盐胁迫对3种平欧杂种榛幼苗叶片解剖结构及离子吸收、运输与分配的影响
2022-03-30吴正保史彦江宋锋惠
罗 达,吴正保,史彦江,宋锋惠,*
1 新疆林业科学院经济林研究所,乌鲁木齐 830063 2 新疆林木资源与利用国家林草局重点实验室,乌鲁木齐 830063 3 新疆林果树种选育与栽培重点实验室,乌鲁木齐 830063
土壤盐渍化是自然界的主要灾害之一,也是威胁农作物生长最严重的非生物胁迫之一。我国土壤盐渍化问题尤为突出,主要发生在西北、华北和东北干旱半干旱地区[1]。盐胁迫对植物生长发育的各个阶段(如种子萌发、生长、开花结实等)以及生理代谢的各个过程(如水分代谢、光合作用、酶系统代谢等)都造成影响,进而使农作物产量和品质下降,严重时甚至颗粒无收。因此,为了提高盐渍化土地资源的利用率,保障生态安全并实现农林业优质高产,开展植物耐盐机理研究、培育耐盐植物品种显得尤为迫切和重要。
植物在遭受盐胁迫时,通常会在根、茎、叶等形态结构上表现出一系列的适应性特征。叶片作为植物生长发育过程中可塑性较强的器官,其形态结构特别是解剖结构特征的变化与植物的耐盐性密切相关[2]。研究表明,随着盐胁迫程度的增强,耐盐性强的象草(Pennisetumpurpureum)叶片厚度,上、下表皮厚度呈先上升后下降的趋势[3];葡萄(Vitisvinifera)叶表皮厚度、栅栏组织和海绵组织厚度显著增加,栅栏组织/海绵组织比显著降低[4]。盐碱环境下的杏(Prunusarmeniaca)表现出叶片肉质化,角质层增厚,栅栏组织发达,气孔下陷等典型的耐盐碱特征[5]。盐胁迫下,沼泽小叶桦(Betulamicrophyllavar.paludosa)叶片栅栏组织厚度[6]和紫花苜蓿(Medicagosativa)叶片栅栏组织/海绵组织比[7]增大。由此可见,开展盐碱胁迫下的叶片解剖结构特征研究有助于进一步揭示植物对盐碱环境的形态响应与适应机制。
盐胁迫下,以Na+为首的盐离子通过直接干扰或竞争性地抑制细胞质膜对K+、Ca2+、Mg2+等营养离子的吸收和转运,造成植物体内的离子失衡、离子毒害和营养亏缺,进而影响植物的正常生长发育[8—10]。植物可以通过细胞膜上的Na+/H+逆向转运蛋白将胞质中的Na+外排以维持胞质中低水平的Na+,或是通过液泡膜上的Na+/H+逆向转运蛋白将Na+区隔于液泡中,从而避免Na+带来的离子胁迫并保持胞内离子平衡[11];也可通过减少体内K+、Ca2+、Mg2+等营养离子的流失来提高其保有能力,进而提高耐盐性[12—14]。Yang等[15]认为植物维持适宜的细胞质K+/Na+比值是耐盐植物对过量离子反应的重要适应性状。研究表明,不同品种或种源植物的离子代谢特征对盐胁迫的响应和适应存在差异。盐胁迫下,蓝莓(Vacciniumcorymbosum)‘蓝丰’幼苗根、茎、叶对Na+和Cl-的吸收低于蓝莓‘奥尼尔’,对Mg2+的吸收则高于‘奥尼尔’,其维持体内离子平衡的能力以及K+和Mg2+的整体离子选择性运输能力强于‘奥尼尔’,耐盐性也相对较强[16]。与银川种源沙枣(Elaeagnusangustifolia)相比,阿拉尔种源沙枣(Elaeagnusangustifolia)能更有效地将Na+阻隔于根系中,减少向地上部分运输,并能更好地减少幼苗各组织中K+、Ca2+和Mg2+的流失,进而维持离子平衡以保证体内各生理代谢过程的正常进行[17]。
平欧杂种榛(Corylusheterophylla×Corylusavellan)是由辽宁省经济林研究所利用原产于我国的平榛(C.heterophylla)与外来的欧洲榛(C.avellan)种间杂交选育而来,因其兼具平榛(耐寒、耐瘠薄、适应性强)与欧洲榛(果大、壳薄、丰产)的优良遗传特性[18],是目前我国广泛推广栽培的榛树品种。新疆自2001年开始开展榛子的区域化引种试验,经过20年的引种、驯化、选育等一系列研究工作,现已筛选出‘达维’、‘辽榛7号’、‘玉坠’、‘辽榛3号’、‘平欧15号’等几个适宜推广栽培的优良品种。土壤盐渍化问题一直是制约农林业开发及可持续发展的重大限制条件和障碍因素。目前,国内外仅在生长和生理生态特性[19—21]等方面对平欧杂种榛的耐盐性开展过相关研究,如盐胁迫下,‘达维’、‘辽榛7号’、‘玉坠’3个品种平欧杂种榛幼苗的新梢生长量、根、茎、叶生物量以及幼苗总生物量显著降低,根冠比则随着盐胁迫的增强而增加,且不同品种的盐胁迫响应差异明显[21]。而对盐胁迫下不同品种的叶片解剖结构以及体内离子平衡的盐适应机制和响应差异的研究尚未探及。本文以不同品种平欧杂种榛幼苗为试材,采用盆栽控盐对比试验,研究盐胁迫下幼苗叶片显微解剖结构以及根、茎、叶中Na+、K+、Cl-、Ca2+的吸收、运输和分配特征,以期探明平欧杂种榛的盐胁迫响应与适应机制及不同品种的耐盐性差异。
1 材料与方法
1.1 试验材料
供试材料为3个优良品种的平欧杂种榛:‘达维’、‘辽榛7号’、‘玉坠’,苗木为2年生压条根蘖苗,由伊犁琨源农林开发有限责任公司提供。
1.2 试验设计
试验在新疆林业科学院树木实验园内进行,试验期间采用美国HOBO小型自动气象站监测6—9月的日平均气温为15.3—28.2℃,相对湿度20.2%—44.3%,太阳辐射125.6—434.4 W/m2。为确保苗木一致性,选择地径和株高基本一致的幼苗进行盆栽试验。将苗木定植在40 cm×35 cm的白色塑料盆钵中,每盆定植1株,之后将苗木统一截干至10 cm高,供试基质为经自来水和蒸馏水冲洗过的蛭石。
幼苗定植后,对其进行日常浇水管理,待新梢长至20 cm左右时开始进行盐胁迫处理。盐胁迫处理前4 d停止浇水,以保证盐溶液在基质中均匀扩散。试验采用随机区组试验设计,根据张丽[20]的试验结果,本研究设置3个盐浓度梯度的胁迫水平,即轻度(50 mmol/L NaCl)、中度(100 mmol/L NaCl)、重度(200 mmol/L NaCl),设对照为0。盐胁迫处理分3个试验区组,每个试验区组每种盐胁迫处理12盆,共计144盆。所用盐溶液为相应质量的NaCl溶于1/2 Hoagland营养液配置而成,每盆植株浇灌1.5 L相应浓度的处理液。为了防止盐溶液对苗木的盐冲击效应,盐溶液分3次浇灌完成,胁迫第1 d用500 mL相应浓度的处理液浇灌1次,第2、3 d各补500 mL。盐胁迫期间根据盆内基质水分状况适时补水(去离子水),每次补水以充分灌透且溶液刚刚渗出为宜,期间每20 d用不加NaCl的1/2 Hoagland营养液浇灌1次,以保证营养供应。试验期间,盆底垫塑料托盘,以保证补水和浇灌营养液时渗出的盐分及时返还盆中,确保盆中盐分总量。试验期间对供试苗木进行日常养护管理。
1.3 测定指标与方法
1.3.1样品采集与处理
盐胁迫处理4个月后,随机选取各处理试验苗木9株作为样株。为确保试验结果的一致性,采集样株一年生枝条由上往下第3—4枚健康成熟叶片足够量,立即放入提前配置好的FAA固定液(70%乙醇90 mL+甲醛5 mL+乙酸5 mL)中,并迅速带回实验室用于叶片解剖结构分析。之后,分别采集幼苗根、茎、叶样品,装入信封后编号带回实验室用于离子含量测定。
1.3.2叶片解剖结构
参照徐青[22]的方法,采用改进后的双重染色技术对叶样进行石蜡切片分析。将固定好的叶片,经各个梯度酒精(70%、85%、95%和100%)脱水、番红染色、二甲苯透明、石蜡包埋切片(厚度6—8 μm)、番红-固绿双重染色,中性树胶封片,于显微镜相连的Nikon照相机下观察、拍照。利用显微镜对各处理的样叶切片依次观测以下叶片解剖结构指标:叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度,所有叶片各指标观测10—15个视野后取平均值。计算栅栏组织厚度/海绵组织厚度比值,并按下列公式计算结构紧密度和疏松度:
结构紧密度=栅栏组织厚度/叶片厚度
结构疏松度=海绵组织厚度/叶片厚度
1.3.3离子代谢特征
取幼苗根、茎、叶样品一定量,经105 ℃烘箱杀青20 min后,于80 ℃烘干至恒质量,磨碎过40目筛后各称取1.0 g,加入30 mL去离子水,摇匀后沸水浴中放置2 h,冷却后定容至50 mL备测。其中,Na+、K+、Ca2+含量采用原子吸收光谱仪(PerkinElmer PinAAcle 900T)测定;Cl-含量采用电位滴定仪(ZDJ-4A)测定。计算各器官的K+/Na+和Ca2+/Na+比值,并按下列公式计算不同器官对离子的吸收和选择性运输能力(SX,Na+)[23]:
SX,Na+=库器官[X/Na+]/源器官[X/Na+]
式中,X为K+含量或Ca2+含量,SX,Na+值越大表示源器官控制Na+、促进X向库器官的运输能力越强,即库器官的选择性运输能力越强。
1.4 数据处理
采用Excel 2007和SPSS 22.0软件对数据进行统计分析。采用单因素(one-way ANOVA)和Duncan法进行方差分析和多重比较(α=0.05)。利用Origin Pro 9.0软件完成绘图。图中数据为平均值±标准差。
2 结果与分析
2.1 盐胁迫对叶片解剖结构的影响
图1可见,平欧杂种榛为异面叶,平均厚度约195.46 μm。叶片由上表皮细胞、下表皮细胞,栅栏组织和海绵组织构成。表皮细胞排列紧密,其中,上表皮细胞呈长形至椭圆形,平均厚度约14.16 μm;下表皮细胞小而不规则,平均厚度约10.87 μm。叶肉中栅栏组织呈长柱形,排列整齐、紧凑,平均厚度约107.96 μm;海绵组织形状不规则,排列疏松,细胞间隙较大,平均厚度约50.85 μm。

图1 盐胁迫下‘辽榛7号’叶片解剖结构图
从图2可以看出,盐胁迫对平欧杂种榛幼苗叶片各解剖结构参数均产生不同程度的影响。不同品种叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度均随着盐胁迫程度的增强呈现出先增加后降低的特点。其中,轻度和中度胁迫下,不同品种叶片各解剖结构参数均显著高于对照(P<0.05);重度胁迫下,‘达维’叶片上表皮厚度显著低于对照(P<0.05),‘辽榛7号’叶片厚度、上表皮厚度、栅栏组织厚度和海绵组织厚度均显著高于对照(P<0.05),‘玉坠’叶片上表皮厚度、栅栏组织厚度显著高于对照(P<0.05),其余叶片解剖结构参数均降低至对照水平(P>0.05)。轻度和中度盐胁迫下,‘达维’、‘玉坠’叶片栅栏组织/海绵组织比显著提高(P<0.05),不同程度盐胁迫对‘辽榛7号’的影响不显著(P>0.05)。中度盐胁迫显著提高了不同品种叶片的结构紧密度(P<0.05)。与对照相比,轻度和中度盐胁迫下,‘达维’、‘玉坠’叶片结构疏松度显著降低(P<0.05),‘辽榛7号’则显著提高(P<0.05)。从不同品种来看,盐胁迫下叶片各解剖结构参数也存在不同程度的差异。轻度胁迫下,叶片厚度、上表皮厚度、下表皮厚度表现为‘达维’>‘玉坠’>‘辽榛7号’;中度胁迫下,栅栏组织厚度、海绵组织厚度、结构紧密度均表现为‘辽榛7号’>‘玉坠’>‘达维’。
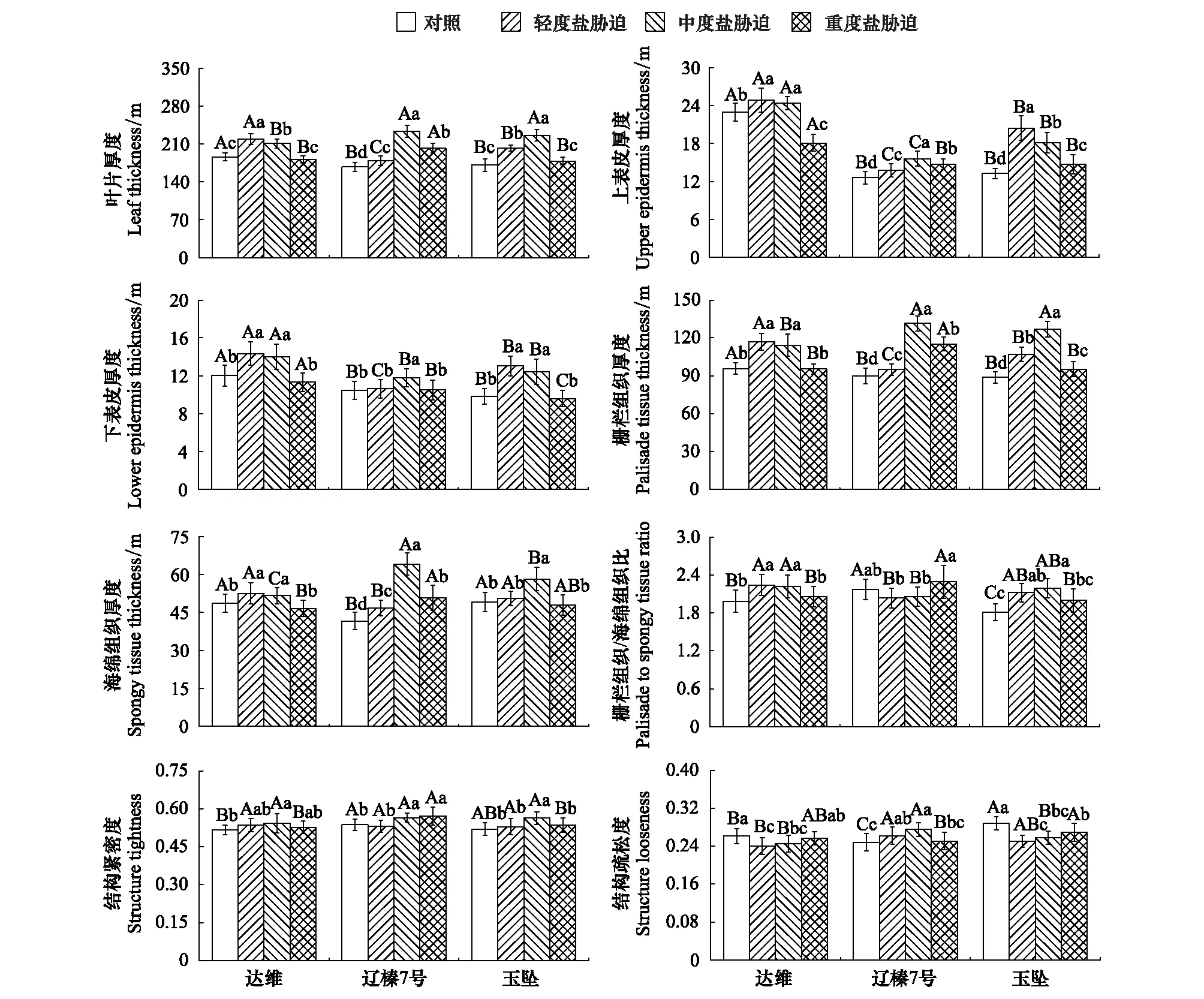
图2 盐胁迫下平欧杂种榛幼苗叶片解剖结构参数
2.2 盐胁迫对离子含量的影响
2.2.1盐胁迫对Na+含量的影响
从图3可知,盐胁迫显著影响平欧杂种榛幼苗根、茎、叶Na+含量(P<0.05)。随着盐胁迫程度的增强,不同品种各器官Na+含量呈递增趋势。不同程度盐胁迫下,叶Na+含量始终高于根和茎。但从Na+增幅来看,轻度、中度和重度胁迫下3个品种根Na+含量分别平均提高1.14、2.24、4.35倍,茎Na+含量分别平均提高0.75、2.83、4.15倍,高于叶Na+含量增幅的0.26、0.76、1.42倍,表明盐胁迫处理后根开始积聚大量Na+。从不同品种来看,同一器官Na+含量也存在明显差异。轻度、中度和重度胁迫下,‘辽榛7号’根、茎、叶Na+含量及增幅均低于‘达维’和‘玉坠’,表明盐胁迫对‘辽榛7号’Na+吸收的影响小于其它2个品种。

图3 盐胁迫下平欧杂种榛幼苗根、茎、叶中Na+含量
2.2.2盐胁迫对Cl-含量的影响
图4显示,盐胁迫显著影响平欧杂种榛幼苗根、茎、叶Cl-含量。随着盐胁迫程度的增强,不同品种各器官Cl-含量呈明显增加趋势,并在重度胁迫下达到最大值。不同程度盐胁迫下,叶Cl-含量最高,其次为茎,根最低。但从Cl-增幅来看,轻度、中度和重度胁迫下3个品种根Cl-含量分别平均提高0.53、1.13、1.59倍,高于茎Cl-含量增幅的0.39、0.88、1.32倍,且明显高于叶Cl-含量增幅的0.16、0.41、0.77倍,表明盐胁迫处理后根系开始大量积累Cl-,并在之后将其转运至茎和叶中。从不同品种来看,同一器官Cl-含量也存在不同程度的差异。不同程度盐胁迫下,‘辽榛7号’根、茎、叶Cl-含量及增幅总体低于‘达维’和‘玉坠’。

图4 盐胁迫下平欧杂种榛幼苗根、茎、叶中Cl-含量
2.2.3盐胁迫对K+含量的影响
从图5可以看出,盐胁迫下不同器官对K+的吸收规律存在差异。随着盐胁迫程度的增强,不同品种根、茎K+含量呈先降低后略有回升的趋势。轻度、中度和重度胁迫下,根K+较对照分别显著降低18.9%、44.0%和25.7%(P<0.05),茎K+较对照分别显著降低22.1%、40.7%和27.2%(P<0.05)。盐胁迫下,叶K+呈先增加后急剧下降的趋势,轻度、中度胁迫下叶K+较对照分别增加20.3%和40.3%(P<0.05),重度胁迫较对照降低29.4%。总体来看,盐胁迫抑制了各品种根、茎K+的吸收,但适度盐胁迫促进了K+在叶片中的积累,这对于植株维持正常的生理活动具有重要作用。不同品种间,各器官K+含量总体表现为‘辽榛7号’>‘玉坠’>‘达维’。同时,‘辽榛7号’根、茎K+的降幅始终低于‘达维’和‘玉坠’,叶K+的增幅则始终高于‘达维’和‘玉坠’。

图5 盐胁迫下平欧杂种榛幼苗根、茎、叶中K+含量
2.2.4盐胁迫对Ca2+含量的影响
图6表明,盐胁迫下,不同器官对Ca2+的吸收规律不同。其中,根、茎Ca2+随着盐胁迫程度的增强表现为先降低后升高,轻度胁迫下根、茎Ca2+(‘辽榛7号’茎除外)与对照无显著差异(P>0.05),中度胁迫下根、茎Ca2+较对照分别显著降低21.8%和46.7%(P<0.05)。轻度胁迫显著提高了‘达维’叶Ca2+(P<0.05),对‘辽榛7号’和‘玉坠’叶Ca2+的影响不显著(P>0.05);重度胁迫显著抑制了各品种叶对Ca2+的吸收(P<0.05)。从不同品种来看,同一器官Ca2+含量也存在显著差异(P<0.05)。不同程度盐胁迫下,‘辽榛7号’根、茎、叶Ca2+含量总体高于‘达维’和‘玉坠’,表明盐胁迫下‘辽榛7号’维持Ca2+吸收的能力强于其它2个品种。

图6 盐胁迫下平欧杂种榛幼苗根、茎、叶中Ca2+含量
2.3 盐胁迫对离子比值的影响
图7显示,盐胁迫对不同品种平欧杂种榛幼苗各器官K+/Na+比和Ca2+/Na+比均产生显著影响(P<0.05)。总体来看,盐胁迫导致不同品种各器官K+/Na+比和Ca2+/Na+比显著降低(P<0.05)。从不同品种来看,各器官K+/Na+比和Ca2+/Na+比在品种间也存在显著差异(P<0.05),具体表现为‘辽榛7号’>‘玉坠’>‘达维’,而盐胁迫增强时K+/Na+比和Ca2+/Na+比的降幅为‘达维’>‘玉坠’>‘辽榛7号’,表明盐胁迫下‘辽榛7号’维持体内离子平衡的能力最强,其次为‘玉坠’,‘达维’最弱。

图7 盐胁迫下平欧杂种榛幼苗根、茎、叶K+/Na+、Ca2+/Na+
2.4 盐胁迫对离子选择性运输能力的影响
从图8可以看出,‘达维’从根到茎的SK+,Na+和SCa2+,Na+随着盐胁迫程度的增强呈升-降-升的趋势。盐胁迫对‘辽榛7号’和‘玉坠’从根到茎的SK+,Na+影响不显著(P>0.05),中度和重度胁迫下‘辽榛7号’和‘玉坠’从根到茎的SCa2+,Na+显著降低(P<0.05)。随着盐胁迫程度的增强,不同品种平欧杂种榛从茎到叶以及从根到叶的SK+,Na+和SCa2+,Na+均表现为先升高后降低的特点,并在轻度或中度胁迫下达到最大值,重度胁迫下显著高于或维持在对照水平。从不同品种来看,各器官间的SK+,Na+和SCa2+,Na+在品种间也存在差异,具体表现为从根到茎、从茎到叶以及从根到叶的SK+,Na+在不同程度盐胁迫下的均值表现为‘辽榛7号’>‘达维’>‘玉坠’,SCa2+,Na+则表现为‘辽榛7号’>‘玉坠’>‘达维’。

图8 盐胁迫对平欧杂种榛幼苗SK+,Na+、SCa2+,Na+的影响
3 讨论
3.1 叶片解剖结构特征对盐胁迫的响应与适应
盐碱生境下的植物通常具有一些特殊的叶片解剖结构特征,如角质层增厚,叶片肉质化,栅栏组织发达,气孔下陷,叶片有盐腺分布等[2,24]。本研究中,不同品种平欧杂种榛叶片厚度、上表皮厚度、下表皮厚度、栅栏组织和海绵组织厚度均随着盐胁迫程度的增强呈先增加后降低的特点,轻度、中度胁迫下各参数显著提高,重度胁迫下又有所下降,可能是高浓度盐胁迫引起叶片失水所致。张霞等[3]在研究盐胁迫对象草叶片解剖结构的影响时也得到类似的结论。一般而言,叶片栅栏组织越厚,细胞排列越紧实,栅栏组织/海绵组织比和结构紧密度越大,植物的耐盐性越强;海绵组织越厚,细胞排列越疏松,植物的耐盐性越差[6—7]。本研究表明,盐胁迫引起平欧杂种榛叶片栅栏组织/海绵组织比、结构紧密度显著增加或维持在对照水平,‘达维’和‘玉坠’叶片结构疏松度则表现出不同程度降低。结合前期研究与本研究可以得出,盐胁迫下,平欧杂种榛叶面积变小[21],上、下表皮增厚,叶片肉质化,栅栏组织细胞伸长,细胞层数增多,胞间隙变小,排列紧密,这些特征的变化是平欧杂种榛对盐环境的解剖结构响应,也是其对生境中大量盐离子造成的生理胁迫的一种适应性表现。一方面,叶小、厚实且紧密的叶肉细胞通过减少体内水分流失,降低蒸腾速率,从而提高水分利用效率[6];另一方面,栅栏组织不仅能高效运输水分,同时细胞内较多、较大的叶绿体能够增强光合作用[4]。
从不同品种来看,轻度和中度盐胁迫引起3个品种平欧杂种榛叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度显著增加,但重度盐胁迫下,‘达维’叶片厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度以及‘玉坠’叶片厚度、下表皮厚度和海绵组织厚度均降低至对照水平,而‘辽榛7号’叶片厚度、上表皮厚度、栅栏组织厚度和海绵组织厚度仍显著高于对照。同时,重度盐胁迫下‘辽榛7号’叶片结构紧密度较对照显著提高6.5%,明显高于‘达维’的2.0%和‘玉坠’的2.8%。上述结果表明,‘辽榛7号’叶片在盐胁迫下特别是重度盐胁迫下仍能通过一些解剖结构特征的调整表现出更强的形态结构适应性与耐盐性。
3.2 离子代谢特征对盐胁迫的响应与适应
NaCl胁迫下,植物体大量吸收并积累Na+和Cl-,并因此引起离子毒害或渗透胁迫,最终导致植物生长严重受阻[25]。面对NaCl胁迫,植物往往通过根系的离子选择性吸收来拒Na+或排Na+,或通过Na+区隔化来维持细胞内离子平衡,以此提高对盐胁迫的适应性[2,26—27]。尽管如此,不同植物对盐离子的区隔化方式存在差异,如唐古特白刺(Nitrariatangutorum)[28]、西伯利亚白刺(Nitrariasibirica)[29]、河滩芦苇(Phragmitesaustralis)[30]将Na+和Cl-主要集中在叶中,红砂(Reaumuriasoongorica)[31]的Cl-主要在茎部积累,而槲树(Quercusdentata)将Na+主要集中在根系[32]。也有研究表明,耐盐型和盐敏感型植物的离子区隔化方式不同,前者将Na+主要集中在根部,后者则将Na+主要集中在叶中[33]。本研究中,不同品种平欧杂种榛根、茎、叶中Na+和Cl-含量均随着盐胁迫程度的增强而显著增加,与对照相比,不同程度盐胁迫下以根中Na+和Cl-含量增幅最大,叶中增幅最小,表明盐胁迫下Na+和Cl-主要在根中积累,其次为茎,叶中积累最少。但从Na+和Cl-的绝对含量来看,叶中Na+和Cl-分布最多,根中分布最少,表明根系作为植物体最早感受盐胁迫信号的器官和最直接的受害部位,盐胁迫下首先会吸收并截留一定数量的Na+和Cl-,然后将其运输至茎和叶中,这与杨秀艳等[28]对唐古特白刺的研究结果一致。相关研究认为,这种适应机制一方面降低了盐离子对根系的毒害作用,保证植株对其它营养元素的吸收;另一方面,植株将盐离子更多地分布于叶中,可以利用盐离子作为无机渗透调节物质来增大地上部分与地下部分的渗透势差,促进水分吸收及其向上运输进而稀释液泡的高盐环境[29,34]。
作为植物生长发育所必需的营养元素,K+在离子平衡调控、细胞膨压和渗透调节等生理方面发挥着重要作用;而Ca2+作为植物信号传导中重要的第二信使参与植物的抗逆信号传导,调节着植物对逆境变化的响应过程[35],K+和Ca2+能调节植物的耐盐性[36—37]。本研究中,轻度和中度盐胁迫下不同品种平欧杂种榛根、茎对K+和Ca2+的吸收保持稳定或减少,而叶对K+和Ca2+的吸收有所增加,表明在一定盐胁迫范围内尽管平欧杂种榛根、茎的K+和Ca2+吸收下降,但叶具有一定的K+和Ca2+保有能力,进而维持体内离子的相对稳定来提高耐盐性。这与乌凤章等[16]对2个蓝莓品种的研究结果一致。一般来说,盐胁迫会抑制植物对营养离子(K+、Ca2+、Mg2+等)的吸收[17,32]。然而本研究发现,随着盐胁迫程度的持续增强,重度胁迫下不同品种平欧杂种榛根、茎对K+和Ca2+的吸收较中度甚至轻度胁迫时增加。与本研究结果相似,杨秀艳等[28]在研究唐古特白刺以及唐晓倩等[34]在研究西伯利亚白刺幼苗的盐胁迫响应时发现,300 mmol/L NaCl胁迫下两种白刺的根对K+的吸收以及唐古特白刺茎对Ca2+的吸收较200 mmol/L NaCl和100 mmol/L NaCl胁迫时增强,特别是两种白刺根中K+含量显著增加。引起这一现象的原因可能在于Na+和K+的水合半径相似,白刺可能通过增强根对K+的吸收以及茎对Ca2+的吸收来抑制根系过多的吸收Na+,从而在一定程度上减缓盐离子造成的危害。此外,研究发现盐胁迫处理四翅滨藜(Atriplexcanescens)7 d后,300 mmol/L胁迫下叶片中的Ca2+含量较对照和150 mmol/L胁迫时显著提高[38]。由此可见,植物在响应盐胁迫时对离子的吸收规律可能因植物材料、胁迫强度和胁迫阶段的不同而存在差异。
Na+是造成植物盐害的主要离子,K+是植物生长发育所必需的大量元素和渗透调节物质的重要组分。然而,由于2种离子具有相似的离子半径和水合能,盐胁迫下Na+对K+的吸收位点和活性位点呈现出明显的竞争性抑制作用,根细胞中的K+被Na+所取代,导致K+吸收减少,依赖K+的酶活性及代谢过程受到抑制[39]。因此,盐渍化土壤上的植物往往受到Na+毒害和K+亏缺的双重伤害,维持细胞中高水平的K+及K+/Na+比可以减少盐离子造成的毒害,是机体进行正常生理代谢所必须的[40]。此外,盐胁迫下茎、花等组织中维持适宜的Ca2+/Na+比已被证实是植物耐盐的关键[41]。本研究中,盐胁迫导致平欧杂种榛各器官K+/Na+比和Ca2+/Na+比显著降低,与盐胁迫下蓝莓[16]和大麦(Hordeumvulgare)[42]的离子代谢特征相一致。盐胁迫下,植物对Na+的大量吸收通过直接干扰和抑制细胞质膜对营养元素(K+和Ca2+)的吸收和转运,造成离子失衡,各器官K+/Na+比和Ca2+/Na+比下降[38],并由此进一步引起植株生长发育受阻。前期我们在研究盐胁迫对3个品种平欧杂种榛生长性状的影响时也得出,盐胁迫显著抑制幼苗的生长量和生物量[21]。
3.3 不同品种平欧杂种榛耐盐性差异
盐胁迫下,植物维持体内相对较高的K+/Na+比和Ca2+/Na+比是植物耐盐性强的一种表现。因此,植物能否在盐渍环境条件下生存在很大程度上取决于其体内维持K+/Na+和Ca2+/Na+平衡的能力[11]。本研究中,不同品种平欧杂种榛各器官K+/Na+比和Ca2+/Na+比在品种间均存在显著差异,具体表现为‘辽榛7号’>‘玉坠’>‘达维’,而K+/Na+比和Ca2+/Na+比的降幅表现为‘达维’>‘玉坠’>‘辽榛7号’,表明‘辽榛7号’对盐胁迫的适应性最强,其次为‘玉坠’,‘达维’最弱。张丽[20]采用主成分分析对盐胁迫下19个榛种质资源的14项生长和生理指标进行综合评价时也得出,本研究中的3个品种耐盐性由强到弱依次为‘辽榛7号’>‘玉坠’>‘达维’。
离子选择性运输系数表征植物对离子向上运输的选择能力,盐胁迫下营养离子的选择性运输系数越大,表明植株促进营养离子向上运输的能力同时抑制盐离子向上运输的能力越强,留在根中的盐离子越多,其耐盐性也越强[43]。本研究中,随着盐胁迫程度的增强,不同品种平欧杂种榛从根到叶以及从茎到叶的SK+,Na+、SCa2+,Na+均表现为先升高后降低的趋势,且在重度胁迫下仍显著高于或维持在对照水平,表明尽管盐胁迫抑制了平欧杂种榛对营养离子K+、Ca2+的吸收,但为了缓解因离子吸收受阻造成的不利影响,植株通过提高从根到叶以及从茎到叶的K+、Ca2+选择性运输以维持较高的生理代谢水平,进而减小盐胁迫对植株生长发育的影响。从不同品种来看,平欧杂种榛各器官间的离子选择性运输能力存在显著差异,其中,SK+,Na+在不同程度盐胁迫下的均值表现为‘辽榛7号’>‘达维’>‘玉坠’,SCa2+,Na+则表现为‘辽榛7号’>‘玉坠’>‘达维’,表明‘辽榛7号’通过加强对K+和Ca2+的选择性运输来维持体内离子平衡的能力强于其它2个品种。
4 结论
平欧杂种榛通过增加上、下表皮厚度,延伸栅栏组织长度,增加细胞层数且排列紧密,减小细胞间隙,形成肉质化叶片等一系列形态结构特征的调整来适应盐渍化生境。盐胁迫下,平欧杂种榛根系首先会吸收并截留一定数量的Na+和Cl-,然后将其运输至茎和叶中;‘辽榛7号’在积累盐离子的同时,通过加强对K+和Ca2+的选择性吸收和运输能力来维持体内的离子平衡以及较高的K+/Na+和Ca2+/Na+比,进而维持盐胁迫下机体正常的生理代谢活动,耐盐性强于‘达维’和‘玉坠’。尽管本研究从静态离子代谢的角度揭示了平欧杂种榛的盐胁迫响应与适应机制及不同品种的耐盐性差异,然而,离子平衡调控的动态转运机制和分子机制仍未明确,未来将利用非损伤微测技术(NMT)和转录组测序技术(RNA-seq)进一步深入探究根细胞离子流(Na+、H+、K+、Ca2+)的动态转运过程以及耐盐基因的调控机制。
