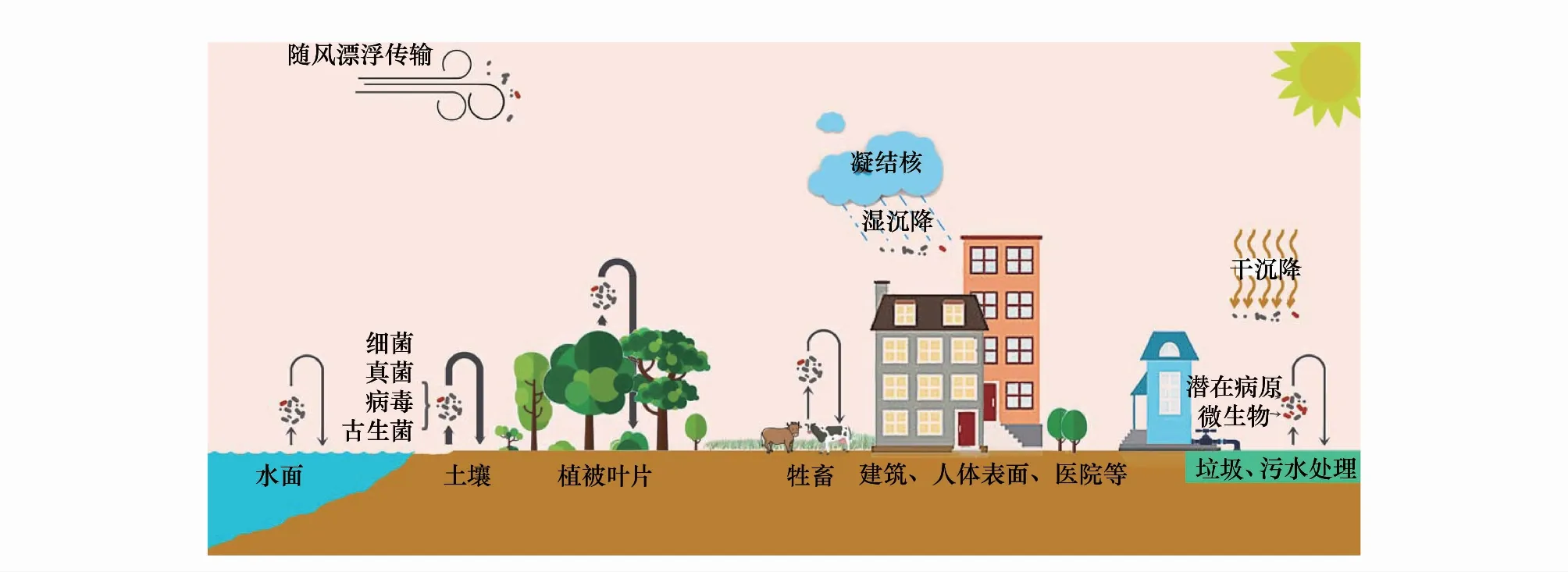
图1 空气微生物的主要来源、种类和潜在影响示意图
城市化过程造成的景观演变导致该区域内的空气微生物的源景观特征明显区别于其他区域。首先,在城市化过程中,大面积原有自然地表被沥青和混凝土为主的不透水面所代替或者切割,自然地表呈现少量化和破碎化,并且城市园林绿化还会引入大量非本地物种。植物表面是空气微生物的重要来源,源景观格局的变化自然会影响到相应微生物的排放特征[20];其次,城市环境污染不仅直接影响人群健康,还可以改变微生物群落特征。土壤是空气微生物的主要来源之一,土壤微生物的数量和活性主要取决于土壤中有机物含量、pH值、水分条件等[21]。然而,机动车尾气排放、化石燃料燃烧、化学用品的大量应用,致使很多城市区域的土壤环境遭受着不同程度的有机物污染[22—25]。土壤有机物污染具有环境持久性和生物累积性,会引起土壤微生物群落结构和多样性发生改变,还会导致病原菌数量和种类的增加[26]。城市大气微生物的其他来源,如水体等,也都受到人类活动不同程度的影响[27—29],从而直接或间接地影响空气微生物群落特征及其生态功能。城市中的污水处理厂和垃圾填埋场也是空气微生物的重要来源[30—31]。污水处理和垃圾填埋等过程均会向大气中释放大量微生物,其中包括了不动杆菌属(Acinetobacter)、假单胞菌属(Pseudomonas)、肠球菌属(Enterococcus)和芽孢杆菌属(Bacillus)等机会性致病菌。这些机会性致病菌随空气流动在一定范围内形成浓度梯度,对该区域内城市居民健康形成潜在威胁。
2 空气微生物群落的时空异质性及其影响因素
2.1 空气微生物群落特征的时间异质性
受多种环境因素影响,空气微生物群落在不同时间尺度上均表现出强烈的不稳定性[32]。例如,Lighthart[33]以不同时间间隔进行空气微生物采样后发现,当样本采集的时间间隔为2 min时,样本中微生物种类的相似度很低,但当采样间隔增加到15 min后,微生物种类的相似性则会大幅增加,其自相关性大于70%。此外,Lighthart[33]还发现,空气微生物的丰度在一天中呈现明显的波动,分别在7:00—8:00、15:00—16:00和18:00—19:00左右出现高峰期,而在21:00到次日5:00,下降至一天中的最低水平。Gusareva等[34]在新加坡对空气微生物群落特征进行了长时间连续监测后发现,微生物群落组成在一天内存在显著变化。该研究所采集的空气样本中的微生物以Ascomycota,Viridiplantae,Proteobacteria,Firmicutes,Cyanobacteria,Actinobacteria和Basidiomycota为主。大多数细菌和真菌在白天的丰度更高,其中Ascomycota在中午时的丰度是夜间的4倍,Proteobacteria和Firmicutes在中午时的丰度是夜间的10倍,只有Basidiomycota在中午时的丰度要明显低于夜间。当把时间尺度由日尺度延伸至月份和季节后发现,空气微生物物种数量在昼夜间的变化幅度要高于月份,而月份间的变化幅度又高于季节[34]。
2.2 空气微生物群落特征的空间异质性
从微生物的组成来看,不同区域空气中的微生物都以细菌和真菌为主,且二者在门的水平上存在较大程度的相似性[35—36]。其中,空气中的细菌主要以变形菌门 (Proteobacteria)、拟杆菌门(Bacteroidetes)、厚壁菌门 (Firmicutes)、蓝藻菌门 (Cyanobacteria)、放线菌门 (Actinobacteria)、酸杆菌门 (Acidobacteria)和异常球菌-栖热菌门 (Deinococcus-Thermus)等为主,真菌则以子囊菌门 (Ascomycota)和担子菌门 (Basidiomycota)为主。Flies等[37]回顾了已发表的关于城市和乡村(郊区)空气微生物群落特征的文章,发现在19项明确区分城乡差别的研究中,多数结果显示在空气微生物的丰度和多样性方面乡村(郊区)明显高于城市。例如,Tanaka等[38]发现郊区所采空气样本中的基因拷贝数是城市区域的1.7倍,空气中的细菌多样性也明显高于城市区域。城市和郊区各有一部分菌种的丰富度较高(Staphylococcus和Propionibacterium的丰度在城市区域较高,Methylobacterium和Sphingomonas的丰度在郊区较高)。从基因水平进行分析发现,城市区域具有更多的人类皮肤相关的细菌种类,而郊区则具有更多的土壤和植物相关的细菌种类。Stewart等[39]也发现城市和郊区在空气微生物群落结构方面具有显著差异。在门水平上,城市空气微生物中的Proteobacteria,Firmicutes,Actinobacteria和Bacteroidetes明显高于郊区,而Cyanobacteria,Tenericutes,Fusobacteria和Deionococcus则明显低于郊区。其溯源分析结果与Tanaka等[38]的研究有所差异,该研究发现城市和郊区分别有相当比例(分别为70.6%和42.9%)的细菌与动物相关。上述研究得出的城市区域空气微生物的丰度和多样性低于乡村或郊区的结论,说明城市区域的微生物群落相似性更高,这可能与城市土地利用方式同质化程度较高有关。在城市化进程中,虽然城市土地被划分成为不同的功能片区,但空气微生物的主要来源(土壤和植物)却在统一的管理措施下呈现出同质化趋势。相似的植物种类搭配和土壤灌溉施肥方式最终形成了相近的空气微生物排放源。然而,需要注意的是,在Flies等[37]所统计的文献中,还有部分文献发现乡村(郊区)的空气微生物丰度和多样性低于城市或者二者之间无显著差异。研究认为这可能与空气微生物群落的高时空变化特征有关,即已有研究中的城市和乡村(郊区)内少量的空气样本可能无法反映当地的空气微生物群落特征,并对城市与乡村(郊区)之间真实差距的客观判断产生影响。城市和乡村(郊区)之间的对比研究仍显不足,还需要在时间和空间分辨率上增加密度,来明确城乡(郊)之间空气微生物群落特征的客观差异和造成差异的原因。从城市的角度来看,在不同城市的空气中,主要微生物种类的占比会有所不同。例如,Liu等[35]对杭州空气样本进行检测后发现,变形菌门占比最高 (32.2%),蓝藻菌门、放线菌门、厚壁菌门和拟杆菌门所占比例分别为18.0%、16.5%、15.5%和11.6%;Li等[40]对厦门空气进行检测后发现最多的四种细菌依次为厚壁菌门、变形菌门、酸杆菌门和放线菌门。城市空气微生物中的细菌在属水平上也具有一定的相似性,例如,Fan等[41]在西安市秋冬季空气中检测到17个属的机会性致病细菌,其中16个属在西班牙马德里市的城市空气中也存在,只有伯克氏菌属 (Burkholderia)没有在马德里市的空气样本中检测到[42]。不同种类微生物的生物属性不同,对环境的适应能力也存在差异,这使得某些环境条件往往可以给特定微生物群落提供更加适宜的生存环境。将城市景观细分为不同的功能区,对比功能区内部空气微生物群落结构特征之间的差异也有少量研究。Hai 等[43]分析了气象因子(空气温度、相对湿度和CO2浓度)对道路、动物园和居住区内部空气中细菌和真菌群落特征的影响,回归分析后发现不同的气象因子对于空气中细菌和真菌的丰度具有不同的贡献度。Li 等[44]按照人为影响程度,对城市内的森林、道路、低密度居住区和高密度居住区进行了空气微生物检测和分析,共检出41种机会性致病菌。在四种采样区内,机会性致病菌丰度最大的区域是道路,最小的区域是森林。为了探明影响机会性致病菌丰度的主要因素,该研究分析了致病菌丰度与人口密度、土壤类型、树种、NDVI等多种因子之间的关系。结果发现机会性致病菌与研究区人口密度具有显著正相关关系,而与NDVI和马尾松覆盖度呈显著负相关关系。Zhai等[36]对在不同场所检测到的空气细菌分析后发现,存在于空气中的细菌种类不仅在城乡环境梯度、城市功能区这种较大空间尺度上存在差异,在更小的空间尺度上也表现出明显的不同。例如,在零售店内检测到的主要细菌种类为Streptophyta,Bacillus,Corynebacteria,Pseudomonas,Acinetobacter,而在住宅检测到的细菌则主要为Sphingomonas,Staphylococcus,Bacillus。因此,将城市、郊区、农村等景观作为整体进行比较,无法揭示景观内部空气微生物的时空分布格局,也无法对导致其时空分布格局变化的主控因子进行详细区分。接下来的研究应首先对研究区进行土地利用类型划分,同时考虑该土地利用类型的存续时间,在此基础上进行空气样本采集并分析微生物的群落特征,才能深入理解导致空气微生物时空变异的原因。
空气微生物群落特征的空间异质性显然会受到附近土地利用的显著影响。郑芷青等[45]对珠三角地区城市空气微生物的研究表明,其多样性具有明显的空间异质性,最高多样性出现在交通枢纽区域,其次是工业区、商业区和居住区,城市绿地内的空气微生物多样性最低。Bowers等[20]研究发现,空气微生物的丰度在不同土地利用类型上保持相对稳定,均为每立方米空气中存在约105—106个微生物,但其群落组成在不同土地利用类型上却存在明显区别。该研究认为土地利用特征(植被覆盖度、管理方式等)对空气微生物群落组成的贡献要大于局地气象条件。土地覆被,尤其是植被,对空气微生物组成具有重要影响。Lymperopoulou等[46]通过对相距50 m的有植被和无植被覆盖的两个区域进行空气微生物采样分析后发现,有植被覆盖区空气中的细菌和真菌数量比无植被覆盖区高2—10倍,并且两个区域的细菌组成也具有明显差别。通过微生物溯源分析发现,有植被覆盖区下风向的空气中50%以上的细菌来源于植物。Mhuireach等[47]比较了美国尤金市5组公园及其附近停车场内空气微生物的群落组成,发现公园内空气微生物的β多样性要高于其附近停车场,采样点周围50 m范围内的绿地覆盖率可以解释空气微生物组成信息的15%。Lin 等[48]对中国台湾省台中市郊区至市中心每隔1 km进行空气真菌采样发现,空气真菌孢子浓度与NDVI呈正相关关系。
气象条件也会影响空气微生物的空间分布格局。微生物可借助风力由生物和非生物表面向大气中释放,导致局部大气中微生物浓度增加,也可以随着空气运动做一定距离的运移,对大气中原有微生物起到稀释作用[49]。微生物在大气中的运移时长受风力和所处大气层位置影响,距离长短不一,最远可达数千公里。因此,源景观在风力作用下会对汇景观上空的空气微生物群落特征产生不同程度影响。例如Barberán等[14]分别在西欧的比利牛斯山和西非的毛里塔尼亚进行了空气、土壤和水汽界面上微生物的采集,发现在撒哈拉沙尘暴发生期间,比利牛斯山脉中的空气微生物沉积样本中可检测到的微生物种群与毛里塔尼亚境内采集的土壤微生物种群具有很高的相似性。多项研究结果显示,空气温度是影响微生物群落特征的主导因素[50—51]。Liu等[52]研究显示,温度和空气微生物多样性呈显著正相关关系。温度的变动也会影响空气微生物的丰度,如Gusareva等[34]研究发现真菌中Ascomycota的相对丰度与温度呈正比,而asidiomycota的相对丰度则与温度呈反比。空气湿度与微生物多样性的关系多呈负相关,这是因为随着空气湿度的增加,大气颗粒物会吸湿增长并导致粒径和重量增加,可加速颗粒物以及附着在颗粒物上的微生物的沉降过程。此外,空气湿度增加还会降低地表微生物向大气释放的速度[53]。气象要素具有强烈的时空异质性,对于微生物释放后的分布格局及其生态功能具有重要影响,是空气微生物研究过程中不可忽略的制约因素。
大气颗粒物,尤其是污染物,与空气微生物的群落结构特征关系密切,可能是造成城乡(郊)空气微生物出现差异的主要原因之一。高敏等[54]在北京的研究发现,生物气溶胶浓度和PM2.5浓度呈负相关。Fan等[41]的研究发现西安市的大气颗粒物(PM10、PM2.5)、SO2、NOX、CO和O3等对潜在病原微生物所占比例的影响要大于相对温度、湿度和风速等气象因素,且不同污染物对病原微生物的影响会随着季节的不同而变化。Liu等[35]在杭州的研究表明,PM10、PM2.5、NO2的浓度和空气中细菌和真菌的群落相似度均呈正相关,而SO2的浓度变化对真菌群落相似度无显著影响。马曼曼等[55]在青岛的研究中发现,由非霾天气到重度雾霾天气的变化过程中,空气中细菌群落的丰富度和多样性均出现下降趋势,不同粒径空气颗粒物上附着的细菌丰度之间差异减小,而复杂度和均匀度的差异增大。Sun等[56]分析了哈尔滨雾霾天气和非雾霾天气对空气中细菌群落结构的影响,结果表明空气污染物给细菌新陈代谢提供了养分(硫酸盐、硝酸盐和铵盐),同时也对细菌细胞造成了破坏并导致更多的细菌死亡。Gao等[57]研究显示空气污染物对细菌群落变化的贡献达到31.9%,并认为空气污染物对细菌群落的构建起到重要作用。然而,也有研究显示大气污染对空气微生物群落特征变化的影响较小。例如,Zhen等[49]发现空气污染物对于空气中细菌群落结构的变化所起作用并不明显。同样,Wei等[58]对雾霾天气和非雾霾天气进行比较后发现,空气中主要细菌丰度并不随空气污染物的浓度变化而出现明显波动。
总体来看,微生物来源、气象条件、人类影响均在塑造微生物群落特征的时空异质性上具有重要作用[59],然而,全面评价城市各类景观组成和结构对空气微生物影响的研究还极为缺乏,各种影响因素对微生物群落特征的作用大小也尚未厘清,以往研究结论还存在诸多矛盾之处。
3 空气微生物的生态与环境效应
3.1 空气微生物的气象效应与生物地球化学循环
微生物在大气中的生存受到气象和环境等诸多因子的影响,同时,空气中的微生物也会通过其自身所具有的气象效应来改变其生存环境内温湿度等物理性质。研究发现,由灰尘和附着其上的生物粒子构成的生物气溶胶可以作为冰核和云凝结核来凝结大气中的水分,影响局地水文循环过程[59—60]。某些微生物细胞的蛋白质结构具有促进或抑制冰晶增长的作用,这种作用可使微生物所处环境维持在一个相对稳定的温湿度状态下[60]。对凝结作用的控制被认为可能是微生物所表现出的一种为改善其生存环境而进化出的功能,这种功能可以改变微生物所处的微环境属性或者微生物的沉降过程。Morris等[61]将空气微生物对水汽的凝结功能放入景观尺度的生态过程中进行分析,发现不同的景观类型会通过近地表能量平衡和水汽蒸散发来影响局地降水过程,从而为土壤和植被中存在的微生物提供了更多生存机会。植被和土壤通过向大气中释放微生物,微生物进入大气后可以通过凝结作用参与水文循环,水文循环又进一步促进微生物在大气和地表之间的交流。微生物因此参与了景观尺度的生态过程,并随着大气的远距离传输,在全球范围内参与更大的气候变化过程。生物气溶胶作为大气气溶胶的重要组成部分,还会通过吸收和散射太阳短波辐射和地面长波辐射来影响地气辐射平衡,同时参与大气物理和化学过程。例如,空气微生物可以以大气颗粒物为载体,通过吸附大气污染物(NOX,VOCs等)、作为氧化剂或还原剂降解或转化污染物、调节大气pH值等参与大气环境的变化[4]。某些好氧微生物还会通过新陈代谢消耗空气中的有机碳、减少自由基来源,由此降低二次空气污染物的产生[62]。
3.2 空气微生物与人群健康
空气微生物可以通过呼吸道和消化道等多种途径进入人体。与空气微生物有关的疾病主要包括传染性疾病、过敏性疾病和癌症等[63]。可引发传染性疾病的微生物主要为细菌和病毒,如结核杆菌(Mycobacteriumtuberculosis)、百日咳杆菌(Bordetellapertussis)、嗜肺军团菌(Legionellapneumophila)、甲型流感病毒、新型冠状病毒等。这些病原体侵入宿主体内后,利用宿主体内的营养物质进行生存和繁殖,并通过呼吸、咳嗽等方式将繁殖后的病原体向周围传播。特定人群对于此类疾病具有更高的致病风险。Baussano等[64]通过meta分析发现,医护人员感染结核病的几率是普通人群的三倍。致敏是空气微生物常见的人群健康效应。许多细菌和真菌均可以分泌过敏原,引发人体过敏反应。然而,也有研究认为人们在儿童时期对空气微生物的暴露可能会对某些过敏性疾病具有保护作用。同时,多项研究证明,随着城市化的进行和人们生活方式的改变,城市居民罹患过敏性疾病的几率明显高于乡村地区[65]。对此,而其中一种解释认为在城市化的过程中,城市区域丧失了一部分环境多样性,并导致环境中微生物多样性的同步降低,环境微生物多样性的降低使人们失去了对个体免疫系统发育的积极影响,由此导致过敏性疾病人数的增加[66]。Alenius等[67]进行了人体免疫机能的城乡空气微生物暴露实验,结果表明乡村空气微生物对人体的早期暴露可以刺激体内调节性T细胞和Th1型辅助细胞的免疫反应,这有利于降低Th2型细胞介导的过敏反应。Roy等[68]研究发现,乡村的居民住房和牲畜棚圈内的空气微生物丰度显著高于城市居民住房,这些高丰度空气微生物群落有利于刺激人体免疫系统释放抗炎性细胞因子。此外,城市大气污染往往高于周围乡村(郊区),而在大气污染条件下,空气微生物和空气污染物对人体的危害还会形成叠加效应[37]。根据以往研究,大气污染对非病原性腐生菌具有灭杀作用,但却可以增加病原菌的相对丰度[52]。空气中的污染物还会对微生物细胞的蛋白质进行转译修饰,增加空气中过敏原和致病原的潜在致病能力[69]。有研究显示,长期暴露于高浓度空气微生物中甚至可以诱发诸如肺癌、胰腺癌和脑癌等重大疾病[70]。例如,Johnson和Choi[71]对屠宰场工人患癌风险和生物气溶胶暴露进行了关联分析,发现这些工人在作业过程中需要暴露于由动物粪便、皮屑、羽毛和各种微生物(细菌、真菌、病毒和内毒素)等组成的高浓度生物气溶胶中,结果显示在控制吸烟的情况下,屠宰场工人罹患肺癌的几率仍然会增加30%左右。
4 研究展望
城市是一类以人类活动为中心的社会-经济-自然复合生态系统。该生态系统与自然状态下的生态系统以及接近自然状态的乡村生态系统在多个方面存在显著差异。相对于自然生态系统而言,人口高度聚集的城市区域面向居民的生产生活需求分化出多种功能片区。不同种类的功能区具有不同的土地利用方式、植被覆盖率以及人为干扰强度等,而这些方面的差异都会反映在主要来源于地表的空气微生物群落特征上。然而,目前对于城市和乡村以及自然生态系统内空气微生物的研究尚缺乏系统性,对于按照人为干扰进行分类的城市-乡村(郊区)-自然生态系统内的空气微生物群落特征差异的讨论还存在争议,对于城市内部不同功能区内空气微生物群落特征的研究更加少见。随着城市化进程的不断推进,高度聚集的城市人口对于空气微生物的敏感程度正在逐步升高,城市居民罹患免疫系统疾病的概率呈增加趋势。部分学者将这种现象归因于城市居民生活方式的改变而导致的居民免疫系统与城市空气微生物环境的不相适应。总之,尽快对城市内部不同功能区、城市-乡村(郊区)-自然生态系统内的空气微生物群落特征从时空两个层面进行深入探索,对于理解空气微生物的生态系统发展过程以及人群健康预测预警均具有重要意义。
据估算,全球每年由地表向大气中排放的生物气溶胶量约占大气气溶胶浓度的30%左右[72]。相对于其他类型气溶胶,针对生物气溶胶的研究还远远不够[73]。同时,城市空气微生物的研究在诸多方面还存在很多尚未解决的问题,比如空气微生物的本地源和外来源的占比、空气污染对于空气微生物群落特征的影响程度及机制、空气微生物在成云和降水过程中的实际作用等。深入理解城市空气微生物的生态与环境效应,不仅需要多个学科交叉集成,还要统筹宏观和微观的研究方法进行深入探索。
4.1 多尺度景观格局与空气微生物群落特征的时空异质性
空气微生物的隐蔽性强,容易成为人群健康的潜在隐患,而消除隐患的前提在于深入了解存在于空气中的微生物与周围环境的关系,包括微生物的来源及其传播途径。了解空气微生物的来源有助于积极地对病原菌、病毒等进行源头控制,明确其传播途径则有利于进行固定方向的扩散或者切断扩散路径以降低关键点的病原体浓度[74]。气象模式在空气污染、热岛效应等涉及空气流动的研究上均起到了重要作用,但对空气微生物的运移及其环境效应等方面的模拟还十分缺乏。此外,分析空气微生物和周围环境的关系不能局限于少数几种土地利用类型,也不能局限于街区尺度,需要在城市或者区域等较大尺度上才可能深入理解微生物的来源及其扩散过程,这涉及到城市或区域景观格局与微生物生态过程的相互作用。探索景观格局与微生物群落特征之间的关系,并在城市尺度上划分人群对病原体的暴露等级,为环境风险管理提供支撑,是景观生态学理论在微生物生态领域的实践,有助于拓展景观生态学的应用外沿,也有助于深化空间格局与生态过程相互作用的内涵。
由于空气微生物群落特征受到温湿度、太阳辐射强度、海拔、采样点用地类型等多方面因素的影响,导致其表现出很强的时空异质性,使得不同研究的观测结果之间往往出现较大差异,外界环境的持续变化也使得短时间定点采样无法反映空气微生物群落特征的变化规律。城市化是区域土地利用变化的主导过程。城市功能区的差异使得城市景观高度斑块化,不同类别斑块的植被覆盖程度、生物多样性、人类活动强度往往区别较大,主要来源于土壤和植被的微生物群落的特征也会表现出显著差异。同时,随着城市更新的进行,斑块内部的生态系统还会受到进一步的扰动,使其在受干扰程度上有别于同属一个功能区的其他斑块。因此,可以按功能和年代将城市景观划分为不同梯度的斑块系统,并在此基础上进行空气微生物群落特征的研究,这种方法有望对城市微生物的生态过程进行系统的解析。
4.2 城市系统多属性特征与空气微生物群落结构刻画
作为人地耦合系统不可或缺的一部分,空气微生物群落特征和环境效应的研究还应从生态系统的视角进行,在城市区域更需要考虑社会、经济和自然因素的复合影响,才能全面了解城市空气微生物群落特征的动态变化规律,解析城市化过程对空气微生物的影响。2004年,方治国等[32]提出城市空气微生物的研究工作大部分是单一研究城市空气微生物的种类、数量及浓度的时间和空间变化,缺乏从生态系统的角度来研究城市空气微生物。时至今日,这种状况并未明显改变。当前研究仍缺乏对城市生态系统的社会、经济、自然组成因素对空气微生物的直接或间接影响及其作用过程的全面考虑,对城市绿地的刻画也仅限于使用绿地覆盖率和NDVI等有限的指标。城市生态系统是人类活动影响下的复合生态系统,其景观多样性和复杂度都显著高于其他生态系统,分析特定样点附近空气微生物群落的组成和多样性对描述整个城市生态系统中空气微生物群落特征的时空异质性贡献有限,需要建立采样点周围一定范围内的景观组成及结构以及社会经济状况与空气微生物群落特征之间的联系。城市景观高度复杂、城市生态过程杂乱多样,利用一种指数来准确表述景观格局与不同生态过程之间的关系并不现实,还需要在深入理解生态过程的基础上,有效组合已有指数或者开发新指数才能更有效地刻画二者之间关系。
4.3 城市空气微生物的健康风险评估与城市规划实践
高密度居住环境给传染性病快速扩散提供了便利条件,这使得此类疾病在城市中的传播速度要远高于乡村(郊区)。将地理空间和城市分区信息纳入到城市公共卫生突发事件的管控中,从十九世纪中期英国伦敦地区爆发霍乱疫情时便有了先例[75]。人们逐渐意识到公共卫生对于城市居民生活和生产的意义,也认识到城市规划对于良好的公共卫生环境的创建所具有的重要性。但随着城市化的不断推进和城市人口的持续增加,突发性传染病疫情仍时有发生,而且每次发生都对城市居民健康和社会经济稳定产生很大冲击。中国香港政府在2003年SARS疫情得到控制后提出对于大型规划和发展项目进行空气流动评估,开设通风廊道,通过空气流动降低区域内病原微生物的浓度,同时提出建设更多绿地和休闲用地。在中国的其他一些城市,SARS疫情的爆发也使城市规划工作者对于城市发展产生了一些思考[76]。城市规划实践是在城市居民健康风险评估的基础上开展的,而针对病原微生物的居民健康风险评估需要对空气微生物群落特征具有全面了解,主要包括三个方面:(1)病原微生物的种属区分、传播途径和致病特征;(2)宿主的生理特征、营养状况和社会属性等;(3)病原微生物的时空异质性、浓度水平、扩散传播模式、生长繁殖特征及其环境影响因素[77]。其中,包括景观生态学、生态系统生态学、城市复合生态系统、遥感和地理信息系统等理论和方法在健康风险评估过程中将起到重要作用。将宏观与微观相结合,厘清城市空气微生物群落特征,并以此为基础参与到公共卫生管理和城市规划实践中,对于快速城市化背景下的居民生活质量的提升、心理和生理健康的维护具有重要意义。
学术信息与动态
