粳稻种质资源稻瘟病抗性及抗性基因分析
2022-04-06邹拓耿雷跃张薇杜琪孟令启张启星
邹拓 耿雷跃 张薇 杜琪 孟令启 张启星
(1 河北省农林科学院滨海农业研究所,河北 唐山 063299;2 唐山市水稻育种重点实验室,河北 唐山 063000;*第一/通讯作者:549282738@qq.com)
稻瘟病是水稻的主要病害之一,由真菌病原体(Magnaporthe oryzae)引起。1919年在印度首次发现稻瘟病,造成当年水稻减产4%左右[1]。在随后的稻瘟病发生过程中,仅仅在印度东部地区就造成水稻减产50%左右[2-3]。稻瘟病菌具有多样性,以致一个抗病品种在生产上使用3~5年就丧失抗性[4-6]。有研究表明,全球每年由于稻瘟病造成的水稻减产量可以满足6 000 万人口的粮食需求,导致的直接经济损失高达50 亿美元[7-8]。因此,筛选并应用新的抗病种质资源,有助于培育广谱、持久的抗稻瘟病新品种。
近年来,随着分子生物学的发展,越来越多的稻瘟病抗性基因被定位,在已定位的约100个稻瘟病抗性基因中有28个被成功克隆[9-10]。本研究中,利用Pi37、Pib、Pi9、Pigm、Pid3、Piz、Pi36、Pi5、Pia、Pikh、Pik 和 Pita基因的特异性分子标记对参试粳稻资源进行抗病基因型鉴定。其中,Pib 来自品种BL1,对日本大多数稻瘟病菌小种有抗性,对中国的菌株ZB13 和ZC15 也表现抗病反应[11];Pita 对稻瘟病菌菌株 ZN57 和 ZN61 表现抗病[12];Pia 对江苏省生理小种表现为强抗性[13]。
本研究中利用现有的粳稻资源,通过表型评价与基因型鉴定相结合的方式,分析基因型与其抗病反应的关系,找到有效的抗病基因或聚合基因型,筛选到对本地区稻瘟病小种具有较强抗性的种质资源,对今后抗稻瘟病水稻育种具有重要意义。
1 材料与方法
1.1 试验材料
共用到157 份粳稻材料,具体信息如表1。2019年4月至10月种植于河北省农林科学院滨海农业研究所试验基地。

表1 粳稻资源信息
1.2 实验方法
1.2.1 抗病基因分子检测
水稻分蘖高峰期取叶片样品,存放至-20℃冰箱。水稻基因组DNA 的提取按上海植物生理研究所的TPS 抽提法稍加修改。Pi37、Pib、Pi9、Pigm、Pid3、Piz、Pi36、Pi5、Pia、Pikh、Pik 和 Pita 等 12个基因的引物序列及扩增产物信息如表2,引物由北京擎科生物公司代为合成。PCR 的反应体系总体积 20 μl,其中包括0.2 μl dNTP(10 mM),2.0 μl 10×PCR buffer(含 15 mM MgCl2),引物各 1.5 μl ,0.1 μl Taq 酶(5 U/μl),2 μl 材料基因组DNA,其余体积量由ddH2O 补齐。PCR 反应程序:94℃预变性 5 min;94℃变性 45 s,55℃退火 45 s,72℃延伸45 s;72℃,延伸5 min(括号部分为35个循环)。待温度降至25℃,移至4℃冰箱待用。利用1%的琼脂糖凝胶电泳进行扩增片段的分离,在凝胶成像系统下拍照读带。
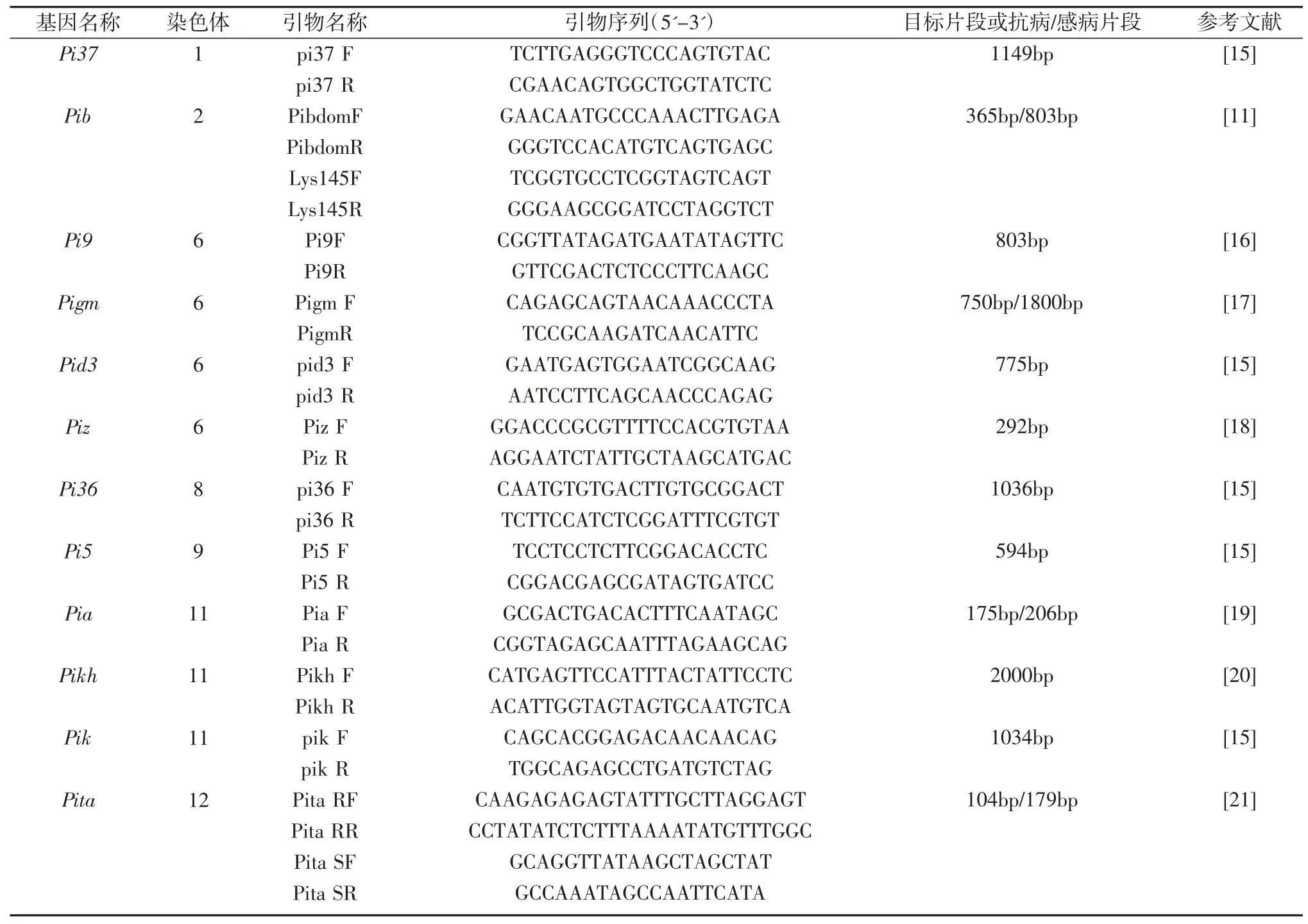
表2 抗稻瘟病基因分子标记引物及扩增片段信息
1.2.2 田间诱发稻瘟病鉴定
供试的 157 份材料于 4月20日播种,5月30日移栽,行株距20 cm×10 cm,每间隔10 份材料种植1 份丽江新团黑谷并在鉴定圃周围种植1 圈丽江新团黑谷。鉴定圃内防虫不治病,增施氮肥,提高田间湿度,其余管理措施同大田。当感病品种丽江新团黑谷发病后确定稻瘟病诱发有效。7月26日至31日进行田间叶瘟发病调查,8月20日开始调查穗瘟。调查方法及病级计算参照NY/T 2646-2014[14]。
2 结果与分析
2.1 粳稻资源田间感病情况
据田间调查结果,在157 份材料中,表现为抗的2份、占比1.27%,中抗70 份、占比 44.59%,中感64 份、占比40.76%,感及高感共21 份、占比13.38%。可见,大多数种质材料抗病综合指数分布在中抗及中感水平间,表现为抗、感及高感的材料较少。参试材料中,叶瘟平均病级4.00,变异系数40.00%;穗瘟平均病级4.23,变异系数50.59%。
2.2 12个抗病基因的检测结果
为明确不同基因型对本地区稻瘟病抗性表现情况,选择抗性表现最好的和抗性表现最差的材料各20份,进行抗稻瘟病基因分子检测。通过比较表现极端的材料,找到材料中基因数量和基因型的差异与抗病性的关系。从图1 可见,检出频率在15 次以上的有8个基因,其中Pid3 和Piz 基因检出频率最高,分别检出40 次和34 次;检出频率在10 次以下的有4个基因,其中,Pi9 未检出。
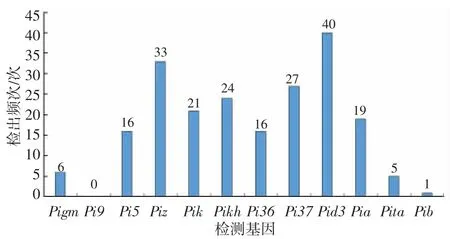
图1 12个抗病基因的检测情况
从表3 可见,40 份材料中,平均含有抗性基因5.2个;抗稻瘟病综合指数表现为抗的材料中含有的抗病基因为5~8个,表现为感病的材料中,含有的抗病基因为2~5个。其中,ZY29 具有抗性基因最多,包含Pi5、Piz、Pik、Pikh、Pi37、Pid3、Pia、Pita 等 8个基因,抗病综合指数为2.5,表现为中抗。ZY115 具有抗性基因数量最少,仅含Pi36、Pid3,抗病综合指数为8.3,表现为高感。
2.3 材料抗性与基因型及基因数量的关系
通过对比抗病材料与感病材料的基因型看出(表3、图2),抗病材料与感病材料相比,大多具有Pigm(5∶2)、Pia(18∶0)、Pita(5∶0)Pik(17∶4)、Pi5(12∶4)这 5个基因,括号中数字为具有此种基因抗病材料与感病材料的数量。由此可见,这5个基因在本地区对稻瘟病的抗病性起到一定作用。利用SPSS 对抗病基因数量与叶瘟病级进行相关性分析,结果(表4)表明,材料中的抗病基因数量与稻瘟病综合指数的相关系数为-0.844,在P=0.01 水平上达到负显著性相关。可见,含有更多抗性基因的水稻材料具有更好的稻瘟病抗性。
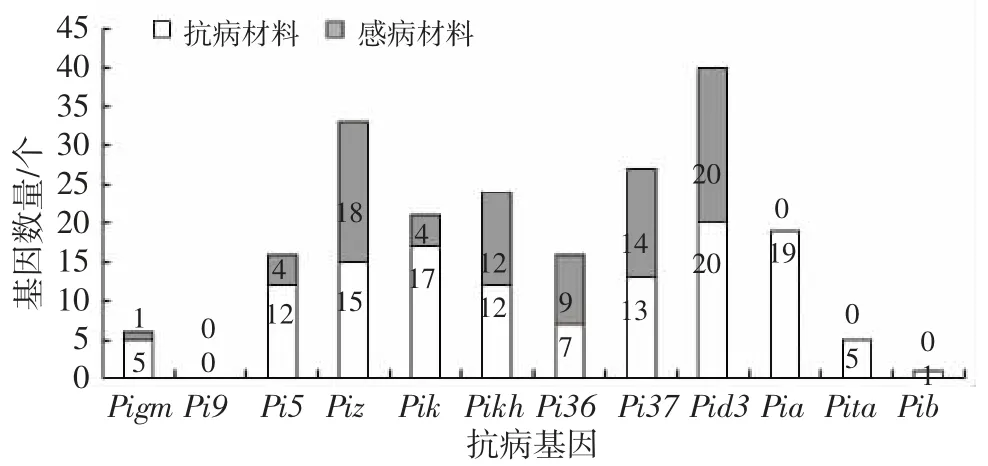
图2 抗病材料与感病材料在基因型上的差异
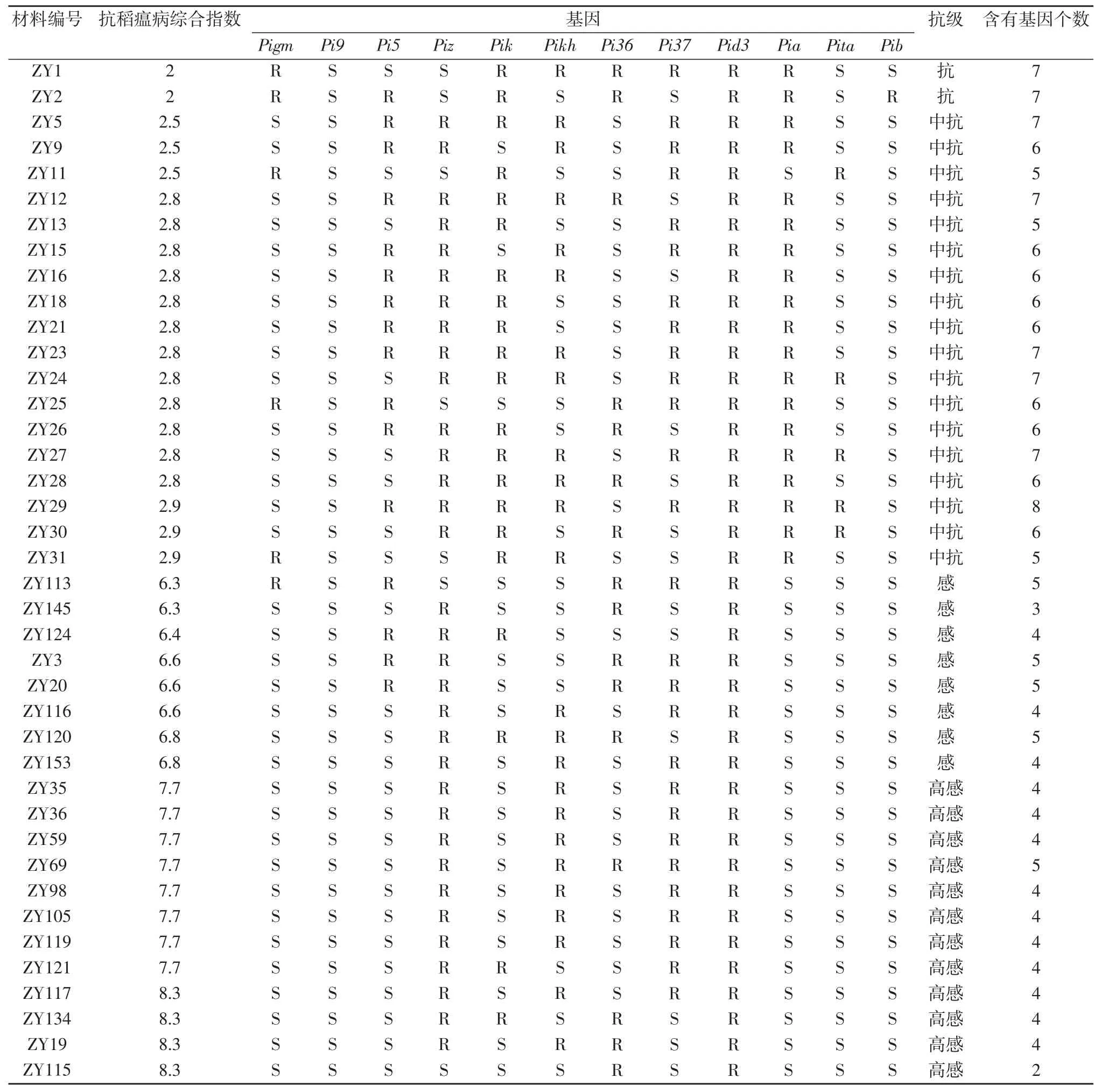
表3 40 份材料的基因型及抗稻瘟病综合指数
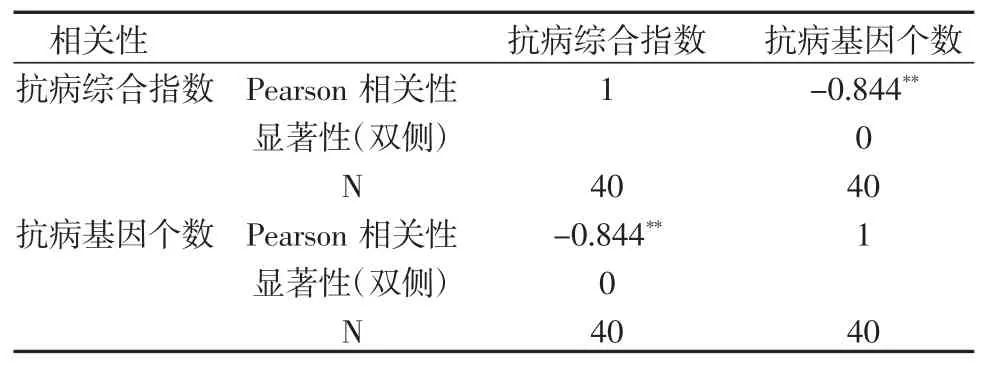
表4 抗病综合指数与抗病基因个数的相关性分析
2.4 种植资源在抗病育种上的应用
从表 5 可见,ZY2、ZY13、ZY15 和 ZY21 等 4个种质资源抗病性好,在株高、穗颈长、产量等性状上能够达到作为育种亲本的要求。因此,育种者可利用这些材料通过分子标记辅助选择与常规育种手段相结合的方式,快速、准确的选育聚合多个抗稻瘟病基因的新品种。
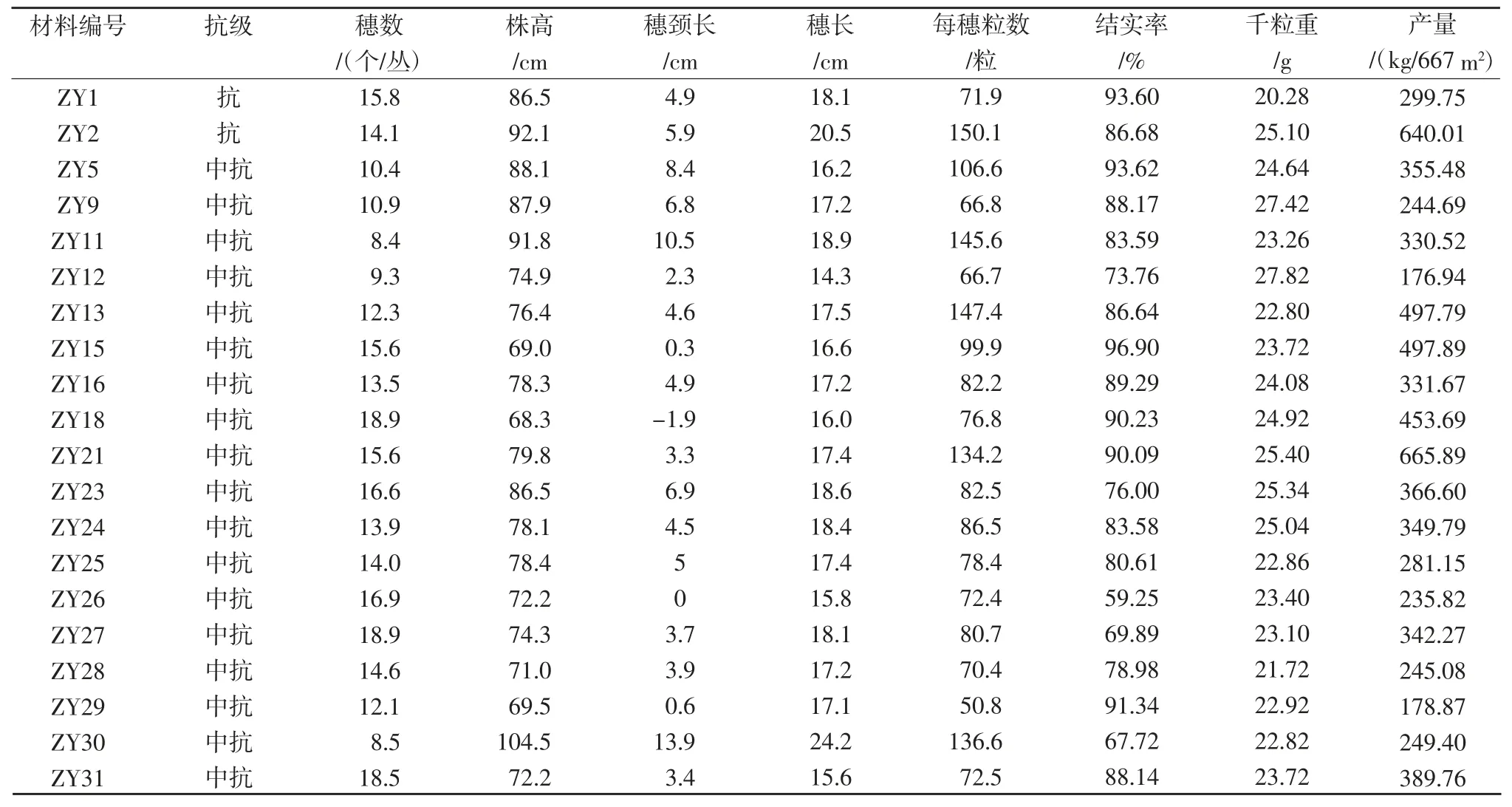
表5 抗病材料的农艺及产量性状
3 讨论
前人研究中,少有利用粳稻资源在冀东地区做稻瘟病抗性评价,而不同稻区稻瘟病生理小种自身也有高度的变异性,气候条件也是影响稻瘟病发病的主要因素[22]。另外,病圃田间的诱发致病的生理小种是未知的混合小种, 而且不同位置的侵染菌种也可能不尽相同,加上小气候差异等因素,所以田间诱发鉴定比人工接种鉴定的可靠性差些[23]。因此,本课题组利用157 份粳稻资源在河北省农林科学院滨海农业研究所试验基地进行田间感病试验,重点结合本地区生态环境及稻瘟病生理小种评价材料的抗病性,再结合分子标记对抗病基因鉴定,挖掘在冀东地区表现高抗性抗病基因,为冀东地区抗病育种提供有效的基因供体。
本次用到的12个抗稻瘟病基因分布在第1、第2、第 6、第 8、第 9、第 11 和第 12 染色体上,全部已经克隆,效应及位点比较明确[9-10]。试验中,用到的标记大多为功能标记。目前定位到的抗性基因除以上7 条染色体外,还存在于第4、第5、第7 和第10 染色体上,但其定位基因较少,未有基因簇的出现[24]。因此,本次选择的基因具有一定的代表性和准确性。
本研究中,通过对病级和抗病基因数量进行相关性分析,得到病级与基因数量成负向极显著,因此聚合更多的抗性基因会提高品种的抗病性,这与张羽提出的稻瘟病抗性也属于数量性状,随着抗性基因数目的增多,品种的抗病性呈上升趋势一致[25]。
4 结论
对157份粳稻资源进行稻瘟病抗性自然诱发鉴定,对稻瘟病表现为抗的有2 份、表现为中抗的有70份。结合稻瘟病基因检测,对比抗病材料与感病材料基因型得到 Pigm、Pia、Pita、Pik、Pi5 等 5个对冀东地区稻瘟病有明显抗病作用的抗性基因,且聚合的抗病基因越多,材料的抗病能力越高。进一步结合农艺及产量性状得到 ZY2、ZY13、ZY15 和 ZY21 这 4个种质资源可作为抗稻瘟病育种中的抗病亲本。
