牙形刺氧同位素古温度计:研究进展与展望
2022-04-02刘康周锡强江茂生
刘康,周锡强,江茂生
1.中国科学院地质与地球物理研究所,中国科学院新生代地质与环境重点实验室,北京 100029 2.中国科学院地球科学研究院,北京 100029 3.中国科学院大学地球与行星科学学院,北京 100049
0 引言

牙形刺在寒武系至三叠系海相地层里分布广泛、种类繁多、特征显著、演化迅速,是古生代地层划分与对比的重要标准化石[26-29]。牙形刺主要由磷灰石(即磷酸钙)有序排列而成,结构致密,能够较好抵抗后期成岩作用改造,化学稳定性好[30-31]。重要的是,磷酸盐氧同位素对环境水体温度变化比较敏感[15,32-33],且易于测量,具备开发为古温度计的必要条件。21 世纪以来,随着地球化学分析测试方法与技术的快速发展,研究人员持续开发、改进和优化牙形刺氧同位素古温度计[31,34-37],并取得重要进展和认识。例如,通过激光剥蚀电感耦合等离子体质谱仪(LAICP-MS)研究揭示,牙形刺不同组织部位抵抗成岩作用改造的能力各异[38];通过透射电镜研究揭示,单颗粒牙形刺内部不同组织部位在晶体大小、形状和晶间(内)孔隙等微观结构上具有差异性[39];通过二次离子质谱仪(SIMS)对牙形刺进行原位微区氧同位素分析,可显著提升古海水温度重建的质量和时间分辨率[35,40]。目前,牙形刺氧同位素已逐渐成为重建深时海水温度的重要手段,得到快速发展和广泛应用,并极大促进了我们对深时地球环境与生物演化的认识。
在此背景下,本文对牙形刺基本特征、磷酸盐氧同位素古温度计原理、分析测试方法与技术,以及古海水温度定量重建影响因素等方面,进行了简要回顾和介绍。同时,进一步讨论了牙形刺氧同位素古温度计的应用现状,并展望其未来发展,以期推动相关研究。
1 牙形刺基本特征
1856 年,俄国学者Pander 在下奥陶统含海绿石砂岩里首次发现了多种呈单锥状或锥齿状的微体生物结构,并创立了“Conodont”一词[41],即牙形刺(或称牙形石)。牙形刺个体微小,长约0.1~3 mm(表1)。成分上,牙形刺主要成分为细晶碳氟磷灰石,可表达为化学式Ca5Na0.14(PO4)3.01(CO3)0.16F0.73(H2O)0.85[42]。形貌上,牙形刺通常呈琥珀色、浅黄色或灰黑色,并可细分为单锥、复合和平台型三种形态类型。结构上,牙形刺可分为基腔(basal body)和齿冠(主齿)(crown)两部分[43],其中齿冠可再细分为半透明的玻璃质(hyaline)和不透明的瓷白质(albid)两部分(图1)。生物学上,牙形刺可能属于底栖或分层游泳无颌类脊椎动物的滤食器官部位[44-48],在浅海陆架至深水盆地环境广泛分布,并常见于碳酸盐岩地层。据统计,牙形刺最早出现于早寒武世,并在早—中奥陶世(约140 属种)和中泥盆世(约160 属种)分别经历了两次辐射高峰,自早石炭世开始逐渐衰减,至晚古生代冰期发生骤减(少于40 属种),最终于三叠纪末期绝灭[44,49]。

图1 牙形刺的形态和结构特征(a,c)臀板刺属,(b,d)镰刺属,样品来自塔里木盆地柯坪地区奥陶系碳酸盐岩。图中A指示瓷白质齿冠,H指示玻璃质齿冠,B指示基腔Fig.1 Morphological and structural features of conodonts(a,c)Pygodus sp.,(b,d)Drepanodus sp.conodonts,which were collected from Ordovician marine carbonates in the Kalpin area,Tarim Basin.A.albid crown;H.hyaline crown;B.basal body

表1 牙形刺的基本特征Table 1 The main characteristics of conodonts
牙形刺中氧的赋存状态及稳定性,对于氧同位素古温度计开发和应用至关重要。理论上,牙形刺中约90%~95%的氧主要赋存于PO43-[50],其余少量赋存于CO2-3和H2O(表1)。其中,CO2-3和H2O易受成岩作用的影响,导致其氧同位素组成发生变化。相比之下,PO3-4在低温环境下具有很强的化学稳定性,能够较好地保留初始氧同位素信号[17,51]。此外,牙形刺磷灰石矿物晶格中的Ca2+、OH-和PO3-4可被其他离子类质同相替代[52],导致其含有一定量的Mg、Sr、Nd、REE 等微量元素杂质,可能会显著降低牙形刺的化学稳定性[42,53]。另一方面,牙形刺的基腔和齿冠存在一定的微观结构和化学成分差异,在遭受机械剥蚀或成岩作用改造时,二者稳定性有所不同。牙形刺基腔部位的磷灰石晶体粒度较细,呈球状或葡萄状集合体,排列不规则,微量元素杂质及有机质含量较高,结构致密性和耐酸性不足,稳定性较差;玻璃质齿冠由细长磷灰石晶体整齐排列而成,具有纹层结构和大量纳米级孔隙,杂质及有机质含量较低,稳定性较好;瓷白质齿冠由粗粒(可达数百微米)磷灰石晶体排列而成,晶间孔隙呈网格状,连通性和渗透性差,微量元素杂质及有机质含量最低,结构致密性和耐酸性强,稳定性相对最好[38-39,54-55]。因此,牙形刺瓷白质齿冠部位的磷酸盐组分,目前普遍作为氧同位素古温度重建的优选对象。
2 牙形刺氧同位素古温度计原理
由于生物矿化成因的磷酸盐矿物具有结构和化学稳定性的优势,研究人员不断探索和开发其氧同位素古温度计,使之日益成熟[32-33,37,56]。理论上,水体溶解态磷酸盐氧同位素组成由水体温度及水分子氧同位素值共同决定[15]:温度越高,溶解态磷酸盐和水分子间的氧同位素分馏系数越小,氧同位素组成越偏轻[57]。生物利用海水磷酸盐矿化生成磷灰石,因此可记录同时期海水温度。需要强调的是,在生物矿化过程中,牙形刺氧同位素组成与同时期海水中水分子在何种程度达到热力学平衡分馏,是否存在同牙形动物生命活动相关的动力学分馏(即生命效应,vital effects),仍无定论[25]。目前,牙形刺作为已绝灭生物的磷酸盐质器官,通常通过借鉴基于现生生物所标定的磷酸盐氧同位素—温度经验公式,定量重建古海水温度。研究人员基于特定生物及其生存的一系列水体温度条件(T),测量发现生物矿化成因磷酸盐矿物(δ18Ophosphate)及周围环境水体(δ18Ow)的氧同位素组成通常呈现较好的线性关系,因此采用数学方法进行线性拟合,即可获得温度T 关于Δ(Δ=δ18Ophosphate-δ18Ow)的经验公式。在此背景下,Longinelliet al.[56]针对含磷酸盐的藤壶和软体动物壳,Kolodnyet al.[32]针对磷酸盐质鱼齿和鱼骨,Pucéatet al.[33]针对人工饲养的海鲤牙齿等,先后开展了一系列研究,采用不同的样品处理手段和磷酸盐标样参考值,依次建立了多种磷酸盐氧同位素—温度经验公式(表2),并被用于牙形刺氧同位素古温度定量重建。

表2 常用的磷酸盐氧同位素—温度经验公式Table 2 Empirical formulas for paleotemperature reconstruction based on phosphate oxygen isotopes
3 牙形刺氧同位素测试方法与技术
牙形刺氧同位素分析测试方法经过多年的探索和开发,不断朝着低样品量、快速高效、高精准度、高分辨率及原位微区的测试方向发展。牙形刺通常以微小个体弥散分布于沉积岩中,丰度很低,通常需要通过物理和化学手段对其进行分选和提取,以便进行氧同位素等各类分析测试。对于碳酸盐岩样品,通常采用醋酸溶解法,依次进行碎样、酸溶、过筛、重液分离和镜下挑选等流程,实现对牙形刺的分离与收集(图2)。对于页岩样品,碎样后,通常采用直接水煮方式将页岩分解成泥浆,再进行过筛将牙形刺牙形刺氧同位素古温度计核心在于准确测量其矿物晶格中PO3-4组分的氧同位素组成,并尽可能消除CO2-3、H2O和有机质的干扰。因此,研究人员先后开发了一系列磷酸盐氧同位素分析测试方法,并应用于牙形刺氧同位素古温度计研究。20 世纪90 年代之前,相关分析方法主要是在酸溶的基础上,将PO3-4特效转化为BiPO4,以测试其氧同位素组成;但是BiPO4具有吸水性,导致其氧同位素组成稳定性较差,且方法比较耗时、对样品需求量大[34,61]。因此,Crowsonet al.[62]对实验流程进行了优化,将PO3-4转化为不具吸水性且易于制备的Ag3PO4,大幅提高了分析测试效率。在此基础上,研究人员针对Ag3PO4氧同位素建立了传统氟化法、高温石墨热还原法、负离子热电离质谱法等多种分析测试方法[30,34,63-65]。然而,这些方法一定程度上具有样品需求量大、费时费力费钱、测试精度有限等缺点,逐渐走向冷落。
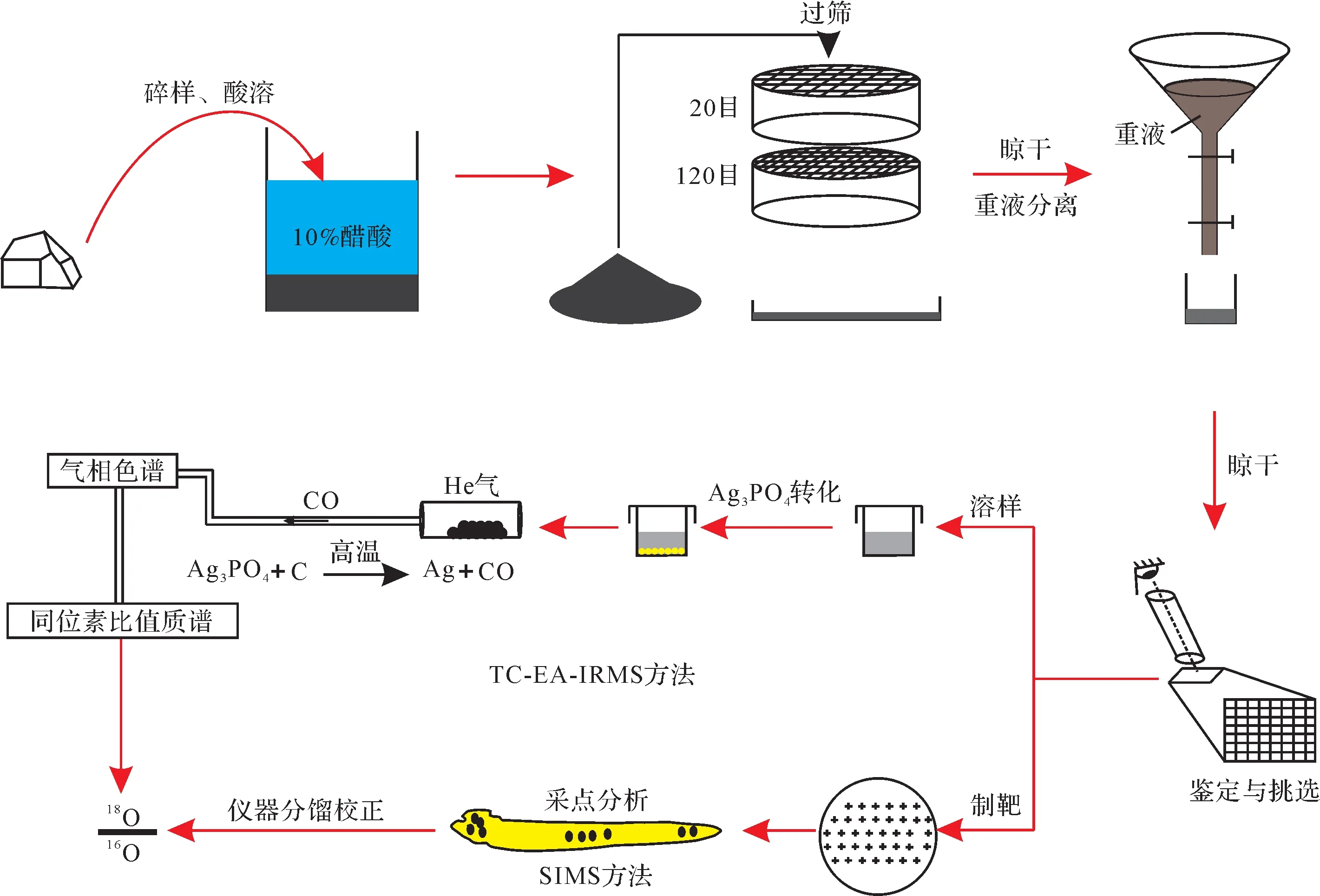
图2 基于TC-EA-IRMS 和SIMS 方法的牙形刺氧同位素分析测试流程示意图Fig.2 Analytical workflow diagram of conodont oxygen isotopes through thermal conversion-elemental analyzer-isotope ratio mass spectrometry (TC-EA-IRMS) and secondary ion mass spectrometry (SIMS) methods
近年,热转换元素分析—气体同位素质谱法(TC-EA-IRMS 方法)逐渐成为牙形刺氧同位素测试的常用方法。该方法首先将牙形刺进行酸溶,释放PO3-4,然后将其转化为Ag3PO4沉淀,再在高温反应炉中被石墨还原生成CO气体,通过连续流质谱仪测量获得氧同位素值[66-70]。此方法能避免牙形刺中CO2-3、H2O 和有机质等组分对PO3-4氧同位素的干扰,而且对样品的需求量较小(0.3~2 mg),分析测试相对快速便捷,测试精度通常可达到约0.20‰(1σ)[20,58,60,71-72]。然而,TC-EA-IRMS 方法可能同时包含不同生物习性、不同成岩作用改造程度的牙形刺属种,导致获取混合的氧同位素信号[21,73]。此外,相关分析流程中磷酸盐的回收率、Ag3PO4沉淀纯度、及酸溶过程中潜在的氧同位素分馏或交换反应等因素,均可能不同程度地影响分析测试精度[30,67,74]。例如,Ag3PO4沉淀过程中容易混入Ag2O 杂质,可干扰磷酸盐组分氧同位素组成的测定[66,70]。近期,Pederzaniet al.[75]提出通过离子交换树脂对样品酸溶后产生的PO3-4依次进行选择性吸附和解吸,并且采用缓慢沉淀Ag3PO4晶体的方式,可提高磷酸盐氧同位素分析测试的精度。
目前诸多研究发现,牙形刺不同属种或单颗粒不同部位均可存在一定程度的组构和组分差异,导致具有非均一的氧同位素组成特征[36,38]。因此,针对特定属种牙形刺的特定部位,高效而精确提取磷酸盐组分原生氧同位素信号,是牙形刺氧同位素古温度计应用的关键技术。随着原位微区分析测试技术与方法的进步,LA-ICP-MS[76]、SIMS[35,40,77-79]等原位微区分析技术已不断应用于牙形刺氧同位素分析。LA-ICP-MS 方法由于激光束斑直径(超过100 μm)较大、空间分辨率较低,难以在单颗牙形刺内部多次采点分析,制约了分析测试数据的精度[76]。相比之下,SIMS方法的离子束斑较小(10 μm)、空间分辨率高,可对单颗牙形刺的不同部位进行高灵敏度的氧同位素分析,同时具有测试快捷、样品需求量小(1~3 ng)、检出限低等优势,逐渐成为分析测试的优选手段。SIMS 方法首先筛选特定属种牙形刺进行制靶,然后利用Cs+一次离子束轰击样品表面,生成的二次离子进入质谱仪分析18O∕16O 比值[80-81],氧同位素测试精度可达0.20‰~0.28‰(1σ)[35,82]。理论上,SIMS 分析方法无法有效区分牙形刺中各类含氧组分(PO3-4、CO2-3、H2O)的来源,但是对比测试显示,SIMS方法和TC-EA-IRMS方法的牙形刺氧同位素测试结果非常接近[36],或者前者相对于后者平均偏高约0.5‰(VSMOW)[40],具有可靠的测试效果。需要指出的是,SIMS 方法对牙形刺制靶质量、采点位置、标样可靠性、仪器系统误差和工作环境等有严格要求,否则一定程度上将影响氧同位素分析测试结果。例如,Sunet al.[83]发现Durango 磷灰石标样颗粒内部δ18O值变化可达0.7‰~2‰(VSMOW),其氧同位素组成非均一性必然会对氧同位素值测试结果标定带来一定的影响。
4 牙形刺古海水温度重建的影响因素
牙形刺氧同位素古温度计基于生物矿化成因磷酸盐组分与同时期海水氧同位素组成,通过氧同位素—温度经验公式(表2),定量重建古海水温度[32-33,56]。事实上,除了分析测试方法相关的误差之外,牙形动物的生物习性、牙形刺成岩作用、数值量化计算等因素,均可能不同程度地影响该古温度计的精准度及其地质意义。
4.1 生物习性
牙形刺氧同位素古海水温度重建,必须考虑牙形动物的生物相或者生物习性[84-85]。牙形动物的生物相是指具有一定沉积环境和生态特征的牙形刺组合,常依据丰度最高的牙形刺属种名称进行命名[86]。例如,Rasmussenet al.[87]基于多元统计学分析,将波罗的陆块中奥陶世牙形刺生物相按照由浅至深的水深顺序依次划分为Baltoniodus-Microzarkodina生物相(内陆架)、Periodon生物相(陆架边缘)和Protopanderodus生物相(远端陆架)。其中,陆架边缘Periodon生物相划分的合理性可得到牙形刺氧同位素证据的支持[85]。由于海水温度存在季节、水深和纬度差异,牙形动物不同属种或底栖[88]、或浮游生活在不同纬度、自表层至数百米不同水深的海洋环境[60,89-90],其生物习性差异导致可能记录不同海水环境的温度信息[91]。因此,同一时期(层位)、不同属种牙形刺的氧同位素组成是否存在属种间差异,会影响古海水温度的重建结果。例如,Buggischet al.[92]通过对奥陶纪牙形刺研究发现,不同水深环境不同属种牙形刺的氧同位素组成不存在显著差别。此外,Joachimskiet al.[60]通过系统比较Frasnian-Famennian界线附近Polygnathus、Icriodus 和Palmatolepis三个属的牙形刺氧同位素组成,也未发现明显的差别;然而华南地区同一层段的研究发现,Palmatolepis属牙形刺的δ18O 值要比前两者分别偏低0.6‰~0.7‰和0.5‰~0.9‰[90]。事实上,诸多研究发现,同一时期(层位)牙形刺不同属种之间的δ18O 差值可达约0.5‰~1.1‰[93-94]。这种差异可能源于:1)不同属种牙形动物记录了不同水深环境的温度信号;2)牙形刺氧同位素或存在属种间的生命效应。另一方面,牙形刺氧同位素可能还存在“属种内差异”。据报道,同一地区奥陶系相同层位同一属种、不同个体的牙形刺瓷白质齿冠的δ18O平均值可相差约1.5‰(VSMOW),体现了部分牙形动物属种的生物习性复杂性[85]。此外,近年有研究发现,基于牙形刺氧同位素所推测的水深与其赋存围岩的沉积水深有时并不相符[58,85,90],二者关系仍需进一步探讨。因此,明确牙形动物的生物相或生物习性,尽量选取特定单一、浮游相属种牙形刺进行氧同位素分析,有助于更精准解译古海水温度变化趋势、更合理解读地质意义。未来,进一步厘定和完善各时代牙形动物的生态分布模型,有助于针对性地挑选适宜的牙形刺属种,优化重建古海水温度曲线。
4.2 成岩作用
牙形刺氧同位素古海水温度重建,必须考虑成岩作用对其组构和组分的改造影响。传统研究认为,牙形刺个体的玻璃质和瓷白质齿冠能较好的保存初始氧同位素信号[35,73]。然而,牙形刺虽然结构致密,但是在一定的成岩作用下,其初始氧同位素组成仍可能会发生改变[95]。因此,评估牙形刺样品的保存情况和成岩作用改造程度,是进行氧同位素分析测试和古温度精确重建的必要前提。
目前,色变指数(Color Alteration Index,CAI)被广泛用于评估牙形刺所经历的最高埋藏温度及成岩作用改造程度。该指数基于实验条件,按照牙形刺遭受热成岩作用改造强度由弱到强的顺序,依次将其对应颜色量化划分为1~8 等级[96-97]:成岩作用温度越高,则牙形刺CAI 值越大。通过对比蒙赛尔颜色图(Munsell color chart),可以定性判别牙形刺CAI值[98]。有研究认为,牙形刺CAI 值不超过5,其氧同位素组成通常保存较好[18,21,60]。该指标是评估牙形刺遭受成岩作用改造程度的常用指标,然而其依赖于主观视觉判断,且难以客观精确量化。另一方面,Shemesh[99]提出以磷灰石结晶度作为判断其遭受成岩作用改造程度的指标:结晶度越低,则改造程度越弱。随后,Pucéatet al.[100]基于拉曼光谱研究进一步建立了磷灰石结晶度定量化指标(crystallinity index,CI);然而,该指标与初始海水信号保存程度的对应关系较复杂,难以准确指示样品化学组分遭受成岩作用改造的程度。基于扫描电子显微镜(SEM)观察牙形刺磷灰石的形貌和表面结晶特征,可以直观定性评估其遭受重结晶和后期改造的程度[90,95]。此外,部分研究者尝试通过稀土配分模式(REE)和阴极发光(CL)特征来判断牙形刺的保存情况,但是同样难以有效评估其初始氧同位素信号的保存程度[101]。最新研究表明,牙形刺中有机质随着热成熟度不断增加而逐渐向石墨转变,据此可通过拉曼光谱定量评估牙形刺中有机质热演化程度,进而评估其遭受成岩作用改造的程度[102]。基于拉曼光谱评估方法针对加拿大密西西比阶—上三叠统牙形刺样品所估算的最高埋藏温度范围,与CAI指标的估算结果总体比较相近[103],支持了该方法的有效性。实践中,牙形刺瓷白质齿冠被认为在成岩改造过程中具有更好的结构和化学稳定性,常作为氧同位素信号提取的首选部位。然而也有研究报道,牙形刺瓷白质齿冠部位相对于玻璃质齿冠和基腔部位,却具有更轻的δ18O值,推测成岩作用对其改造程度却更显著[98]。因此,如何精确评估牙形刺氧同位素组成遭受成岩作用的改造程度,并优选氧同位素测试对象,有待进一步深入探讨。采用多种手段综合评估牙形刺不同部位的成岩作用改造程度,有助于有针对性地提取初始氧同位素信号,提升古温度重建可靠性。
4.3 量化计算
牙形刺氧同位素古海水温度重建,必须考虑量化计算所带来的不确定性。磷酸盐氧同位素—温度经验公式的适用性、磷酸盐标样标定值的准确性、同时期海水氧同位素赋值的合理性等因素,均可能影响海水温度的定量重建结果,以及相关数据的对比分析及其地质意义解读。
目前,针对相同的牙形刺氧同位素值,基于不同的氧同位素—温度经验公式,重建的温度结果可呈现明显的偏差(表2)。事实上,相关氧同位素—温度经验公式各自基于不同种类的生物组织或器官(如藤壶、舌形贝和鲨鱼牙齿等)、不同的化学处理流程和磷酸盐标样而建立,它们是否、以及在多大程度上适用于已经绝灭的牙形动物,需要进一步探讨或优化。同时,由于不同生物生存的温度适宜区间有所不同,相关经验公式理论上也受限于一定的温度适用范围。目前,磷酸盐氧同位素—温度经验公式仍在不断优化改进[32-33,37,104],至今尚未达成共识。近年,Lécuyeret al.[37]通过将舌形贝和鲨鱼牙齿磷酸盐和同时期碳酸盐的氧同位素温度重建结果进行对比和校正,建立了新的磷酸盐氧同位素—温度经验公式(表2),得到广泛认可和应用[105-106]。此外,由于测试条件和流程差异,不同实验室对于国际磷灰石标样佛罗里达磷块岩氧同位素组成δ18ONBS120c的测试结果存在可达0.9‰(VSMOW)的偏差[34-35,59]。这一定程度上也会影响相关测试结果,进而影响古海水温度量化计算结果及实验室间数据的对比讨论。对此,Pucéatet al.[33]将磷酸盐标样NBS120c 的δ18O 测量值作为浮动变量纳入氧同位素—温度经验公式(表2),以弱化不同实验室测试偏差对温度重建结果的影响。
另一方面,海水δ18Ow值是基于磷酸盐氧同位素—温度经验公式(表2)定量重建古海水温度的关键参数之一[15],但是其受控于多种影响因素而变化。目前普遍认为,由于冰川相对海水富集16O,全球海水平均δ18Ow值主要受控于全球冰量体积的变化[2]:在冰期时偏高,间冰期时则偏低[15],呈现较好的冰期和间冰期旋回波动。同时,显生宙海水δ18Ow值被认为受到海底热液活动的调控,长期稳定在0±1‰(VSMOW)[107-108],并呈现一定的长期演化[109]。Veizer 和Prokophet al.[4]基于对58 532 个低镁方解石质生物壳体δ18O 值的统计分析认为,显生宙海水氧同位素组成以约+0.01‰∕Ma(VSMOW)的速率不断变化。另一方面,局地海水δ18Ow值如果受区域气候条件、水体循环特征等因素的影响,可以偏离全球海水平均值,呈现一定的空间变化[110]。例如,基于现代全球表层海水(水深<5 m)年平均氧同位素组成的观测结果表明,由于存在蒸发量∕降水量比值差异,赤道地区海水δ18Ow值相对极地地区偏重超过5‰,存在明显的纬度差异;同时,局限水体的δ18Ow值相对开阔大洋可明显偏高[111-112]。因此,基于经验公式定量重建深时海水温度绝对值,其实是局部环境与全球气候背景的叠加信号[4,15],需综合考虑各种因素才能给予合理的地质解释(详见本文第5部分)。实践中,通常将δ18Ow值简化为全球海水平均值并进行特定赋值,对于地球两极无冰时期(如早奥陶世)常假定为-1‰(VSMOW)[35],对于地球两极有冰时期(如二叠纪乌拉尔世)常假定为+1‰(VSMOW)[58]。
5 牙形刺氧同位素古温度计的应用
5.1 应用现状
牙形刺氧同位素古温度计的应用虽然受限于多种因素的影响,但是对于重建深时海水温度仍具有不可替代的优势,可为我们理解古海洋、古气候、古地理和古生物演化提供宝贵信息,并已取得了大量研究成果。
牙形刺氧同位素古温度重建,大致勾画出了晚寒武世至三叠纪海水温度演化曲线,重点揭示了关键地质历史时期海水温度特征,促进了对深时环境和生命演化的认识(图3)。例如,奥陶纪劳伦和冈瓦纳大陆牙形刺氧同位素证据显示:早奥陶世初期海水温度超过40 ℃,然后逐渐降低,至中奥陶世时与现今赤道海水的温度范围(28 ℃~32 ℃)相当,修正了奥陶纪总体处于超级温室状态的传统观点[35]。泥盆纪牙形刺氧同位素证据显示:中泥盆世温室气候传统观点难以成立,反之具有适宜的海水温度(23 ℃~25 ℃),并促进了生物礁的繁盛[60];晚泥盆世弗拉期—法门期(Frasnian-Famennian)转折时期全球气候变冷,低纬度海域降温5 ℃~7 ℃,生物面临严峻生存压力并走向了大绝灭[31,90]。晚石炭世北美中大陆地区牙形刺氧同位素研究揭示,同一准层序内牙形刺δ18O差值可达1.7‰(VSMOW),可能受控于冰川成因的全球海平面变化,一定程度上可对比更新世冰期-间冰期情形[59]。早二叠世华南地区牙形刺氧同位素证据显示,晚古生代冰期的结束发生在空谷期,较传统观点(晚萨克马尔期)推迟了约10 Ma[58]。二叠纪—三叠纪转折时期华南和伊朗地区高分辨率牙形刺氧同位素证据显示,全球气候变暖滞后于该时期的生物大绝灭事件[40,115],更可能是早三叠世生物迟缓复苏的原因[24]。目前,牙形刺氧同位素古温度重建的相关研究主要集中于关键地质历史时期,而其他时期则相对薄弱甚至缺失,有待跟进;部分地质时期(如晚奥陶世和晚石炭世)牙形刺氧同位素所重建的古海水温度分歧较大,有时存在极值(如早奥陶世和早三叠世)(图3),其可靠性仍需进一步验证等。总体而言,牙形刺氧同位素古海水温度重建结果与前人基于其他指标重建的大气二氧化碳浓度曲线总体上具有相似的变化趋势[114](图3),进一步证实了该古温度计的有效性和合理性。
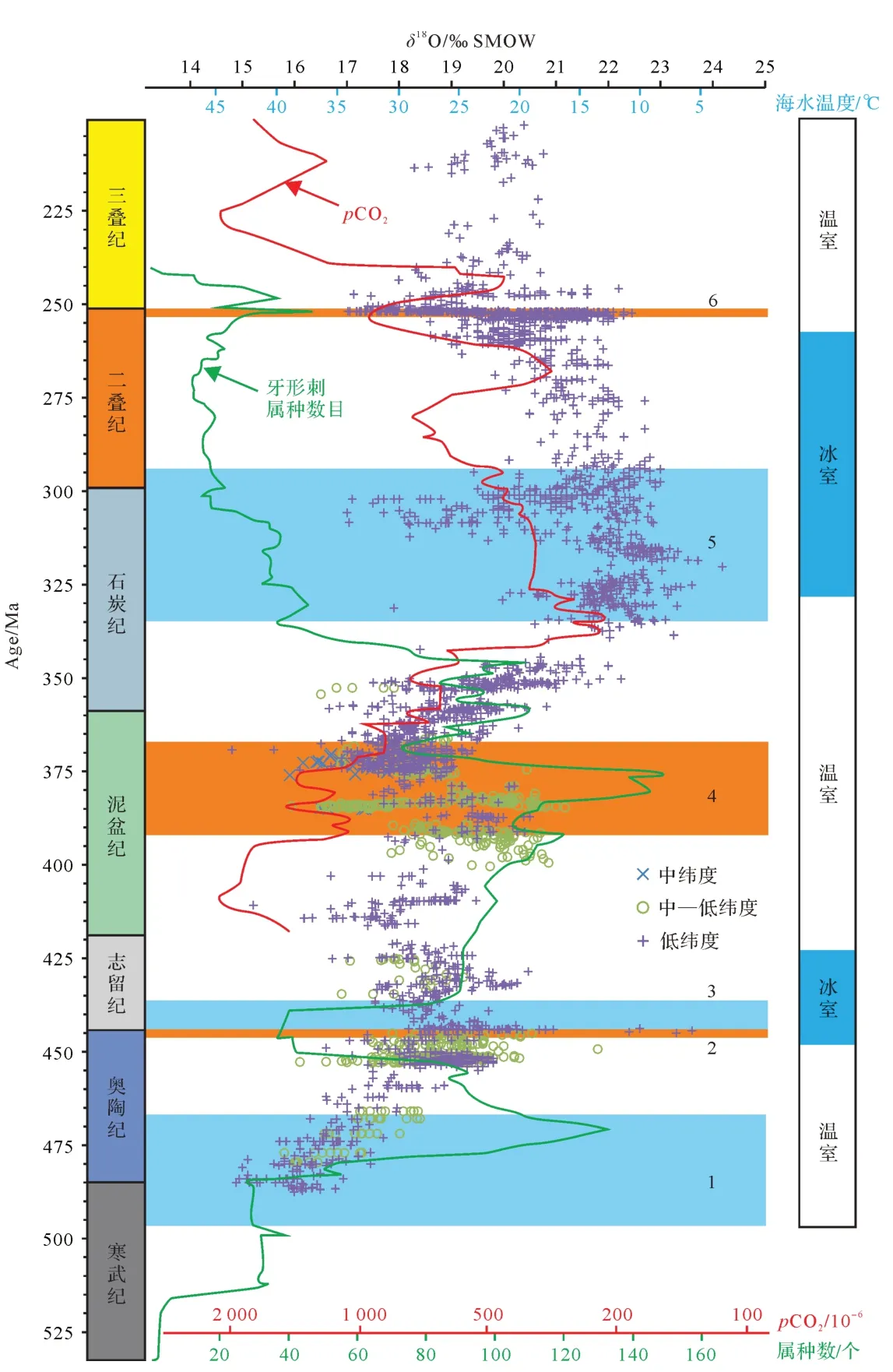
图3 寒武纪至三叠纪牙形刺氧同位素及其定量重建的海水温度演化序列和环境—生物演化背景牙形刺氧同位素数据整理自Huanget al.[90];Songet al.[105];Albanesiet al.[113]。CO2浓度重建结果和温∕冰室气候划分据Fosteret al.[114]修改。古温度的计算据Kolodnyet al.[32]。假设温室气候下海水δ18O平均值为-1‰(VSMOW),冰室气候时则为+1‰(VSMOW)。牙形刺属种数目变化曲线和重大生物演化事件据Fanet al.[49],其中1.奥陶纪生物大幅射;2.晚奥陶世生物大绝灭;3.早志留世生物辐射;4.中—晚泥盆世生物多样性骤减;5.晚石炭世—早二叠世生物辐射;6.二叠纪末期生物大绝灭。古纬度划分据Veizeret al.[4],分为中纬度(58°~35°)、中—低纬度(35°~26°)和低纬度(26°~0°)Fig.3 The distribution of conodont oxygen isotopes and calculated paleotemperatures from the Cambrian to the Triassic and associated major environmental and biological eventsOxygen isotope data of conodont apatite compiled after Huanget al.[90]; Songet al.[105]; and Albanesiet al.[113]. CO2concentration reconstruction and green∕icehouse climate identification after Fosteret al.[114].Paleotemperature calculation is based on Kolodnyet al.[32],assuming the seawater averageδ18O value of -1‰ (VSMOW) and +1‰ (VSMOW) for greenhouse and icehouse climates, respectively. Conodont species diversity curve and major bio-events are according to Fanet al.[49]:1.Great Ordovician Biodiversification Event;2.End-Ordovician mass extinction;3.Early Silurian radiation;4.Middle to Late Devonian diversity decline;5.Late Carboniferous-Early Permian biodiversification;6.End-Permian mass extinction.Paleolatitude is classified as middle latitude(58°-35°),mid-low latitude(35°-26°),and low latitude(26°-0°)according to Veizeret al.[4]
另一方面,牙形刺氧同位素古温度重建,促进了对牙形动物生物习性的认识。例如,基于早奥陶世弗洛期劳伦大陆斜坡—盆地相牙形刺的氧同位素研究发现,不同属种牙形刺的δ18O 值存在1.6‰~1.8‰(VSMOW)的差异,被认为源于在海水温跃层背景下牙形动物的生物习性差异,并证明牙形动物以游泳分层而非底栖游移的方式生活[85]。基于泥盆纪弗拉期—法门期(Frasnian-Famennian)转折时期华南地区常见牙形动物的氧同位素研究发现,Polygnathus和Icriodus 属牙形动物生活的水深范围较大,而Palmatolepis属牙形动物生活在表层水体中,是重建表层海水温度的优选对象[90]。基于石炭纪Donets 盆地牙形刺氧同位素的研究发现,Gondolella属和Idioprioniodus属牙形刺的氧同位素值相对于同时期生活在表层水体的属种,在宾夕法尼亚亚纪早期较一致,在宾夕法尼亚亚纪晚期则偏高0.5‰~0.8‰(VSMOW),推测它们随着海侵过程中水体循环样式和初级生产力的变化而主动调整了生活水深[23]。基于二叠纪不同板块牙形刺的氧同位素研究发现,Streptognathodus属和Hindeodus属牙形刺主要生活于近表层海水环境,然而Jinogondolella属和Clarkina属牙形刺则可能在海水表层和深水环境中都能生存[58]。基于晚三叠世意大利Lagonegro盆地牙形刺的研究发现,Epigondolella triangularis种牙形刺氧同位素可合理对应同时期热带—亚热带海表温度范围,表明其属于海洋表层透光带内游泳生活方式[84]。因此,牙形刺氧同位素相关研究,通过重建特定属种牙形动物生活的海水温度,有效揭示了其生物习性与环境响应,具有重要应用价值。
此外,牙形刺氧同位素古温度重建,为古地理格局提供了关键约束。例如,基于奥陶纪阿根廷Precordillera地块高分辨率牙形刺氧同位素古温度重建结果,对比发现该地块与劳伦大陆所记录的古海水温度变化样式在早—中奥陶世基本一致,至晚奥陶世早期则存在约6 ℃的偏差,表明二者古地理格局发生了一定的演化[113]。通过三叠纪卡尼期-诺利期(Carnian-Norian)转折时期牙形刺氧同位素重建结果,对比发现Wrangellia地块和Pangea大陆该时期海水温度偏差约3 ℃~5 ℃,并结合古地磁数据可将前者古地理位置约束在约10°~15°N[116]。因此,牙形刺氧同位素相关研究,通过重建和对比不同地块的古海水温度特征及变化样式,可揭示它们之间的古地理演化关系,并有助于结合古地磁证据共同精准约束古地理位置。
5.2 应用须知
牙形刺氧同位素古温度计由于具有定量重建古海水温度的优势,具有重要发展潜力;但是其地质意义的合理解读,受制于牙形动物自身生物习性、成岩作用改造程度、量化计算方式、古地理背景等因素,在应用时需加以关注和考量。例如,早三叠世同一层位不同属种牙形刺δ18O值差异,响应了牙形动物属种差异[24];二叠纪—三叠纪转折时期扬子板块台地相至斜坡相不同剖面牙形刺δ18O值差异,源于成岩作用不同程度的改造[40];石炭纪乌克兰半局限盆地相相对于中国华南地区斜坡相(纳庆剖面)牙形刺δ18O 值存在的显著差异(1‰~6‰),源于区域性季风环流的影响[23,117];二叠纪不同板块牙形刺δ18O 值差异,可能响应了各自蒸发量∕降水量比值的不同[58]。
同时,牙形刺氧同位素古海水温度重建结果的对比与解析,需考虑古纬度、古地理和古海洋因素(图3)。例如,早—中奥陶世牙形刺氧同位素重建古温度显示,中—低纬度地区的海水温度低于低纬度地区[35,113,118],具有显著的温度—纬度分布特征。此外,Albertiet al.[119]在对中侏罗世腕足动物和双壳类壳体氧同位素研究的过程中,将海水δ18Ow值分别进行-1‰(VSMOW)的传统赋值和参考温度—纬度梯度因素的改进赋值,发现基于后者所重建的古海水温度更加真实合理。另一方面,上升洋流发育的海域,如果深部不同温度和氧同位素特征的水体运移至陆架地区表层水体,可导致牙形刺氧同位素古温度重建结果具有明显的地域特色[120]。此外,局部水团的物理化学性质(如营养水平、浑浊度及氧气含量)也会影响牙形动物的生态分布[121],进而为牙形刺氧同位素古温度数据的地质意义解析带来一定的复杂性。
综上所述,基于有限研究区、特定属种牙形刺的氧同位素数据解读局部海水温度信号时需要十分谨慎。在确保牙形刺保存情况较好的前提下,不仅要考虑牙形动物自身生物习性对牙形刺氧同位素变化趋势的影响,也需要考虑研究区的古地理特征,包括局部气候条件(如盆地水体的局限性、蒸发量∕降水量比值、季风气候等)、古纬度位置、洋流特征等因素对局部水体温度和氧同位素组成的影响,以正确对比和合理解读局部温度信号、及其与全球气候的关系。
6 结语
牙形刺能较好的记录和保存原始海水氧同位素信号,同时,多年的机理探讨和分析测试方法开发使其氧同位素逐渐成为重建深时海水温度的关键温度计。目前,对牙形刺氧同位素古海水温度精准重建和精确解读,需综合考量生物习性、成岩作用、量化计算、古地理背景等多种因素的影响。该古温度计的广泛应用,极大地促进了我们对深时环境、海洋和生命演化的认识。未来,随着牙形刺氧同位素与海水温度关系的深入研究,分析测试方法的不断改进,其他古温度指标(如团簇同位素)的综合应用,以及对地质背景的全面理解,牙形刺氧同位素古温度计将在深时海水温度重建中发挥更大的作用。
致谢 非常感谢审稿专家对文章提出的细致而中肯的宝贵修改意见,对本文水平提升有很大帮助。
