光强对4种鸭跖草科植物生长和光合特性的影响
2022-03-31刘筱玮
陈 斌,刘筱玮,贾 琳,杨 扬,何 淼,*
1 东北林业大学园林学院, 哈尔滨 150040
2 东北林业大学林学院, 哈尔滨 150040
光是自然界中一切植物进行各项生命活动的物质基础[1],主要通过光强、光质、光周期等因素影响植物的地理分布和生长发育,光强作为其中一个重要因素,对植物的生长发育、形态结构、生理代谢、信号转导以及光合生理等方面均产生重要影响[2—6]。当外界光强发生变化时,植株体的生长状态必然会发生一定程度的改变,这主要是由于植物对光的主动捕获能力以及光合产物的合成能力受到了影响造成的[7],研究发现,植物在弱光环境会通过增加叶面积、总生物量、比叶面积、叶面积比、叶绿素含量的策略,使叶片变大变薄,增大对光的截获面积,提高对有限光资源的利用效率,增加气孔导度,维持较高的光合速率,以此来确保植株体在弱光环境中进行正常的生命活动[8—9],而在强光环境中,植物则通过增加叶片厚度,缩小叶片面积,降低叶绿素含量的方式减少对过剩光源的捕获,又可通过关闭气孔的方式减少叶片水分的蒸腾,达到减轻强光对植物体伤害的目的[10—11]。
随着强调生态平衡又兼具注重人类健康和文化发展的“绿色城市”理念的提出以及党的十八大“美丽中国”战略的实施,城市生态环境的保护和改善受到了前所未有的关注,城市居民对生活环境提升改善的需求也日益增加,这在很大程度上促进了城市园林建设事业的发展。但当前城市建设缺乏科学合理的统筹规划,使得许多建筑物布局不合理,间距狭小,导致大量建筑物间的园林绿化用地光照不足。此外,日益发达的立体交通也导致了大量极端阴蔽空间的出现,有研究表明,当前城市中有一半以上的园林绿地常年接受不到充足的光照[12],这使得城市中发挥“绿肺”功能的园林植物生长受阻,无法发挥最佳的生态环境效益,同时这也给广大园林建设者带来了巨大的挑战。因此,选育观赏价值高且对光环境要求低的园林植物对阴蔽空间进行科学有效的绿化,对发挥城市绿地最佳的生态环境效益具有深远意义。鸭跖草科(Commelinaceae)植物有40属500余种,其中有许多种为彩叶植物,颜色丰富观赏效果极佳。该科植物不仅对生长环境要求不高,具有一定的耐阴能力,且繁殖简单,生长迅速,能在短时间内呈现良好的景观效果,目前在我国广东、福建等地已有部分种类作为观叶地被植物应用于滨水驳岸、林下绿地等多种形式的城市园林中,在东北地区也作为地被植物被用于观光温室中的林下或苗床下空间,起到美化裸露地表的作用。其中,紫鸭跖草(Commelinapurpurea)、花叶水竹草(Tradescantiaflurnuensis‘Variegata’)、吊竹梅(Tradescantiazebrina)、绿叶水竹草(Tradescantiaflurnuensis‘Vairidia’)是目前最常用于城市园林绿化中的4种鸭跖草科植物,虽然在实际应用中发现它们具有较好的耐阴性,但每种植物生长所需的最适光强以及适光策略目前尚不明晰,因此,本试验欲通过不同密度的遮光网模拟5种不同光强的立地环境,对紫鸭跖草、花叶水竹草、吊竹梅和绿叶水竹草进行光处理,测定不同光强下4种植物的形态特征、叶片解剖结构以及光合生理等指标,探究4种植物生长的适宜光强以及适光策略,为在园林中更加科学地栽培应用提供理论依据。
1 试验材料与方法
1.1 试验材料
试验于2019年3—7月在黑龙江省哈尔滨市东北林业大学花卉研究所苗圃进行。以紫鸭跖草(Commelinapurpurea, CP)、花叶水竹草(Tradescantiaflurnuensis‘Variegata’, TF)、吊竹梅(Tradescantiazebrina, TZ)、绿叶水竹草(Tradescantiaflurnuensis‘Vairidia’, TV)为试验材料,于3月初剪取当年生健壮枝条顶部约5 cm的茎段,用湿润的报纸包裹好后放置在阴凉处备用。将腐殖土和蛭石(体积比为3:1)充分混匀,120 ℃高温高压灭菌后装入直径12 cm,高18 cm的花盆中,将盆中的栽培基质充分浸湿,待多余水分控出后进行扦插。每盆扦插2个茎段,每种植物材料扦插60盆。将扦插苗放置在温度为24—26 ℃,相对湿度为60%—70%,光强为480 μmol m-2s-1,光照时长12 h的育苗室内缓苗。扦插苗生长1个月后,选取生长良好,无病虫害的植株进行后续的试验[13—14]。
1.2 试验方法
于2019年4月30用木方搭设遮光架,每个遮光架间隔50 cm,顶部距离植株体顶端大于30 cm,然后使用不同密度的黑色遮光网罩在遮光架上,设置5种光强环境(分别为自然光强的100%、75%、50%、25%、5%,记作L0、L1、L2、L3、L4),每个光强环境每种植物材料处理12盆,3个重复。每5 d浇一次水,处理45 d后,进行各项指标的测定[13—14]。
1.3 形态指标的测定
株高的测定:用直尺测量植株根茎部至茎尖的高度,将此高度定义为株高。
叶片夹角测定:用量角器测定植株第5片完全展开叶片的主脉与茎之间所形成的夹角,将此夹角定义为叶片夹角。
叶面积的测定:取植株茎顶部第3—5片成熟的叶片,每盆取10片,每个处理3个重复,用Li- 3000A叶面积仪测定叶面积。叶面积测定后,将叶片用报纸包好后放入60℃的烘箱中烘干至恒重,计算比叶面积(比叶面积=单叶面积/单叶干重)。
1.4 叶片显微结构的观察
叶片解剖结构的测定:摘取植株顶端第3片完好无病虫害的叶片,在叶片上切取0.3 cm×0.1 cm的组织块迅速放入用福尔马林-乙酸-乙醇(Formalin- acetic acid-alcohol,FAA)固定液中固定,每盆取3片,每组处理3个重复。采用常规石蜡法制片[15],用Leica DM2500显微镜拍照,并用显微镜自带的标尺测量叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度,并计算栅栏组织和海绵组织比。所得数据为9个视野的平均值。
叶片气孔开度的测定:于晴天10:00选取植株茎顶部第3—5片成熟完整的叶片用去离子水擦除叶面灰尘,并用吸水纸吸附叶表残余的水分,摘取预处理好的叶片,迅速将无色指甲油均匀涂抹在叶片正面远轴面中部,采用指甲油印迹法对植物材料进行取样[16—18],待指甲油自然晾干后,用尖嘴镊子从一侧轻轻撕下叶表透明的指甲油膜,放到载玻片上制作临时装片,用Leica DM2500显微镜观察并拍照,随机选取每个视野上3个完整清晰的气孔,测量气孔开度(保卫细胞之间形成的孔隙的最大值),所得数据为9个视野的平均值。
1.5 叶绿素含量的测定
叶绿素含量的测定:摘取茎顶部第3—5片成熟叶片,参照刘萍等[19]的丙酮和乙醇混合液浸提法提取并测定叶绿素a(Chlorophyll a,Chl a)、叶绿素b(Chlorophyll b,Chl b)和总叶绿素(Chl (a+b))的含量并计算叶绿素a/b (Chl a/b)的值;
1.6 光合生理指标的测定
选择晴朗无风的9:00—11:30进行光合生理指标的测定,选取植株顶部第4片完全展开且完整无病害的叶片为测定叶片,将 Li- 6400光合仪设置为开放气路,叶室空气流量为500 mL/min,然后利用仪器自带的红蓝光源叶室(PAR=800)测定净光合速率(Pn)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)和蒸腾速率(Tr),每个处理重复3次。
1.7 数据处理
用Excel 2016对数据进行整理和作图,利用SPSS 22.0对数据进行方差分析,用Duncan 法进行多重比较(P<0.05),并对结果进行双因素(ANOVAs)分析。
2 结果与分析
2.1 不同光强对形态指标的影响
不同光强对4种植物的株形产生了显著的影响(P<0.05)(图1和图2)。CP、TF、TZ、TV的株高随光强的降低而显著降低(P<0.05),与L0相比,在L4条件下分别升高了1.07倍、0.81倍、0.87倍、1.35倍。相同的是,叶片夹角也随光强的降低而显著增加(P<0.05),与L0相比,在L4条件下分别增加了0.81倍、0.98倍、1.43倍、1.08倍。
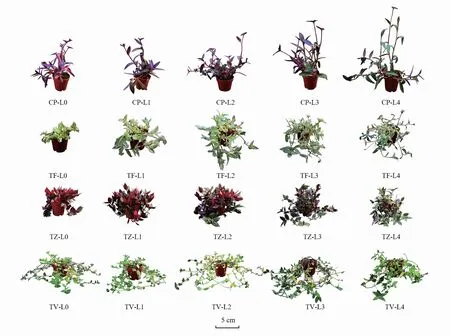
图1 不同光强对株形的影响
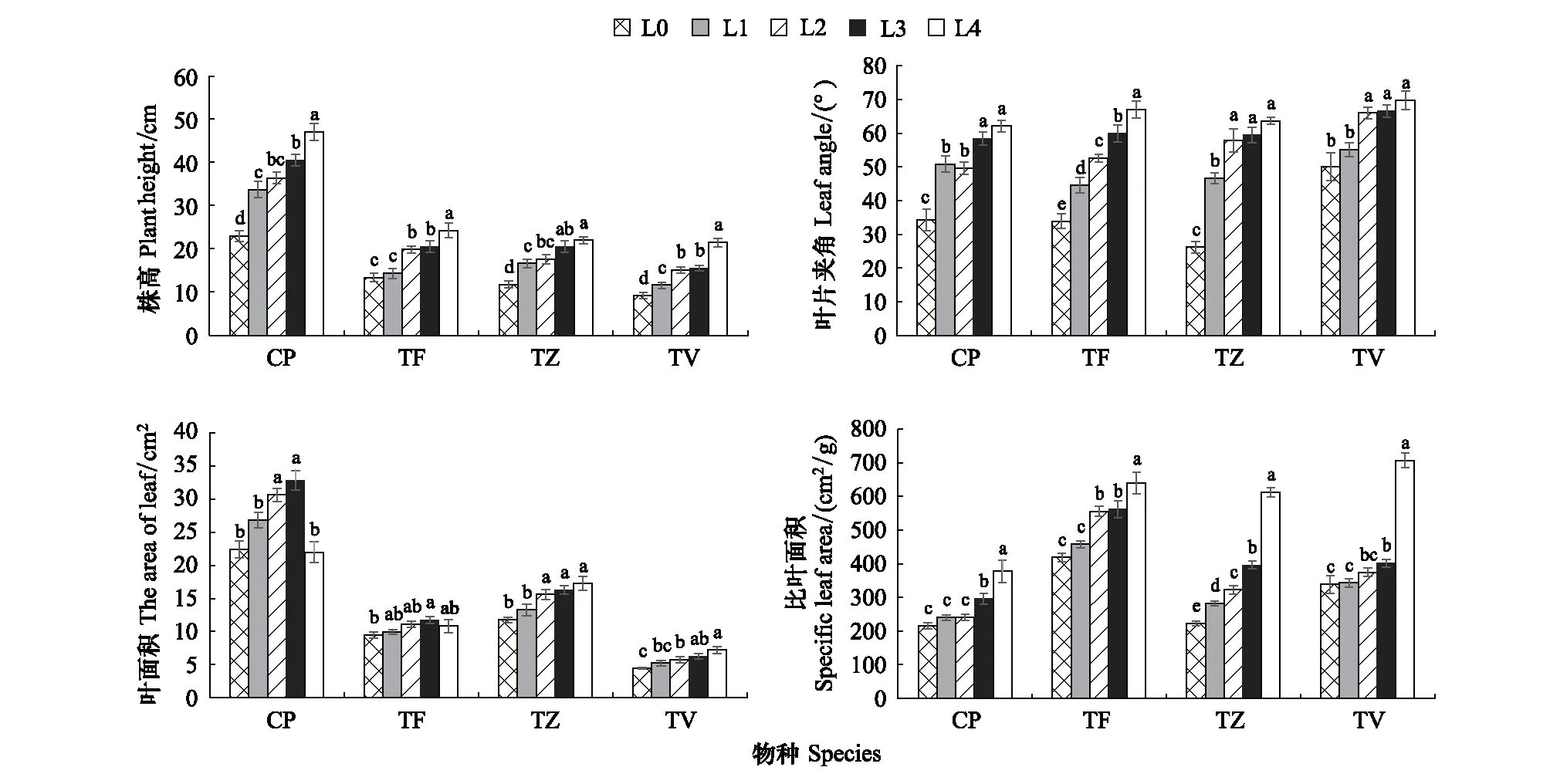
图2 不同光强对形态指标的影响
4种植物的叶面积在不同光强下呈现不同的变化规律,CP的叶面积在L0—L3条件下显著增加(P<0.05),并在L3条件下达到峰值32.88,而在L4条件下显著降低(P<0.05),这表明CP的叶面积指标在适应光强变化时存在一定的局限性;TF叶面积在L1—L4光强下无显著差异,但在L3条件下达到峰值,与L0条件下的最低值相比显著(P<0.05)增加了24.26%;而随光强的降低,TZ和TV的叶面积显著升高(P<0.05),均在L4条件下达到最大值,与L0相比分别显著增加了0.46倍和0.64倍(P<0.05)。
比叶面积是评价植物耐阴能力的重要指标之一[20], CP、TF、TZ、TV的比叶面积均随光强的降低而显著升高(P<0.05),在L4下达到峰值,与L0下的最小值相比,分别增加了0.75倍、0.53倍、1.75倍、1.08倍。
2.2 不同光强对叶片解剖结构的影响
由图3可知,4种植物的叶片解剖结构从上至下依次为上表皮、栅栏组织、海绵组织和下表皮,上表皮由1—2层长椭圆形的细胞组成,切面面积较大;下表皮由一层形状不规则的细胞构成,细胞排列紧密,横截面面积相对较小。CP、TF、TZ、TV的叶片厚度、上表皮厚度、下表皮厚度均与光强正相关(图4),且均在L0下达到最大值,在L4条件下降到最低值。
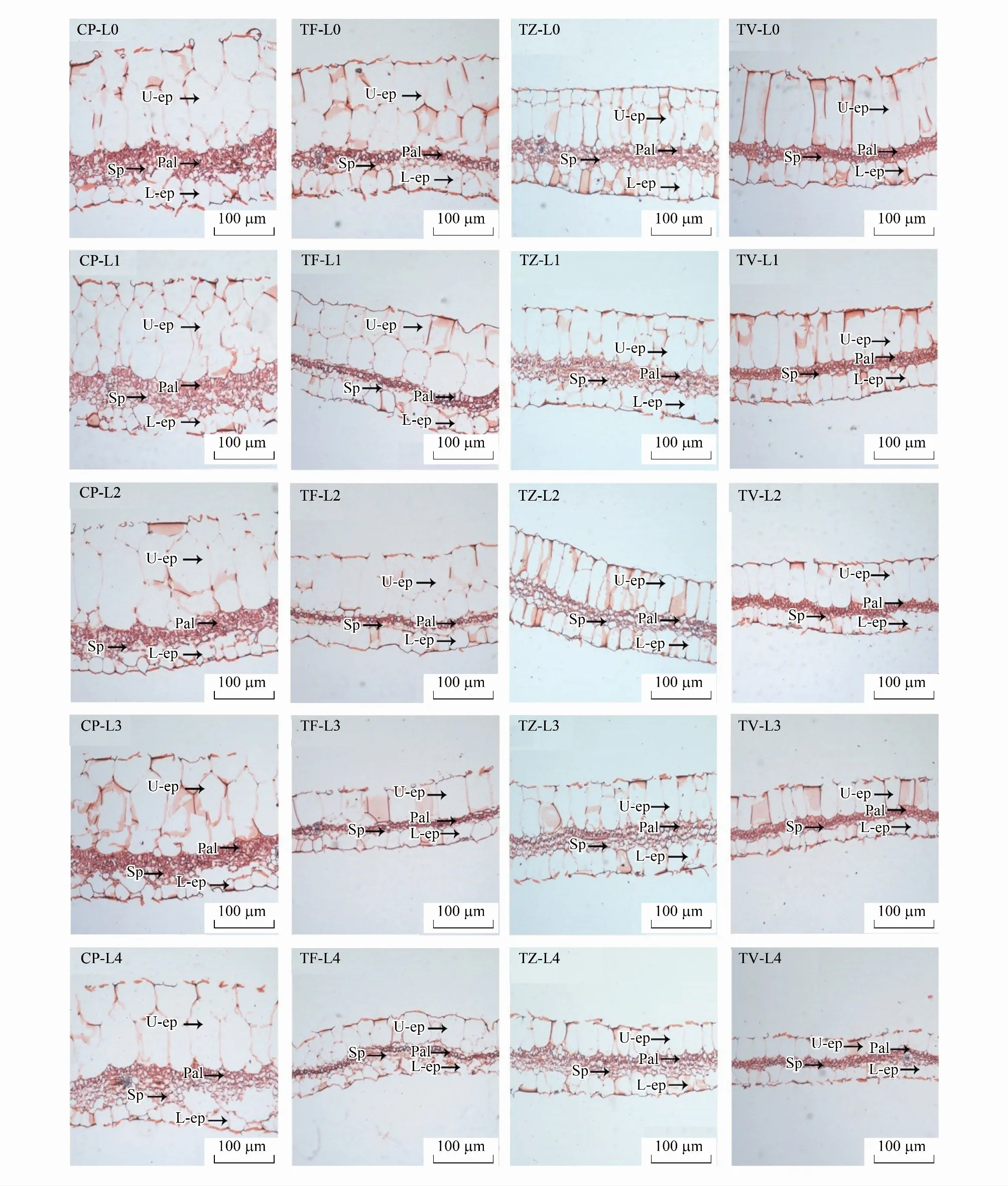
图3 不同光强对叶片解剖结构特征的影响
4种植物的叶肉解剖结构指标在不同光强下产生了明显的变化(图3和图4):即随光强降低,CP的栅栏组织厚度显著下降了0.29倍(P<0.05),且3层栅栏组织也变为2层,构成栅栏组织的细胞纵横轴比减小,由长椭圆形变为短椭圆形。不同的光强虽然没有导致海绵组织厚度产生显著变化,但海绵组织细胞排列逐渐松散,组织细胞间隙变大;TF的栅栏组织和海绵组织厚度均与光强正相关,与L0下的最大值相比,L4条件下的栅栏组织和海绵组织厚度分别显著降低了0.46倍、0.37倍(P<0.05),2层栅栏组织细胞减少为1层,且组织细胞松散,纵横轴比减小,但海绵组织细胞形态与分布没有明显的变化;TZ的栅栏组织和海绵组织厚度没有随光强的降低而发生显著的变化,但栅栏组织形态发生了明显的变化,组织细胞排列逐渐松散;TV的栅栏组织和海绵组织厚度与光强正相关,与L0相比,L4下栅栏组织和海绵组织厚度分别显著降低了0.35倍、0.37倍(P<0.05),栅栏组织也由2层变为1层,但组织细胞在不同光强下始终排列紧密,与其余3种植物相同的是,TV的海绵组织也随光强的降低而变得松散。值得注意的是,在不同的光强下,4种植物的栅栏组织和海绵组织的比值均没有发生显著变化,这表明4种植物的叶肉解剖结构具有较强的稳定性。
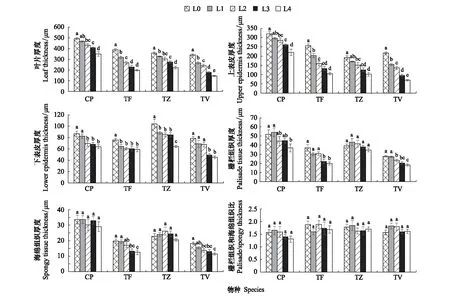
图4 不同光强对叶肉结构指标的影响
2.3 不同光强对叶片气孔开度的影响
随光强的降低,4种植物气孔开度的变化趋势较为一致(图5和图6),呈先增后减的变化趋势且均在L0下最小,但达到峰值时的光强不同,CP、TF、TZ和TV分别在L3、L2、L3和L1下达到峰值,与各自在L0下的最低值相比,分别显著增加了1.25倍、0.83倍、0.34倍、0.34倍(P<0.05),可见CP的气孔开度对光强的变化最为敏感。

图5 不同光强下气孔的变化
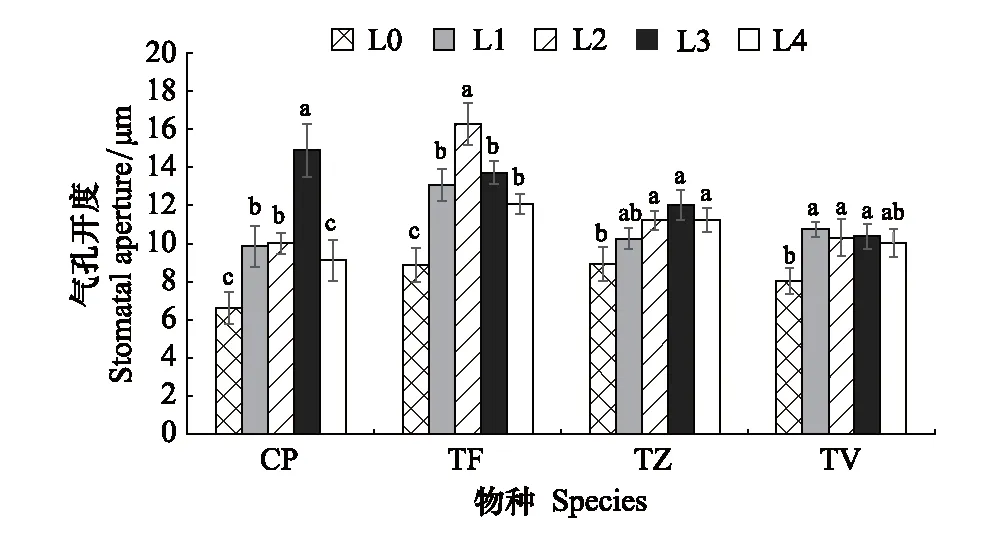
图6 不同光强对气孔开度的影响
2.4 不同光强对叶绿素含量的影响
4种植物在不同光强下的叶绿素含量均有显著的变化(图7)(P<0.05)。CP和TF的Chl a、Chl b和Chl (a+b)含量随光强的降低呈先增后减趋势,但2种植物的Chl a、Chl b和Chl (a+b)在L4下的含量均大于L0。不同的是CP的Chl a、Chl b和Chl (a+b)的含量在L3下达到最大值,分别比L0下的最小值显著增加了0.68倍、0.26倍、0.56倍(P<0.05),而TF的Chl a、Chl b和Chl (a+b)的含量在L2下最大,分别比L0下的最小值显著增加了1.19倍、0.70倍、1.08倍(P<0.05);而TZ和TV的Chl a、Chl b和Chl (a+b)含量与光强负相关,TZ在L4下的Chl a、Chl b和Chl (a+b)含量与L0下的最低值相比分别显著增加了1.95倍、1.60倍、1.72倍(P<0.05),TV在L4下的Chl a、Chl b和Chl (a+b)含量与L0下的最低值相比分别显著增加了3.20倍、2.67倍、3.03倍(P<0.05); CP、TF、TZ 和TV的Chl a/b与光强正相关,虽然在随光强的降低过程中有小范围的降低趋势,但未达到显著水平。
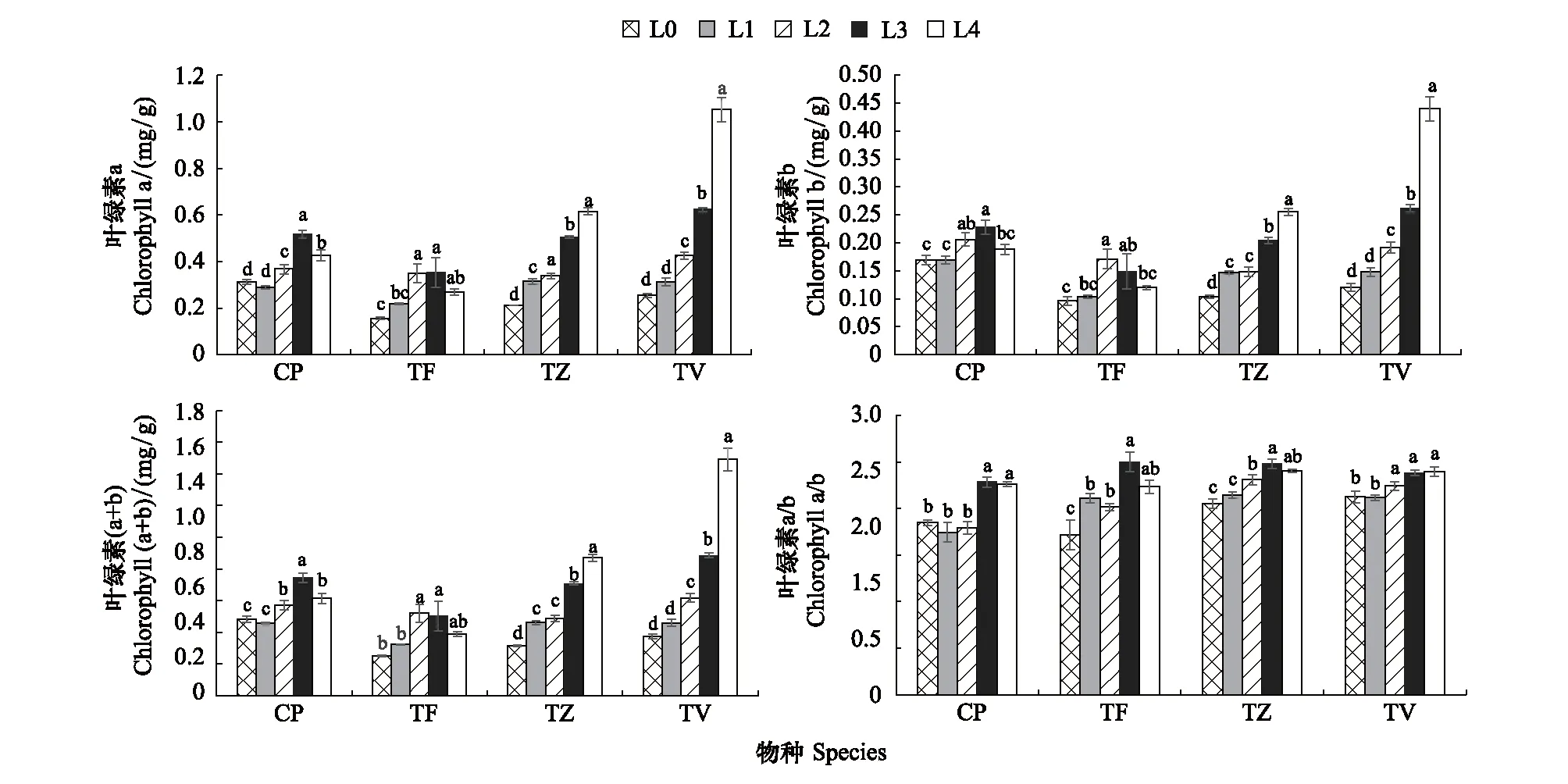
图7 不同光强对叶绿素含量的影响
2.5 不同光强对光合生理的影响
不同光强下,CP、TF、TZ和TV的Pn和Tr的变化规律基本一致,均先升高后降低(图8),且4种植物的Pn和Tr均在L4下降到最低。不同的是4种植物的Pn分别在L1、L1、L2和L2下达到峰值,与L4的最低值相比,CP、TF、TZ和TV的Pn分别显著升高了0.65倍、1.40倍、0.73倍、1.25倍(P<0.05),而4种植物的Tr分别在L2、L1、L2、L3下达到峰值,与L4下的最低值相比,分别显著降低了0.27倍、0.31倍、0.36倍、0.27倍(P<0.05)。
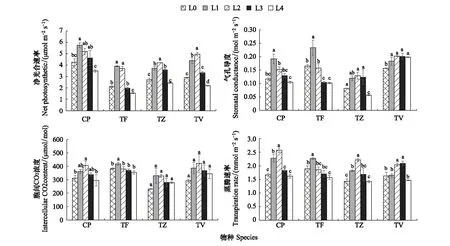
图8 不同光强对光合指标的影响
随着光强的降低,CP、TF、TZ和TV的Gs均先升高后降低,分别在L1、L1、L2和L2下达到峰值,其中CP、TF和TZ的Gs在L4下降到最小值,与峰值相比分别显著降低了0.42倍、0.57倍、0.54倍(P<0.05),而TV在L2—L4下有降低的趋势,但差异不显著,且在L0下降到最低值。
随光强的降低,TZ和TV的Ci无显著变化,而CP和TF的Ci则呈先升高后降低的趋势,且分别在L2、L1下达到峰值,与L4下的最小值相比分别显著降低了0.27倍、0.15倍(P<0.05)。
2.6 物种和光强对不同指标的双因素方差分析
方差分析表明(表1),物种因素对株高、叶片夹角、叶面积、叶片厚度、比叶面积、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度、气孔开度、Chl a、Chl b、Chl (a+b)、Chl a/b、Pn、Gs、Ci和Tr具有极显著影响(P<0.01),对栅栏组织和海绵组织比具有显著影响(0.01
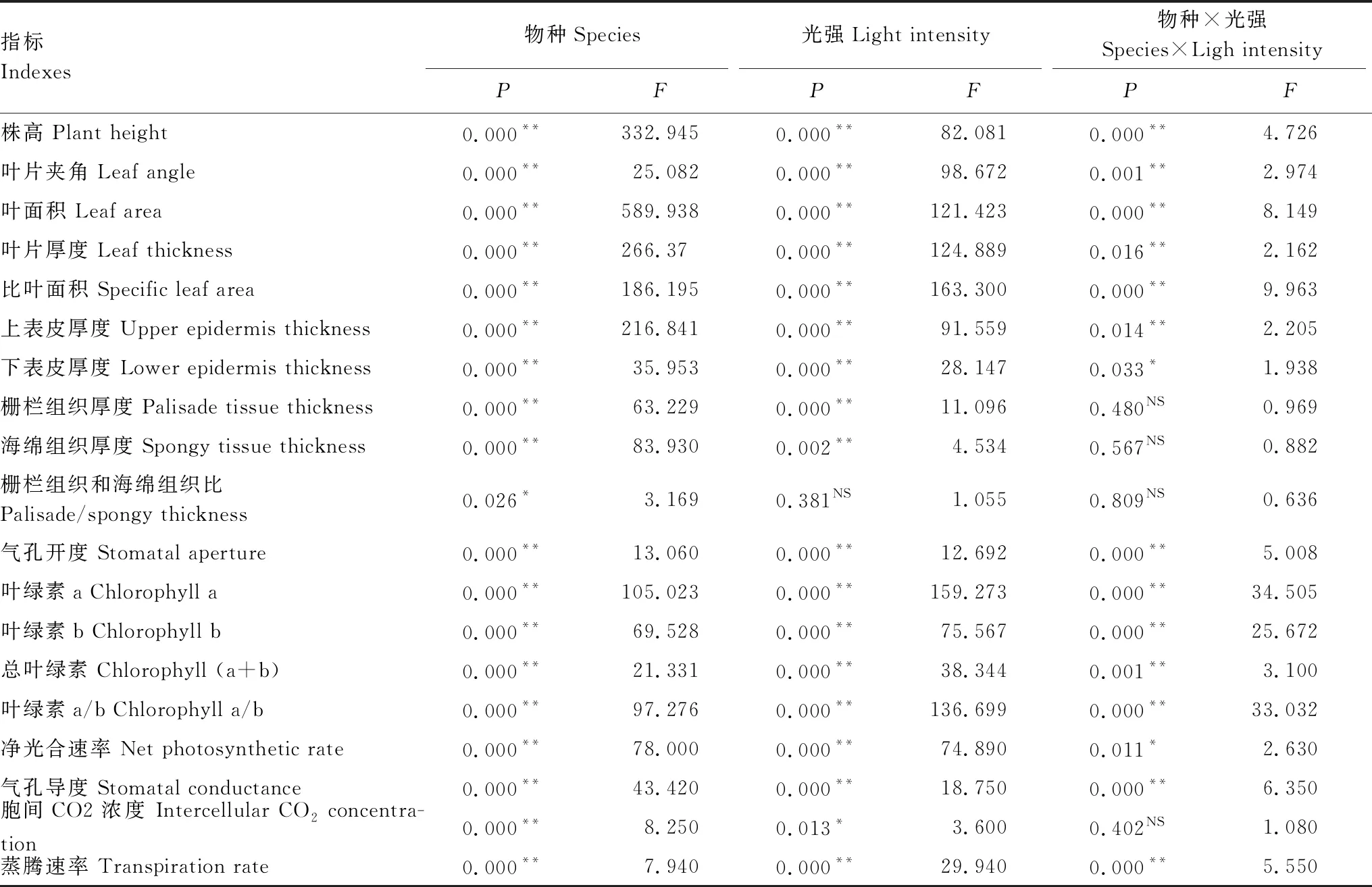
表1 物种和光强对不同指标的双因素方差分析
3 讨论
光强是重要的环境因子,与植物的生长发育息息相关。叶片作为植物的六大器官之一,是进行光合作用的主要场所。由于叶片是植物体感受外界环境的最大的器官,其形态结构对于生态环境的改变非常敏感,因此形态结构特征和生理特性的变化最能反应光环境对植物的影响以及植物对光环境的适应能力。相关研究表明:大多数植物在弱光环境中能够改变叶片形态,如通过增加叶长、叶宽、叶面积、叶片数量和株高的方式适应光强不足的环境[21—24],而在强光下则会通过增加叶片厚度、分泌蜡质层和减小叶面积的方式应对强光抑制[10—11, 25]。本研究发现4种植物的株高、叶片夹角、叶面积、比叶面积,花叶水竹草、吊竹梅和绿叶水竹草的叶面积随光强的降低而显著增加,紫鸭跖草的叶面积在L0—L3下显著升高,而L4下叶面积则显著降低。这表明当外界自然光强减弱时,为了捕获更多有限的光能,4种植物增加了植株高度、扩大了叶片面积、增大了叶片夹角,与此同时为了降低植株体营养物质的消耗,4种植物通过增加比叶面积,在有限的范围内牺牲叶片厚度扩大叶面积以增强自身对光能的捕获效率,补偿较低光量子通量密度造成的光合速率下降的现象,从而达到在弱光环境中生存的目的,这与Moreira[26]的观点一致。而紫鸭跖草的叶面积在L4条件下显著降低,这表明其叶片形态可塑性在弱光环境存在明显的阈值。
叶肉是进行光合作用的主要场所,叶肉解剖结构的变化必然会影响植物的光合效率[27]。研究发现,较薄的上下表皮和栅栏组织厚度、具有较低纵横轴比的栅栏组织细胞以及排列松散的海绵组织细胞能提高光的透射率,增加叶绿体对光的捕获能力[28—29]。在对4种鸭跖草科植物的研究中,进一步印证了这一结论,即随着光强的降低,4种植物的叶片厚度,上下表皮厚度,紫鸭跖草、花叶水竹草和绿叶水竹草的栅栏组织厚度、花叶水竹草和绿叶水竹草的海绵组织厚度均显著降低,且上下表皮细胞以及栅栏组织细胞的纵横轴比降低,海绵组织细胞间的排列逐渐松散,间隙逐渐变大。与这3种植物不同的是,不同光强下吊竹梅的栅栏组织和海绵组织厚度未发生显著的变化,这表明不同的植物在适应不同的光环境时叶肉结构的适应策略存在明显的差异。值得注意的是,在不同的光强下4种植物的栅栏组织和海绵组织比没有发生显著的变化,这表明减弱的光强虽然使得4种植物的叶片厚度显著降低,但栅栏组织和海绵组织结构能保持较高的稳定性,从而维持叶片正常的生理机能。
Chl a、Chl b是主要的光合色素,其含量的高低和比例是植物应对环境胁迫的重要指标[30]。在本研究中,紫鸭跖草和花叶水竹草在L0—L3下Chl a、Chl b和Chl (a+b)含量显著增加,虽然在L4下有降低的趋势,但其含量仍大于L0条件,吊竹梅和绿叶水竹草的Chl a、Chl b和Chl (a+b)含量则随着光强的降低而显著升高,这表明这4种植物在各种弱光环境下均能够通过增加叶绿素含量的方式提高捕获光辐射的效率,以适应不同的弱光环境。CP、TF、TZ 和TV的Chl a/b则与光强正相关,虽然Chl a/b在随光强的降低过程中有小幅度的降低趋势,但未达到显著水平,且4种植物的Chl a/b均一直小于3.2,这表明它们都属于典型的阴生植物[31],对生长环境中光强的要求不高。
通常植物在逆境胁迫下,光合速率的下降主要是由于气孔限制和非气孔限制导致的[32]。气孔限制是由于气孔导度下降,外界环境中的CO2被限制通过叶片气孔进入植株体内,从而导致光合速率降低。而非气孔限制导致光合速率降低的情况主要是由于胁迫导致光合相关酶活性降低,光合器官受损造成的,此时气孔导度和胞间二氧化碳可能仍保持较高水平。在本研究中,随着光强的降低,4种植物Pn的变化趋势与气孔开度、Gs和Ci的变化趋势基本保持一致,即呈先升高后降低趋势,且在100%自然光强或5%自然光强下降到最低值,这表明4种植物的光合速率的变化主要是由于气孔限制所导致,在100%自然光强下,植株体产生了光抑制现象,为了减少水分的散失,叶片气孔关闭,降低了蒸腾速率,与此同时也限制了植物对CO2的吸收,从而减少了对植物光合作用时底物的供应,最终导致光合速率的下降[7],而随着光强的减弱,气孔逐渐开放,外界的CO2通过气孔进入植株体内,不断供给光合作用所需的原料,从而促进光合速率的提高,当光强只有5%自然光强时,由于过度阴蔽,植株体产生了光亏缺,无法获得生命活动的所必须的光能源,导致光合速率降到了最低。
4 结论
本研究表明,4种鸭跖草科植物的光合速率主要受“气孔因素”影响,能在25%—75%自然光强下保持较高的净光合速率,并且还通过能增加叶面积、植株高度、叶片夹角度数、比叶面积、叶绿素含量,降低上表皮厚度、下表皮厚度、栅栏组织厚度以及整个叶片的厚度的方式来获取和利用有限的光资源,提高光的利用率,降低养分消耗,以维持自身良好的生长状态,而在100%自然光强和5%自然光强时分别产生了光抑制和光亏缺现象,不利于植株体的正常生长。
