巨桉幼树对镉胁迫的光合生理响应
2022-04-01胡红玲刘喜建代大川张如义
文 静,胡红玲,陈 洪,刘喜建,代大川,张如义
(1 四川农业大学 林学院,四川 成都 611130;2 西昌学院 农业科学学院,四川 西昌 615000;3 内江市农业科学院,四川 内江 641000)
由于工农业的快速发展,土壤重金属污染速度加快,同时土壤中重金属的有效性进一步提高[1]。对植物而言,镉(Cd)虽然不是其生长所需的营养元素,但Cd的移动性较强,易被植物根系吸收,土壤中镉过量会严重影响植物的生长发育[2],导致植物及其产品品质下降[3],并通过食物链直接威胁人类的健康[4]。镉进入植物体后通常会改变其细胞膜透性,使植物的光合作用、呼吸作用及氮同化能力等受到抑制,引起植物体内的生理生化过程紊乱,植物生长发育受阻,严重时会导致植株死亡[5]。四川省矿产丰富,重金属污染形势也较为严峻,如李志涛等[6]研究发现,江安县某硫铁矿区周边农田土壤镉超标率达99%。
巨桉(Eucalyptusgrandis)为桃金娘科(Myrtaceae)桉属(Eucalyptus)的一个种,具有适应性强、速生、高产等特点,是联合国粮食及农业组织推荐的三大速生造林树种之一[7],为当今世界栽培面积最大的一种桉树[8]。巨桉在我国南方地区已成为营造短周期工业原料林的重要树种,也是四川栽培的主要树种之一,广泛分布在川中、川南、川东南、川西南边缘地带。因其特殊的材质、纹理特征和药用成分,被广泛应用于造纸、人造板、农药和医药等行业,对解决我国木材供需矛盾、农林业产业结构调整以及地方经济的提升具有重要作用[9-11]。目前对于巨桉的研究多集中在栽培经营技术、耗水特性、干旱胁迫响应等方面[12-14],而有关巨桉对重金属胁迫耐受机制的相关研究并不多见,特别是从光合生理特性方面进行的探索更少。植物的光合作用对重金属较为敏感,而叶绿素荧光是研究植物光合性能的理想探针,其参数的变化不仅可以直观反映胁迫对植株的影响,还可用于评价植株的抗逆性能[15]。
本研究以巨桉幼树为材料,通过盆栽试验探讨了其光合速率、光响应、叶绿素荧光等对不同水平镉胁迫的响应,以期为巨桉适宜立地条件的选择以及镉污染生境修复的可行性提供参考依据。
1 材料与方法
1.1 试验地概况
试验地位于四川农业大学(成都校区)教学科研试验基地,地处东经103°51′29″,北纬30°42′18″,属亚热带湿润季风气候区,四季分明,气候温和湿润,夏季高温多雨。年均总日照时数840.2 h,年均降水量759.1~1 155.0 mm;年均气温16.8 ℃,月平均最高温为25.9 ℃(7月),月平均最低温为6.9 ℃(12月),极端高温38 ℃(7月),极端低温-2 ℃(1月);年均相对湿度84%。
1.2 试验材料
1.2.1 栽植土壤与容器 土壤取自雅安坡积土,使用前除去土壤中过大的植物残体和石砾。土壤理化性质为:pH 6.5,全氮15.06 g/kg,全磷1.69 g/kg,全钾12.10 g/kg,镉0.72 mg/kg。栽植容器采用规格一致的塑料盆(盆口直径30 cm、高26 cm、底径25 cm),每盆等量装土8 kg(干土质量)。
1.2.2 供试植株及其栽培管理 2017年12月,选取长势良好、健康无病虫害的巨桉幼苗(组培苗,苗龄近4个月,平均高15 cm),每盆栽植1株,移栽后常规水肥管理,进行5个月的适应性培养。在此期间采用HH2土壤水分测定仪(ML2x,GBR)定期监测土壤水分状况,使土壤体积含水量保持在18%左右(即该试验用土田间持水量的70%~80%)。试验开始前15 d,将所有待处理植株移入通风透气的大棚内并停止施肥,以避免肥料对镉效应的干扰。施镉时苗龄约9个月(2018年5月),平均株高(86.3±4.2) cm,平均地径(7.98±0.96) mm,植株健康整齐。
1.3 试验设计
本试验采用单因素完全随机试验设计。参照国家环境质量标准(GB 15618-2018)[16]以及四川省农村土壤铅、镉含量调查[17]结果,试验共设置5个镉处理水平,分别为CK(0 mg/kg)、T1(12.5 mg/kg)、T2(25 mg/kg)、T3(50 mg/kg)和T4(100 mg/kg),每个处理重复3次,添加镉的形态为CdCl2·2.5H2O,对应于5个处理的添加量依次为0.00,25.39,50.78,101.56和203.11 mg/kg,均以溶液的形式添加到土壤中。为方便分析阐述,本研究将处理T1、T2称为低水平, T3、T4称为高水平。
试验于2018年5月8日开始,为符合镉在自然条件下逐渐排放的实际,将各处理镉施入量等分为5份,于2018年5-9月分5次施入。每盆用500 mL去离子水与CdCl2·2.5H2O配制成溶液,以植株根颈处为中心逐步向外围均匀施入,避免溶液溅到植物表面。CK浇等量的去离子水,试验期间的水分管理亦使用去离子水。2018年10月底,每处理选取3~4个植株上相同叶位(树冠上2/3处)的成熟功能叶,采用Li-6800便携式光合测定仪(Li-Cor Inc., USA)分别测定气体交换参数、光合-光响应特征曲线以及叶绿素荧光参数;随后采集功能叶,装入密封袋(不封口),标记好后置于冰盒中,即刻带回实验室测定叶绿素含量。
1.4 测定指标及方法
1.4.1 光合色素含量 叶绿素含量参照熊庆娥[18]的方法,采用直接浸提法测定。采集叶片后,将其擦净、剪碎,置于V(丙酮)∶V(无水乙醇)∶V(水)=4∶5∶1的提取液中提取光合色素,再用分光光度计分别在波长665,649和470 nm下测定吸光度,然后计算叶绿素a、b和类胡萝卜素的含量。
1.4.2 光合特性 净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)使用Li-6800便携式光合测定仪测定。参照李辛等[19]的测定方法并进行适当调整。于2018年10月选取晴天上午9:00-11:30时段,选取完全展开的成熟功能叶,每个处理重复测定3株,每株测定3个叶片。测定时设定参比室CO2含量为400 μmol/mol,叶室温度为30 ℃,叶室相对湿度为50%,光照强度设定为环境光强(1 000 μmol/(m2·s))。
1.4.3 光合-光响应曲线 通过预试验测得巨桉幼树的光饱和点在800 μmol/(m2·s)左右,故首先将巨桉幼树叶片在800 μmol/(m2·s)的光强下进行光诱导直至Pn稳定(约需30 min),然后使用Li-6800便携式光合测定仪的光响应曲线自动测定程序进行测定,测定时间和各参数设置同上,光强梯度设定为1 800,1 500,1 200,1 000,800,600,400,200,150,100,75,50,25和0 μmol/(m2·s)。
1.4.4 叶绿素荧光参数 选择与光合特性测定相同的巨桉叶片,使用Li-6800便携式光合测定仪进行测定,测定时间和各参数设置同上。参照Ali等[20]的方法并进行适当调整。测定前首先对叶片进行暗适应(用锡箔纸包裹待测叶片),待暗适应一整夜后,第2天测定初始荧光(F0)和最大荧光(Fm),然后根据各遮阴处理的环境光强设定活化光,光活化50 min以上后,再测定任意时间实际荧光产量(F)、光下最小荧光F0′和光下最大荧光Fm′,每处理测定3片叶子。根据测定数据,计算可变荧光Fv(Fm-F0)、PSⅡ潜在活性(Fv/F0)、PSⅡ最大光化学量子产量(Fv/Fm)、PSⅡ实际光化学量子产量ΦPSⅡ((Fm′-F)/Fm′)、电子传递速率ETR(ΦPSⅡ×0.5×PPFD)(其中PPFD表征吸收光的光量子通量,单位为μmol/(m2·s))、光化学淬灭系数qP((Fm′-F)/(Fm′-F0′) )和非光化学淬灭系数NPQ(Fm/Fm′-1)[21]。参照Demmig-Adams等[22]提出的方法计算PSⅡ吸收光能的分配比例,光化学反应耗散的能量可用公式P=qP×Fv′/Fm(其中Fv′指光下可变荧光产量,Fv′=Fm′-F0′)求得,根据D=1-Fv′/Fm′可求出用于天线热耗散的能量;通过Ex=(1-qP)×Fv′/Fm′可求得反应中心非光化学反应耗散的能量。
1.5 数据处理与分析
使用Microsoft Excel 2010软件对试验数据进行整理,采用SPSS 22.0(SPSS Inc., USA)统计分析软件进行单因素方差分析(One-way ANOVA),显著水平α设定为0.05,用Duncan’s新复极差法进行多重比较,用Origin 2018软件(Origin Lab.,USA)绘图。
2 结果与分析
2.1 镉胁迫对巨桉幼树叶片光合色素含量的影响
由表1可以看出,随着镉处理水平的提高,巨桉幼树叶片叶绿素a(Chl a)、叶绿素b(Chl b)、总叶绿素(Chl(a+b))和类胡萝卜素(Car)含量均呈下降趋势,但Chl a、Chl b和Chl(a+b)含量仅以T4处理较CK下降显著(P<0.05),降幅分别达34.2%,31.0%和33.2%;而Car含量从T2处理开始显著下降(P<0.05)。此外,各处理叶绿素a与b的比值(Chl a/b)与对照间均无显著差异。上述结果表明,巨桉幼树叶片类胡萝卜素含量对镉处理的反应较叶绿素更加敏感,而叶绿素a对镉处理的敏感程度与叶绿素b相似。
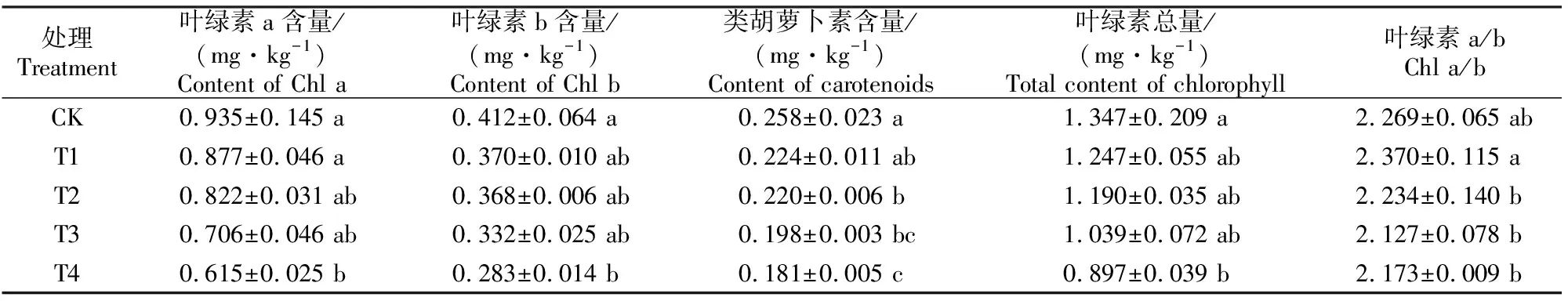
表1 不同水平镉处理下巨桉幼树叶片的光合色素含量Table 1 Changes of photosynthetic pigment contents in E. grandis saplings under different cadmium treatments
2.2 镉胁迫对巨桉幼树光合特性的影响
由表2可知,随着镉处理水平的提高,巨桉幼树叶片的Pn和Gs均呈下降趋势,到高水平处理(T3、T4)时与CK差异均达显著水平(P<0.05);Tr表现出相同的变化趋势,但从T2处理开始较CK显著降低;而Ci随镉处理水平的提高呈上升趋势,且从T2处理开始显著高于CK(P<0.05)。这说明巨桉幼树叶片光合作用能力下降的主要原因是非气孔限制因素所致。

表2 镉胁迫对巨桉幼树光合特性的影响Table 2 Effects of cadmium stress on photosynthetic characteristics of E. grandis saplings
2.3 镉胁迫对巨桉幼树叶片光合-光响应特征的影响
由图1可以看出,不同水平镉处理巨桉幼树光合-光响应曲线的变化趋势基本相似,即在低PAR(≤150 μmol/(m2·s))条件下,巨桉Pn随PAR的增强而迅速增加;之后随着PAR的持续增强,各处理植株Pn增长变缓,逐渐接近最大净光合速率Pmax;而当PAR≥1 000 μmol/(m2·s)时,Pn则缓慢下降。在同一PAR下,随着镉水平的提高,Pn呈下降趋势,且始终表现为CK>T1>T2>T3>T4。
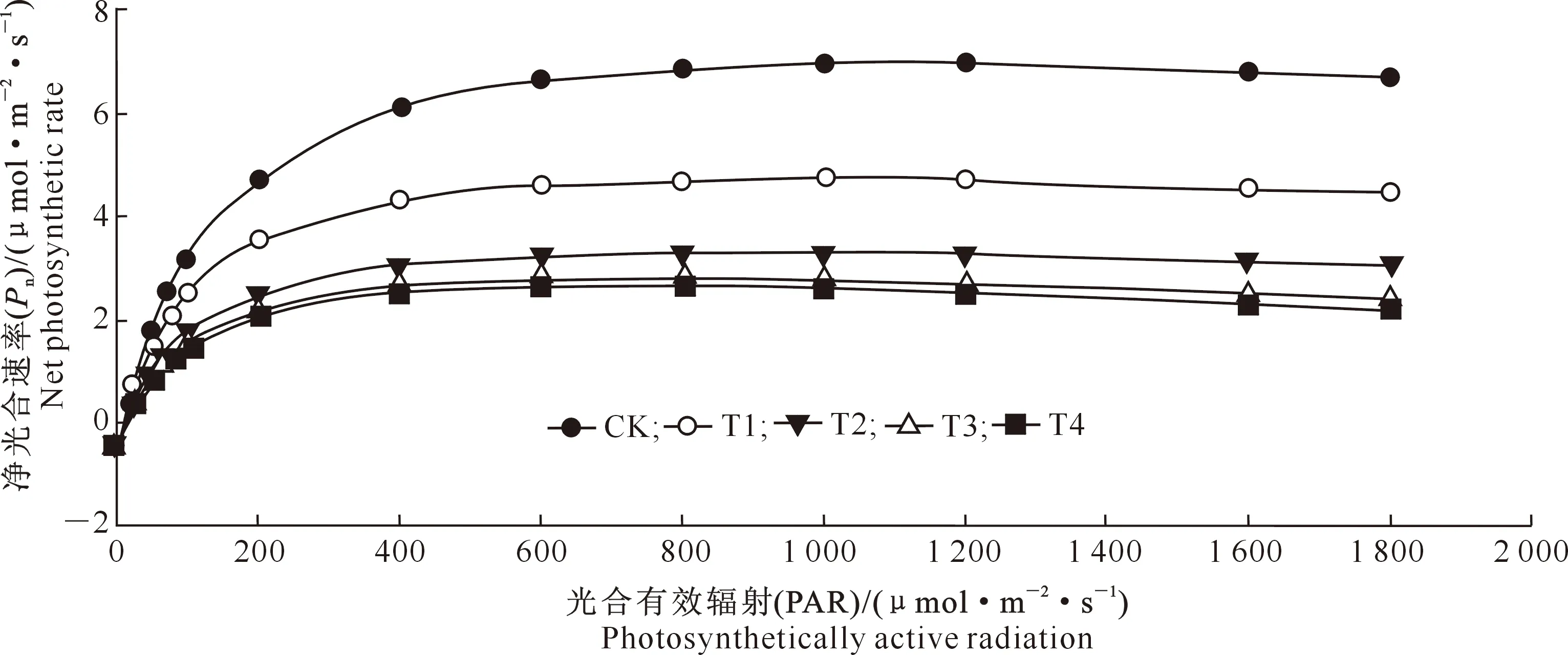
图1 镉胁迫对巨桉幼树光合-光响应曲线的影响Fig.1 Effect of cadmium stress on photosynthetic-light response of E. grandis sapling
由表3可见,随着镉处理水平的提高,巨桉幼树叶片的Pmax呈下降趋势,各处理均显著低于CK(P<0.05),T3与T4处理之间差异不显著(P>0.05)。AQY、LSP和Rd也随着镉胁迫加重而逐渐降低,并从T2开始显著低于CK(P<0.05)。LCP则随着镉胁迫加重逐渐升高,并从T3处理开始显著高于CK(P<0.05)。该结果表明,在镉胁迫下,巨桉幼树光合机构遭受到一定程度的破坏,使得有机物合成、光转化效率、弱光利用能力均有不同程度的下降。

表3 镉胁迫对巨桉幼树光响应曲线特征参数的影响Table 3 Effects of cadmium stress on characteristic parameters of photosynthetic light response of E. grandis saplings
2.4 镉胁迫对巨桉幼树叶绿素荧光参数的影响
由表4可知,巨桉幼树叶片F0、Fm和Fv/Fm均随着镉处理水平提高而逐渐降低,Fv/F0随着镉处理水平提高先升高后降低,但均仅在T4处理下与CK达到差异显著水平(P<0.05),分别较CK下降23.7%,33.0%,10.2%和6.7%;ΦPSⅡ在T2~T4处理下显著低于CK(P<0.05),ETR则表现为所有镉处理均显著低于CK(P<0.05),而qP和NPQ在各处理间无显著变化(P>0.05)。这表明,过量的镉导致巨桉幼树叶片PSⅡ受损,并出现明显的光抑制现象。

表4 镉胁迫对巨桉幼树荧光参数的影响Table 4 Effects of cadmium stress on fluorescence parameters of E. grandis saplings
2.5 镉胁迫对巨桉幼树吸收光能分配的影响
巨桉幼树叶片对吸收光能分配的耗散情况如表5所示。由表5可知,对照(CK)巨桉幼树将吸收光能的66.2%用于天线热耗散(D),6.1%用于光化学反应耗散(P),29.8%用于非光化学反应耗散(Ex)。随着镉处理水平的提高,用于天线热耗散的光能比例逐渐减小,在T4处理时与CK差异达到显著水平(P<0.05),降幅为3.9%;用于光化学反应耗散的比例也逐渐下降,且其对镉胁迫的反应更敏感,从T2处理时就有显著下降(P<0.05);而用于非光化学反应耗散的比例逐渐增加,在T4处理时与CK差异达到显著水平(P<0.05),增幅为12.1%。该结果表明,随着镉胁迫的加重,巨桉叶片光合保护和清除功能逐渐减弱,过剩光能增加。
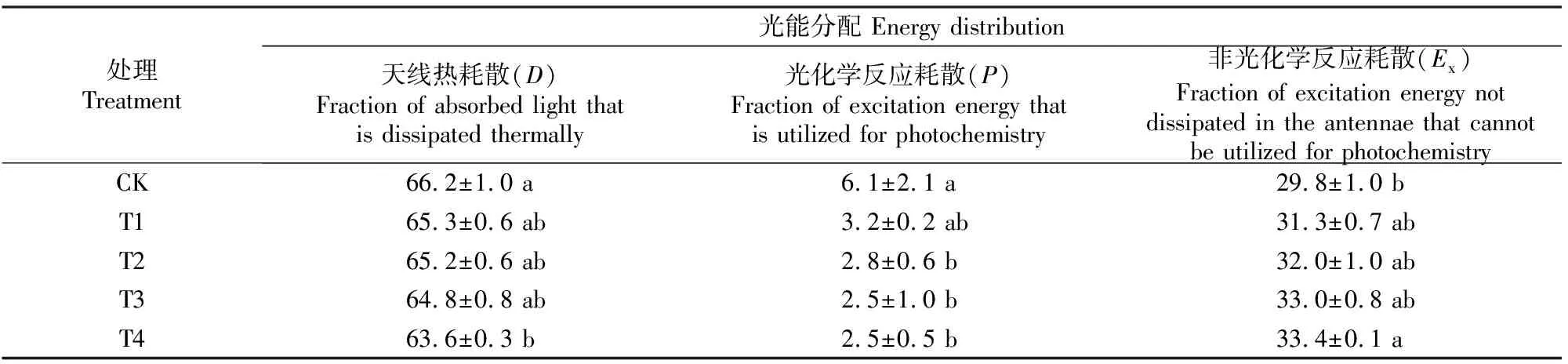
表5 镉胁迫对巨桉幼树叶片吸收光能分配的影响Table 5 Effect of cadmium stress on distribution pattern of absorbed light in leaves of E. grandis saplings %
3 讨 论
3.1 镉胁迫对巨桉幼树光合色素含量的影响
叶绿素含量是衡量植物光合作用的重要指标,在光合作用中发挥着吸收、传递及转化光能的作用,可在一定程度上反映植物的光合作用水平[23]。本研究中,巨桉叶片Chl a、Chl b含量随镉水平升高呈下降趋势,这与赖秋羽等[24]的研究结果相似。镉胁迫造成巨桉幼树叶绿素含量减少,可能是因为镉与叶绿体蛋白上的巯基结合破坏了叶绿体结构[25],也有可能是镉与叶绿素的某些酶,如胆色素原脱氨酶结合,造成酶失活,导致叶绿素合成受阻[26],还有可能是镉胁迫造成内囊体数量减少所致[27]。Car是叶绿体光合天线的辅助色素,能吸收过剩光能,以保护叶绿素和光合机能[28]。本研究中,巨桉幼树叶片Car含量随镉水平升高显著降低,佐证了叶绿素和光合机能受损的推断,因而其光合效率降低。Chl a/b值反映了叶绿体中类囊体的垛叠程度[29],逆境胁迫下植物Chl a/b值变化越小,表明其对逆境的适应性越强[30]。本研究中,巨桉幼树叶片Chl a/b在镉处理水平≥25 mg/kg时较对照明显降低,可能是镉水平过高时诱发的活性氧引起了叶绿素的破坏,致使叶绿素含量下降,同时Chl a/b也有所降低,但与对照差异不显著,表明巨桉对镉有一定的耐受能力,这与张明生等[31]的研究结果相似。
3.2 镉胁迫对巨桉幼树光合特性和光合-光响应特征的影响
植物受到胁迫时,其Pn降低的主要原因有气孔限制因素和非气孔限制因素[32],而判断依据是Ci和Gs2个指标值的变化方向。本研究中,随着镉胁迫程度的加重,Pn逐渐降低,Ci增加,表明镉胁迫环境中,限制巨桉幼树光合速率的主要原因是非气孔限制因素,这与107杨(Populus×euramericanacv‘74/76’)在镉胁迫条件下的响应特征相似[33]。Ci增加的原因可能是在镉胁迫下,巨桉幼树叶片光合器官受损,色素合成受到抑制,进而影响碳固定,从而造成Ci积累。高会玲等[34]的研究也得到了类似的结果。
植物对光的利用效率是其生长发育的重要指标。光补偿点LCP和光饱和点LSP分别反映植物对弱光和强光的适应能力,表观量子效率AQY则体现植物对光的利用能力[35]。本研究显示,镉胁迫下巨桉幼树叶片的AQY和LSP降低,LCP升高,这与张仁和等[36]的研究结果相似,表明镉胁迫抑制了巨桉幼树对光能的转换效率,导致光合速率下降。LSP降低和LCP升高也说明,在高水平镉胁迫下巨桉幼树叶片固定CO2的能力有所降低。
从4个光合特性参数的变化趋势可以看出,在T1处理时,巨桉幼树的Pn、GS、Tr和Ci均与CK差异不显著。从光合-光响应特征参数的变化趋势也可以看出,除Pmax外,其他特征参数(AQY、LSP、LCP、Rd)在T1处理时均与CK差异不显著,说明低水平镉胁迫对巨桉幼树光合生理特性影响不显著,巨桉的生长也未受到明显抑制。而在重度镉胁迫(T3、T4)时,各参数均与CK差异显著,表明受胁迫巨桉的生长受到明显抑制,这与万晋麟等[32]对辣椒(CapsicumannuumL.)的研究结果相似。
3.3 镉胁迫对巨桉幼树叶绿素荧光参数的影响
由于常温下叶绿素荧光主要来源于光系统Ⅱ的叶绿素a,而光系统Ⅱ处于整个光合作用的最上游,因此包括光反应和暗反应在内的多数光合过程的变化,都会反馈给光系统Ⅱ,进而引起叶绿素a荧光的变化,几乎所有光合作用过程的变化都可以通过叶绿素荧光反映出来[37]。因此叶绿素荧光参数的变化可以反映出植物光合机构在重金属胁迫下所表现的适应性[38]。本研究中,在镉胁迫程度最重(T4)时,巨桉幼树叶片光系统Ⅱ(PSⅡ)最大光化学量子产量(Fv/Fm)、PSⅡ潜在活性(Fv/F0)值减小,说明镉胁迫导致巨桉幼树叶片PSⅡ潜在活性中心受损,从而使原初光能捕获效率降低,这可能是由于叶绿体超微结构的改变所致[39];PSⅡ实际光化学量子产量(ΦPSⅡ)的下降则表明植物的光合磷酸化受阻,也可能与镉引起的叶绿体膜系统紊乱有关[40];电子传递效率ETR反映了实际光强条件下的表观电子传递效率,其数值降低表明光合电子传递受到抑制,进而导致ΦPSⅡ的下降。光化学淬灭系数qP反映PSⅡ反应中心的开放程度,非光化学淬灭系数NPQ是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分,反映了光系统对过剩光能的耗散能力[41]。本研究表明,镉胁迫处理巨桉幼树叶片的qP和NPQ与对照均无显著差异,NPQ无显著变化可能与此时镉胁迫使巨桉幼树热耗散机制受到损坏、PSⅡ原初光化学反应也受到了镉的伤害有关,这与前人研究的镉胁迫对黄瓜(Cucumissativus)[42]、龙葵(Solanumnigrum)[43]和滇苦菜(Picrisdivaricata)[44]的影响结果类似。
3.4 镉胁迫对巨桉幼树吸收光能分配的影响
在逆境条件下,植株为减轻过剩光能对叶片光合机构造成的损伤,会主动降低对光能的吸收,通过多种光能耗散机制调节光能的转换和分配[45]。植物叶片吸收的光能分为3部分,即天线热耗散的能量(D)、光化学反应的能量(P)、反应中心由非光化学反应耗散的能量(Ex)[46]。本研究发现,镉胁迫会使巨桉幼树PSⅡ叶绿素荧光能量分配发生改变,导致Ex比例增加而引起P和D比例降低,造成植物光合机构的可逆失活甚至破坏。D的变化与天线色素变化有关[47],而天线色素主要由Chl a、Chl b和Car组成,Chl a、Chl b含量的减少导致D降低;Ex是吸收光能中通过光呼吸、Mehler反应等非光化学反应进行耗散或将电子传递给氧而使其形成单线态氧的部分,Ex的上升反映过剩光能对光合结构的破坏程度加重。
4 结 论
镉胁迫导致巨桉幼树光合色素含量及光合特性和光合-光响应特征中的大多数指标或参数明显下降,光合电子传递活性减弱,造成两个光系统之间激发能分配不平衡、光化学效率降低、CO2同化能力下降;同时在光能分配受到影响后,引起PSⅡ反应中心的光能转化效率降低,导致巨桉幼树叶片光合效率下降,因此应当尽量避免在镉污染严重地区发展巨桉人工林,也不宜选择巨桉作为镉富集植物用于镉污染区的生态修复。但在镉污染程度较轻的地区,如土壤中镉含量在12.5 mg/kg以下时,种植巨桉还是可行的。
