大豆胞囊线虫相关G2-like转录因子生物信息学分析
2022-03-30韩英鹏徐爱佟白芮瑶卜凡珊姜海鹏
韩英鹏,李 一,吴 桐,徐爱佟,白芮瑶,卜凡珊,姜海鹏
(东北农业大学大豆研究所,哈尔滨 150030)
大豆胞囊线虫(Soybean cyst nematode,SCN,Heterodera glycines Ichinohe)是世界范围内影响大豆产量的重要病害,是限制大豆高产和稳产重要因素[1]。SCN主要寄生于大豆植株根部,发病时地上部位叶片逐渐黄化、发育缓慢、植株矮小,严重时导致产量减少甚至绝收[2-3]。我国大豆胞囊线虫病主要分布于东北和黄淮两个大豆主产区。根据其致病力差异分为16个生理小种,其中东北地区主要为3号生理小种,而黄淮地区主要为4号生理小种[4]。
Golden2 like转录因子,又名G2-like和GLK,属于GARP转录因子超家族,首先在玉米中被发现,后续在苔藓(Physcomitrella patens)、拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)和番茄(Lycopersicon esculentum)等植物中陆续发现G2-like转录因子[5-6]。G2-like转录因子与光合作用密切相关,并参与叶绿体发育。然而在抗病方面研究较少,目前主要集中在模式植物拟南芥中。Savitch等在拟南芥中过表达G2-like基因(AtGLKl),发现转基因植株提高拟南芥对禾谷镰刀菌抗性[7]。Han等对G2-Like基因(glkl,glk2)作双突变,发现突变后拟南芥植株对黄瓜花叶病毒表现高度敏感性[8]。Nakamura等发现G2-Like基因(OsGLK1)在水杨酸合成途径中表达量明显上调,参与水稻相关病害防御[9]。
利用转录组测序技术可揭示某些特定植物生物学过程及抗逆机制。近年转录组测序技术广泛应用于大豆胞囊线虫病抗性基因筛选[10]。2005年,Klink等利用Williams82,PI88788,Peking(PI548402)接种SCN后转录组数据发现并鉴定SCN抗性基因[11]。2006年,Alkharouf等分析SCN侵染不同阶段转录组数据,发现SCN侵染时段不同基因表达也不同[12]。2017年,Zhang等利用大豆胞囊线虫侵染野生大豆幼苗根系后转录组测序数据探究大豆与SCN互作机制[13]。越来越多学者利用转录组测序技术挖掘与大豆胞囊线虫病抗性相关基因。然而,关于G2-like家族在SCN胁迫下行使功能和作用机制却鲜有研究。
本研究依据SCN 3胁迫大豆转录组数据,对筛选得到的G2-like家族差异表达基因作生物信息学分析,以期为进一步研究SCN抗性相关G2-like转录因子功能和提高大豆SCN抗性提供理论支持,也为抗SCN育种提供基因资源。
1 材料与方法
1.1 材料
大豆材料“东农L-10”和“Lee”取自东北农业大学大豆生物学教育部重点实验室。选用大豆材料经Riggs和Schmitt鉴别模式鉴定为大豆胞囊线虫病3号生理小种[14],均于东北农业大学大豆研究所试验田采集。
1.2 方法
1.2.1 材料及处理
首先种植感病品种“Lee”于病土中繁殖胞囊,30 d后取出完整根系,用自来水快速冲洗,用40+60目筛收集胞囊。用橡胶塞轻轻研磨胞囊,显微镜下定容卵溶液至2 000个·mL-1。然后将“东农L-10”种植于蛭石中,设置对照,3次重复,每次10株,昼夜温度约为27~28℃,相对湿度为60%~70%。待长成幼苗后,接种胞囊,每株植株接种卵液1 mL。以接种SCN 3号小种为试验组,未接种为对照组。分别于接种0和10 d后采集根部样品,并对其作酸性品红染色,以确认接种成功。
1.2.2 转录组测序
将上述所取根样迅速置于液氮中,提取总RNA,利用TRNzol Universal试剂盒,保存于-80℃冰箱备用。构建cDNA文库,利用Illumina TruSeq RNA文库制备试剂盒,最后利用Illumina HiSeqTM2500 cDNA作转录组测序。分析测序结果,筛选出大豆胞囊线虫病3号生理小种胁迫下东农L-10中差异表达的G2-like转录因子。
1.2.3 qRT-PCR检测
应用Primer Premier 5.0设计G2-like基因荧光定量引物(见表1),内参基因选用GmActin4(Gen-Bank No:AF049106)。使 用Applied Biosystems 7500荧光定量PCR仪,软件系统为7500 System Software Version 2.3。依照TaKaRa公司荧光定量TB Green试剂盒说明书操作,共40个循环,首先在95℃下预变性30s,于95℃变性3s,接下来60℃退火30s,最后72℃延伸30s。相对基因表达分析采用2-ΔΔCt法,其中CT值由3次重复试验取平均值计算。

表1 差异表达基因荧光定量引物Table 1 Quantitative primersfor the differentially expressed genes
1.2.4 蛋白质理化性质分析
通过在线软件Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)下载大豆胞囊线虫病3号生理小种相关转录因子信息,通过在线软件PrtoParam(https://web.expasy.org/protparam/)分析G2-like转录因子成员氨基酸残基数、等电点、不稳定系数等理化性质。
1.2.5 亚细胞定位
通过在线软件Cell-PLoc2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)分析G2-like转录因子亚细胞定位。
1.2.6 蛋白质结构预测
利用在线软件SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测分析G2-like转录因子蛋白质二级结构,利用SWISS-MODE(https://www.swissmodel.expasy.org)预测分析其三级结构。
1.2.7 蛋白质保守基序分析
通过在线软件MEME(http://meme-suite.org/tools/meme)分析G2-like转录因子蛋白家族保守基序,motifs数值设为10,其余参数默认,利用TBtools软件作可视化。
1.2.8 基因结构分析
通过在线软件Gene Structure Display Server 2.0(http://gsds.gao-lab.org/index.php)预测G2-like基因结构。
1.2.9 构建系谱进化树
通过在线软件NCBI(https://www.ncbi.nlm.nih.gov)数据库筛选选得到拟南芥、水稻、玉米等不同物种6个G2-like转录因子家族成员,使用Clustal X对上述蛋白多重序列比对,参数默认。采用邻接法(Neighbour-Joining,NJ),通过软件MEGA7构建系谱进化树,Bootstrap replications值设定为1 000。
1.2.10 基因表达模式分析
利 用Phytozome(https://phytozome-next.jgi.doe.gov)数据库下载大豆根茎叶5个组织表达量数据,然后利用TBtools软件绘制表达谱热图。
1.2.11 启动子顺式作用元件分析
G2-like基因上游2 000 bp DNA序列利用phytozome数据库获取,通过在线软件PlantCare(http://bioinformatice.psb.ugent.be/webtools/plantcare/html)预测启动子上游顺式作用元件,结果通过Excel 2016软件整理分析。
2 结果与分析
2.1 G2-like基因表达分析
本研究通过RNA-seq方法分析SCN 3胁迫和未胁迫下抗病品种“东农L-10”中差异表达基因,发现与其他基因家族相比,G2-like家族成员表达量差异倍数相差较大,利用qRT-PCR方法比较分析筛选出G2-like家族8个成员在SCN 3胁迫0和10 d后根部基因表达量,发现这8个G2-like基因相对表达量在10 d时显著上调,与RNA-seq数据相比,表达趋势相同,Chen等发现G2-like基因家族与病害防御相关基因表达有关[6],表明这8个G2-like基因可能在SCN 3胁迫过程中发挥重要作用。
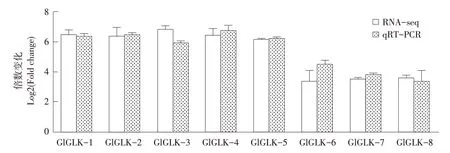
图1 SCN3胁迫下G2-like基因相对表达量变化Fig.1 Change of relative expression of G2-like gene under SCN3 stress
2.2 G2-like转录因子蛋白理化性质分析
通过在线软件PrtoParam分析大豆胞囊线虫相关G2-like转录因子理化性质,结果表明,8个上调表达基因氨基酸残基数范围为237(Gl-GLK-8)~427(GIGLK-3),分子质量范围为26.80 ku(GIGLK-8)~47.70 ku(GlGLK-3);等电点范围为5.51(GlGLK-4)~9.45(GlGLK-8),其中GlGLK-4、GlGLK-5和GlGLK-7等电点均小于7,为酸性蛋白,其余等电点均大于7,为碱性蛋白;脂肪系数为62.61(GlGLK-2)~71.97(GlGLK-2),表明G2-like转录因子家族蛋白成员间热稳定差异较小(见表2)。
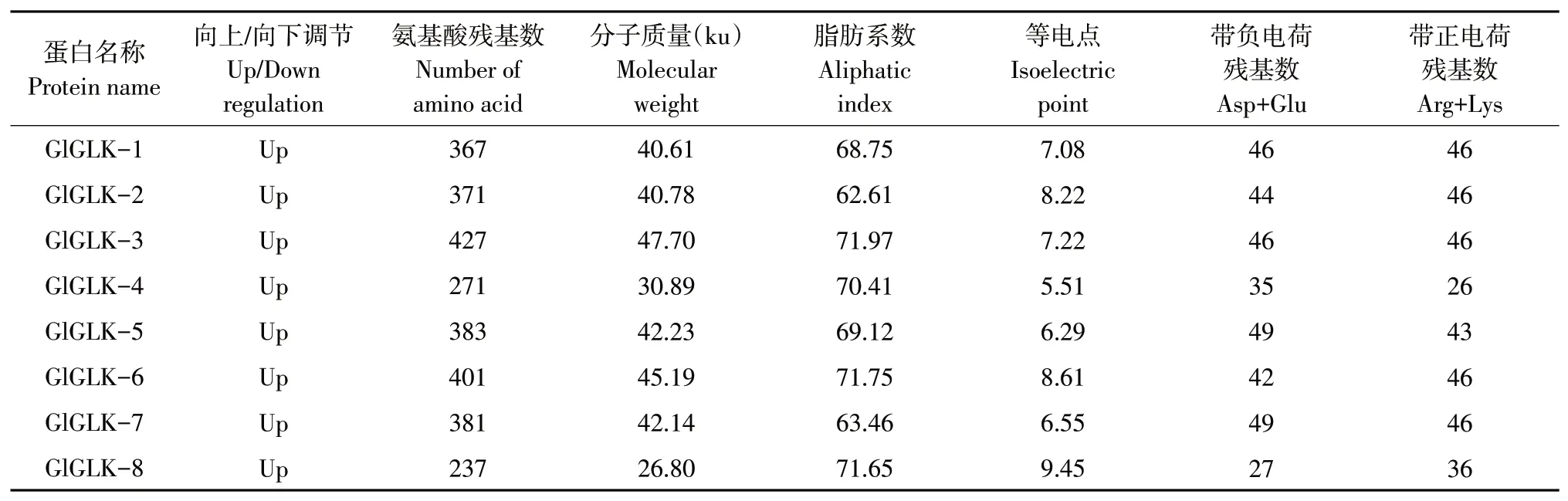
表2 G2-like转录因子家族蛋白理化性质Table 2 Protein member information of soybean G2-like transcription factor
通过Protscole预测G2-like转录因子氨基酸亲疏水性,结果表明,8个G2-like转录因子总平均亲水性值均为负数,均为亲水蛋白。其中总平均亲水性最大为GlGLK-3,为-0.603,该成员第383位氨基酸表现为最大亲水性,为-2.933;第187位氨基酸表现为最大疏水性,为1.589。总平均亲水性最小为GlGLK-2,为-0.788,该成员第102位氨基酸表现为最大亲水性,为-3.178;第288位表现为最大疏水性,为2.467(见表3)。
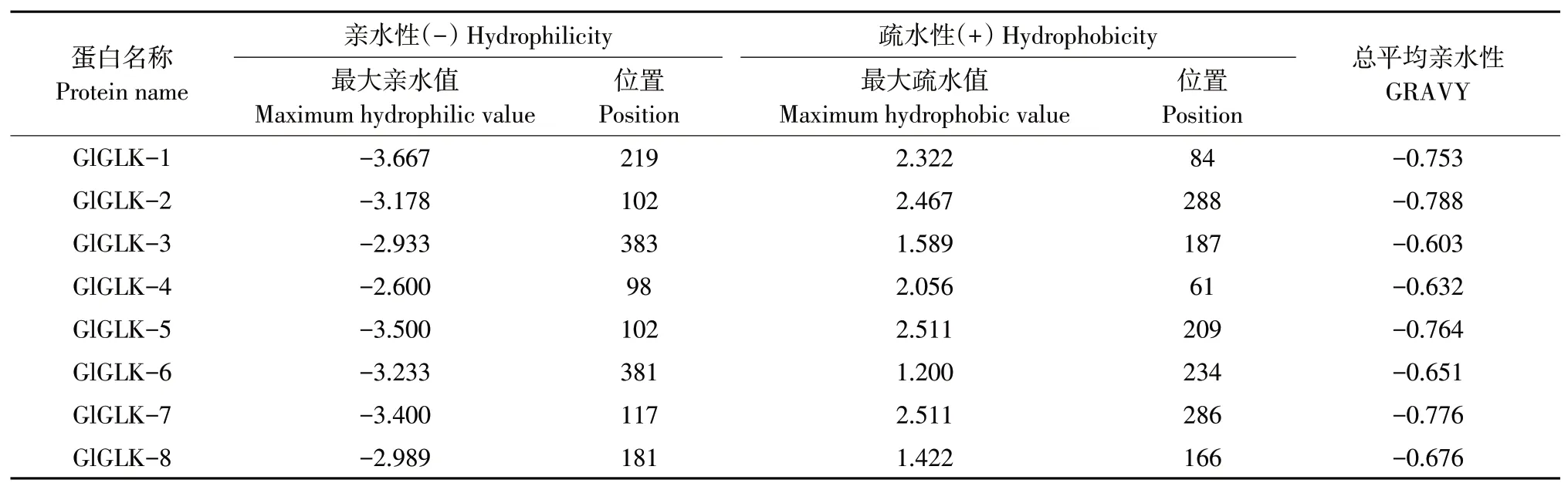
表3 氨基酸亲疏水性Table 3 Amino acid hydrophilicity and hydrophobicity
2.3 蛋白质结构预测
通过在线软件SOPMA分析大豆胞囊线虫G2-like转录因子蛋白质二级结构,结果表明8个G2-like转录因子成员除GlGLK-8外,其他蛋白质二级结构与拟南芥相似,均以无规则卷曲为主,其次为α-螺旋和延伸链,β-转角含量最少;GlGLK-8二级结构以α-螺旋为主,其次为无规则卷曲和延伸链,β-转角含量最少(见表4)。
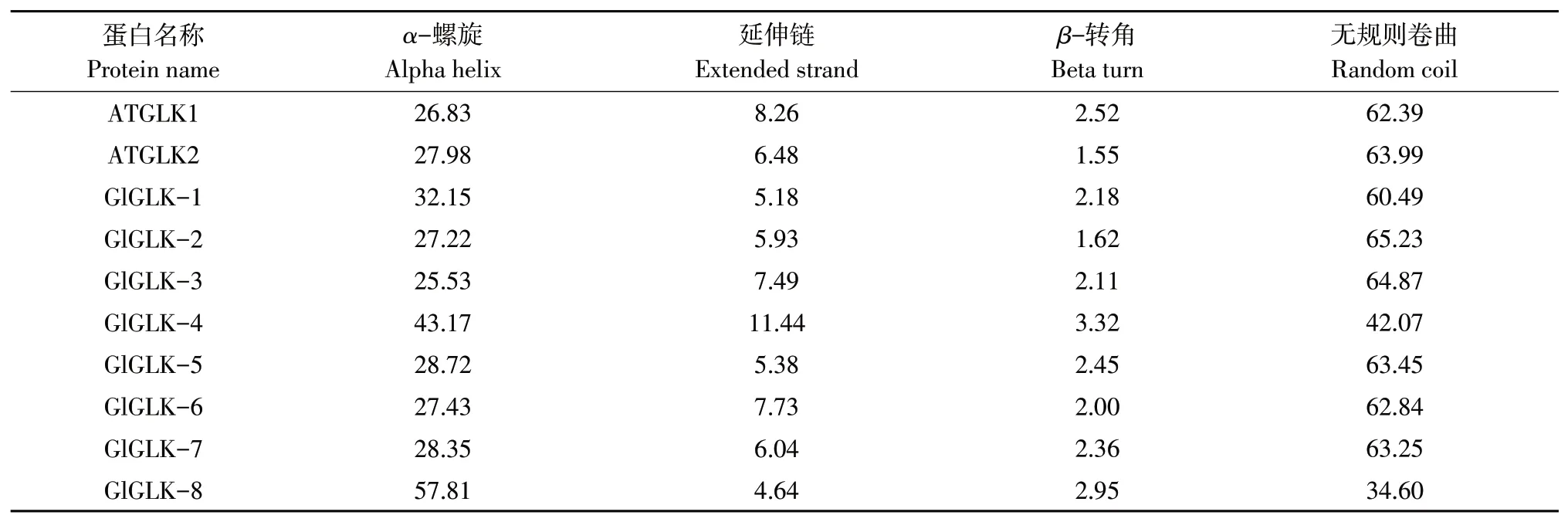
表4 二级结构预测Table4 Secondary structureprediction (%)
通过在线软件SWISS-MODE对大豆胞囊线虫G2-like转录因子蛋白质三级结构与拟南芥作比对(见图2),ATGLK2蛋白与GlGLK-5、GlGLK-7蛋白三级结构近乎相同,推测其行使相同或相似功能。
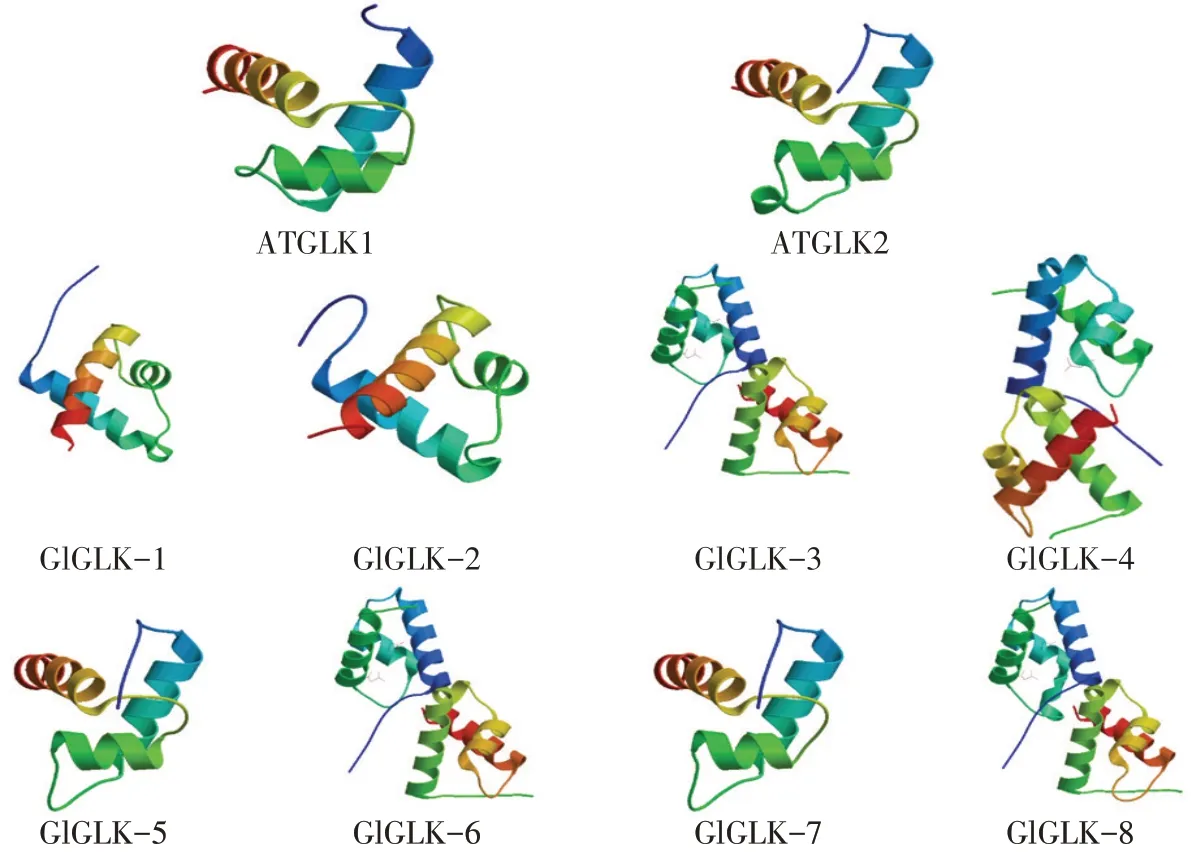
图2 G2-like转录因子家族蛋白质三级结构预测Fig.2 Tertiary structure prediction of G2-like transcription factors
2.4 亚细胞定位
对8个大豆G2-like转录因子基因编码氨基酸序列作亚细胞定位,可了解基因在植物生长发育过程中遗传机制。通过预测G2-like蛋白质亚细胞定位(见表5),8个大豆G2-like转录因子成员与ATGLK2相同,均定位于细胞核,说明植物在受到大豆胞囊线虫病胁迫时,G2-like转录因子家族发挥重要作用的位置可能为细胞核。
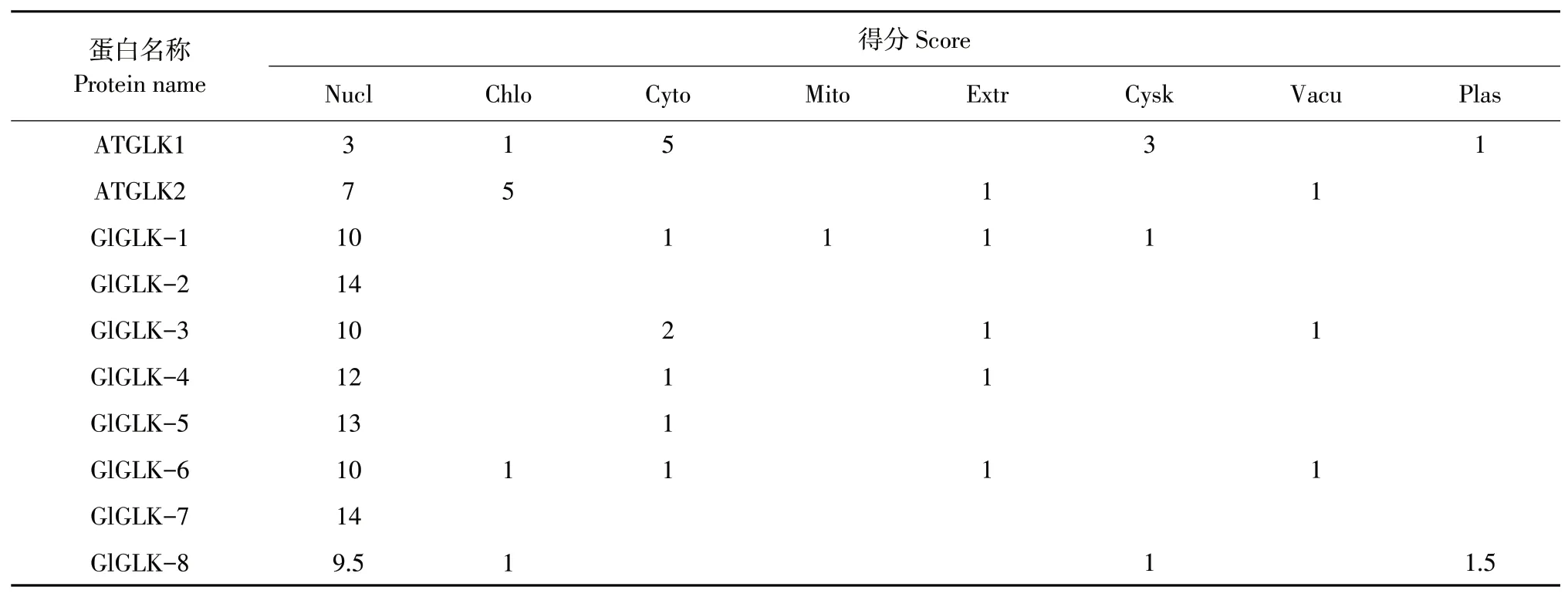
表5 G2-like基因家族亚细胞定位预测Table5 Prediction of subcellular location of G2-likegenefamily
2.5 蛋白质保守基序分析、基因结构分析及构建系谱进化树
利用在线软件MEME分析G2-like转录因子保守基序,结果表明,G2-like转录因子成员共有10种类型蛋白保守基序(见图3),且不同转录因子具有不同数目及不同类型保守Motif结构,但均含有Motif 1和Motif 2结构,不同蛋白序列含有不同基序类型和数量,蛋白功能构成的基本单位是基序,因此其功能存在一定分化。
基因结构多样性可作为基因家族分类重要参考依据。为进一步探究基因功能和进化之间关系,利用G2-like基因成员CDS序列和DNA序列分析其基因结构。结果表明(见图3),大豆GlGLK-8基因长度最长,GlGLK-1基因长度最短;大豆G2-like基因外显子内含子数量差异较大,外显子数目均为两个,内含子数目不同,但差异较小,最多为6个(GlGLK-4),最少为4个(GlGLK-1、GlGLK-2、GlGLK-5和GlGLK-7)。

图3 G2-like基因家族分析Fig.3 Analysis of G2-like gene family
为进一步研究G2-like基因进化关系,采用NJ法将筛选得到8个大豆G2-like基因和6个其他物种G2-like基因,利用全长蛋白质序列构建系统发育进化树(见图3)。从图中可看出,GlGLK-1、GlGLK-2、GlGLK-5和GlGLK-7与拟南芥ATGLK1和ATGLK2相距较近,说明其同源性高,GlGLK-3、GlGLK-4、GlGLK-6和GlGLK-8处于相距较远的分支,其功能有待进一步研究。
2.6 不同部位G2-like基因表达量分析
为进一步了解在大豆生长发育过程中G2-like基因家族发挥的功能与作用,本研究于phytozome在线网站下载大豆根、茎、叶中G2-like基因表达量数据,并绘制基因表达热图(见图4)。结果表明,G2-like基因在根、茎、叶部位均有不同程度表达,根中表达量最高,因SCN主要通过侵染大豆根部,影响相关基因表达[3],因此推测G2-like基因与SCN抗性相关。其中GlGLK-2在根中表达量最高,GlGLK-6表达量最低,说明GlGLK-2在SCN侵染过程中可能发挥更重要作用,GlGLK-2和GlGLK-5在叶中表达量显著高于茎中表达量,其余GlGLK-2基因在茎与叶中表达量无显著差异。
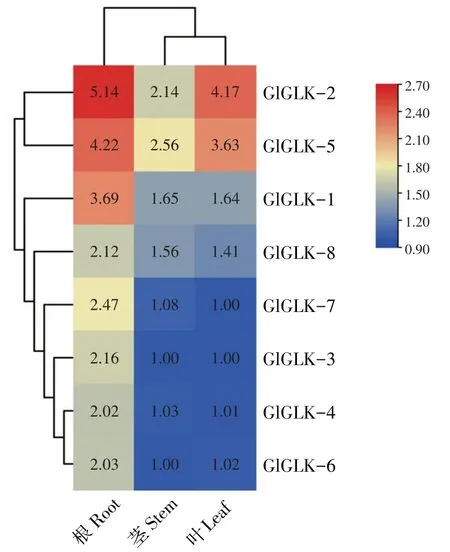
图4 大豆G2-like基因在不同组织下表达Fig.4 Expression profilesof soybean G2-likegenesin different tissue conditions
2.7 启动子元件分析
在基因调控中,启动子顺式作用元件发挥重要作用。通过在线软件PlantCare分析G2-like基因顺式作用元件(见图5),结果表明,大豆G2-like家族基因上游存在4种类型启动子顺式作用元件:ABRE、TCA-element、P-box激素响应相关元件等;AE-box、MRE、Box4光合反应相关元件等;生长发育相关元件ARE;TGACG-motif、LTR、O2-size等逆境胁迫相关元件。其中GlGLK-3所含启动子元件数目最多,包括光响应元件Box4和CTAAT-box、呼吸元件ARE和逆境相关元件MBS,表明GlGLK-3基因在大豆生长发育及逆境胁迫中发挥重要作用。
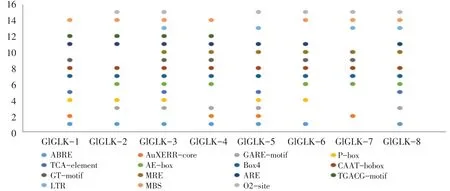
图5 大豆G2-like基因上游启动子区顺式作用元件分析Fig.5 Analysisof cis-acting elements in the promoter region of soybean G2-like gene
3 讨论
G2-like转录因子广泛存在于植物中,其主要功能表现在叶绿体发育、果实品质、生物胁迫和非生物胁迫、植物衰老和激素影响等方面[14]。目前G2-like转录因子在抗病方面研究主要集中于拟南芥,大豆中相关功能研究尚无明确报道。本研究通过转录组测序技术在大豆品种东农L-10(抗病)中筛选出与大豆胞囊线虫3号生理小种相关的8个G2-like转录因子,分析其生物信息学,初步判断G2-like转录因子在大豆胞囊线虫3号生理小种胁迫下应答机制。
亲缘关系远近与转录因子行使功能密切相关,Matus等通过构建系谱进化树,将位于相邻位置上的转录因子认定为具有相同或相似功能[16]。拟南芥G2-like家族基因AtGLK1和AtGLK2在功能上等效,与植株抗病性密切相关[17]。GlGLK-1、GlGLK-2、GlGLK-5和GlGLK-7与拟南芥ATGLK1和ATGLK2相距较近,说明其同源性高。G2-like转录因子蛋白质基序具有高度保守性,均含有Motif 1和Motif 2结构;GlGLK-5、GlGLK-7与ATGLK2蛋白值三级结构近乎相同,可进一步推测GlGLK-5、GlGLK-7蛋白行使功能与ATGLK2相似,参与大豆植株逆境胁迫过程,其功能有待进一步研究。
不同植株中G2-like基因表达情况也不同,且不同基因在不同组织器官中不同表达显示其所发挥作用有差异[18-19]。本研究主要统计大豆8个G2-like转录因子基因在根、茎、叶不同组织中特异表达情况,结果发现G2-like转录因子基因在大豆根部表达量最高,同时G2-like转录因子对于植物根部研究已成为新研究热点。刘俊芳等发现番茄G2-like基因(SlGlk1、SlGlk7和SlGlk15等)在根部具有较高表达量[20]。刘芳等发现玉米G2-like基因(ZmGlk9、ZmGlk35和ZmGlk48等)在根中表达量也相对较高[21]。拟南芥中G2-like基因过表达不仅影响拟南芥根部叶绿体发育,可增加根部CO2固定且在一定程度上提高植物对碳的利用率,对植物生长起重要作用[18]。GlGLK-2和GlGLK-5在根部表达量相对较高,推测其通过影响根系发生,调节植株生长情况,参与植株光合作用及抗逆机制。
启动子是调控基因表达重要元件,因启动子上游顺式作用元件是转录因子结合位点和决定转录激活和抑制关键因子[22]。Ahmad等发现拟南芥glkl和glk2双突变体在植发育过程中表现出ABA超敏表型,且调控ABA应答基因wrky 40[23]。G2-like转录因子可能参与调控拟南芥光合器官发育。Han等在玉米中发现茉莉酸(JA)参与G2-like转录因子介导病毒抗性,同时也发现G2-like基因在低温、干早胁迫方面发挥重要作用[8]。通过分析大豆G2-like基因启动子元件发现,启动子序列中含有光响应元件AE-box、GT-motif、MRE;参与脱落酸反应的元件ABRE及保守DNA模块的参与光响应一部分Box 4;还包括对厌氧诱导至关重要顺式作用元件ABRE、参与植物防御和胁迫反应的MBS、响应JA顺式作用元件TGACG-motif。初步表明大豆G2-like基因在大豆生长发育及逆境胁迫中发挥重要作用。
综上所述,亲缘关系相近蛋白质,结构相似,可能发挥相似生物学功能。本研究结果得到大豆胞囊线虫3号生理小种胁迫下筛选出的G2-like转录因子成员及其序列特征以及相应表达信息,为进一步全面解析G2-like转录因子各成员生物学功能及作用机制提供参考,也为探究大豆胞囊线虫病抗病机理提供理论依据。
4 结论
利用东农L-10(抗病)在SCN 3胁迫和未胁迫条件下转录组数据,通过对比数据筛选出8个与SCN抗性相关G2-like转录因子。生物信息学分析结果表明8个G2-like转录因子蛋白质氨基酸数目为237~436;相对分子质量为26.80~47.70 ku,均定位于细胞核,表现为亲水性蛋白;蛋白质二级结构除GlGLK-8以α-螺旋为主外,其余均以无规则卷曲为主;G2-like基因结构均含有不同数目外显子和内含子;8个G2-like转录因子成员共有10种类型蛋白保守基序,均含有Motif 1和Motif 2结构;G2-like基因在根、茎、叶部位均有不同程度表达,根中表达量最高,GlGLK-2和Gl-GLK-5在叶中表达量显著高于茎中表达量,其余G2-like基因在茎与叶中表达量无显著差异;Gl-GLK-3含有Box4和CTAAT-box(光响应元件)、ARE(呼吸元件)和MBS(逆境相关元件)等与大豆生长发育及逆境胁迫密切相关顺式作用元件。系统进化树结果分析表明,GlGLK-1、GlGLK-2、GlGLK-5和GlGLK-7与拟南芥ATGLK1和ATGLK2相距较近,预示功能相似性,而GlGLK-3、GlGLK-4、GlGLK-6和GlGLK-8处于相距较远分支,功能有待进一步研究。
