磴口农田防护林新疆杨不同时空叶生物量比较
2022-03-25赵英铭张家祺黄雅茹马迎宾唐进年郝玉光刘明虎
赵英铭,褚 洋,张家祺,黄雅茹,马迎宾,唐进年,郝玉光,刘明虎†
(1.中国林业科学研究院沙漠林业实验中心/国家林草局内蒙古磴口沙漠实验研究站/乌兰布和沙漠综合治理国家长期科研基地,015200,内蒙古磴口;2.中国林业科学研究院荒漠化研究所,100091,北京;3.中国林业科学研究院华北林业实验中心,102300,北京;4.内蒙古工业大学航空学院,010051,呼和浩特;5.甘肃省治沙研究所,730070,兰州)
全球气候变暖引发一系列问题[1],而植物叶片是吸收CO2合成葡萄糖等物质的主要场所[2],叶片是进行光合作用将光能转化为化学能的主要器官[3],也是生态系统中固碳减缓全球气候变化最重要的器官。所以叶生物量的大小,对于整个生态系统而言都有着不可估量的作用。
我国北方农田防护林以杨树为主要造林树种,形成林网规格不一的落叶阔叶林。每年10—11月叶都会周期性落尽,故树木各生长器官生物量、碳储量作比较时,叶生物量、碳储量只能统计年均生物量、碳储量,但干、皮、枝、根都是连年生物量,时间尺度不一样,比较结果叶生物量、碳储量必然最小[4],很难得出具有科学合理的比较结果。目前没有一个可行的研究方法计算连年叶生物量和碳储量,以进行同时间尺度各生长器官生物量、碳储量的比较。为比较不同时空年均和连年叶生物量差异性,将叶与其它器官放在同一时空尺度进行比较,客观评价新疆杨叶器官的固碳功能,以精准计算出连年累积叶碳储量,便可知叶片在固碳中的累积作用,这对于计算植物叶片固碳累积意义重大。
1 研究区概况
绿洲集中了90%以上的人口和财富[5],是西北干旱区经济发展的主要区域。以河套平原为例,是我国小麦、玉米的主产区,国家的中央粮库储备基地,对于我国人口众多的国情而言,其重要性不言而喻。然而,该区西侧分布着乌兰布和沙漠,风沙危害严重,春季常常出现沙打苗、沙埋和沙尘暴等灾害[6],致使农业减产[7-8]。农田防护林可起到有效的防风固沙作用,尤其叶片的滞尘作用可净化空气[9],对于改善环境质量有明显的作用[10]。叶片的滞尘作用又取决于叶表面积,表面积越大滞尘作用越大[11]。研究区位于内蒙古磴口绿洲乌兰布和综合治理长期科研基地——中国林业科学研究院沙漠林业实验中心第二实验林场,地理坐标为E 106°47′5.47″、N 40°27′28.19″,海拔1 051 m,原始地貌为沙丘及丘间低地,亦有白刺风积沙丘,沙壤土和黏土。试区属大陆性季风气候区,全年气候干旱少雨,年均降雨量144 mm,蒸发量3 000 mm左右,地下水位7~8 m,是沙尘暴发源地之一。试区常年风沙较大,尤以3—5月风沙活动最强烈。为此,建设大范围多树种多结构绿洲农田防护林网,有明显的防风固沙保护农田、改善环境质量[10]等效果,为试验提供了丰富的实验材料和研究基础。20世纪80年代考虑到造林成本,绿洲人工农田防护林多采用速生的新疆杨(Populusalbavar.pyramidalis)等为主要造林树种,结构为多行的宽林带或者片林。新疆杨具有良好的速生、耐盐、抗旱、抗寒、抗虫和干形良好等生态适应特性,使其迅速成为目前沙区主栽树种之一,目前仍是磴口乃至多地干旱区、半干旱区绿洲农田防护林的主要树种。由于宽林带或片林凸显耗水量大、边行效应显著、保存率低等问题,所以后期林带结构多选用株行距4 m×4 m,行数6~8行的,但仍存在耗水大和边行效应的问题。为消除边行效应并保证防护林充足的水源目前采用2行渠道林结构,株距1 m,行距2~7 m,该结构表现出良好的生长势,在5~7年便起到良好的防风效果[12]。
2 数据与方法
2.1 数据来源
研究数据来源于内蒙古磴口县中国林科院沙林中心二实验场水权置换渠道整改林带更新采伐,实地调查田渠林路式2行新疆杨防护林,株行距3 m×7 m,2行,林带年龄20年,林带结构参数调查106株,平均胸径27.75 cm,平均树高19.96 m,采伐调查生物量样本数20株,渠道宽4 m,渠道边至新疆杨1.6 m,新疆杨至柏油路1 m。林网规格300 m主林带×140 m副林带,主林带与副林带树种和株行距一致。龄级是按照幼龄林0~10 cm胸径,中龄林10~20 cm胸径,近熟林20~30 cm胸径,成熟林30~40 cm进行划分的,经过树干解析,幼龄林需要7年(包括苗圃地2年),中龄林4年,近熟林和成熟林各5年。叶生物量调查每株按照冠层分上、中、下3层,分别称量总鲜质量,然后每层取基径居中的样枝3个,分别称量样枝叶、样枝和样叶鲜质量(g),精确到1 g,得到样品枝叶鲜质量比。将每个样枝的叶采用全部取样法全部取样编号,如1(树号)-上(冠层)-1(样枝编号)-叶,并装入牛皮纸袋中并封口,待烘干。每个样枝均匀取样500 g左右并编号,如1(树号)-上(冠层)-1(样枝编号)-枝。再将枝、叶样品65 ℃烘干称至恒质量,得到样枝干鲜比,再由冠层总鲜质量与枝叶鲜质量比得到每个冠层叶鲜质量,再与叶干鲜比乘积得到叶干物质的量,然后累加3个冠层得到叶总干物质的量即叶生物量。
2.2 建模方法
2.2.1 单株年均叶生物量建模 采用空间代替时间的方法,用同一空间内不同径级胸径相对应的年均叶生物量代替不同林龄相应径级叶生物量,与林带结构参数胸径、树高、冠幅等因子建模,得到年均叶生物量模型。幼龄林胸径10 cm以下,中龄林胸径10~20 cm,近熟林胸径20~30 cm,成熟林30~40 cm。结合树干解析得到年龄和相应胸径,将逐年胸径代入年均叶生物量模型得到不同年龄相应的一年生叶生物量,并直接与年龄建模,得到年龄为变量因子的一年生叶生物量模型。
2.2.2 单株连年生叶生物量建模 通过树干解析,得到不同年龄对应的胸径,将树干解析胸径代入年均叶生物量模型得到每年年均叶生物量,然后逐年累加得到连年叶生物量,与胸径、树高、冠幅建立年均连年叶生物量模型。亦可通过树干解析将胸径和年轮建立增长模型,再由胸径和叶一年和连年生物量模型得到一年叶生物量和连年生物量数据,然后和年龄直接建模。
1)林带叶生物量建模方法。林带叶生物量应为单位林带长度内所拥有的叶生物量与林带长度的乘积,而林带叶生物量模型应为单株叶生物量模型与行数、保存率之积除以株距再乘以林带长度,模型如下:
(1)
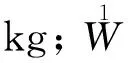
式(1)中林带树种、株距、年龄、行数均相同则可合并为:
(2)
式中:1W为林带叶生物量,kg;W为林带单株叶生物量,kg;k为林带行数;ρ为林带保存率,%;M为主林带长,m;O为副林带长,m;L为林带株距,m。
2)林网叶生物量建模方法。林网叶生物量应为林带叶生物量与林网面积之比,则模型为:
(3)
式中2W为林网叶生物量,kg。
式(3)中林带树种、株距、年龄、行数均相同则可合并为
(4)
3 结果与分析
3.1 单木年均叶生物量及模型
1) 新疆杨单木年均叶生物量。1~7年新疆杨处于幼龄期,包括苗圃地2年,第7年单木叶生物量为(1.112 8±0.628 6)kg,年增量约为0.159 0 kg/a。8~11年为树木中龄林阶段,叶生物量增长5.0倍,此阶段叶生物量增幅最大,进入生长旺季,年增量约为1.125 5 kg/a。12~16年新疆杨进入近熟林时,叶生物量增长1.7倍,年增量为0.830 7 kg/a。17~21年新疆杨进入成熟龄,叶生物量增长2.5倍,年增量约为3.027 8 kg/a,此时年增量最大。随着林龄增长年均叶生物量呈增长趋势,涨幅呈先增大后缩小趋势,中龄林出现极值;年均叶生物量年增量随林龄亦呈增长趋势,成熟林最大;幼龄林所需时间最长,中龄林所需时间最短,近熟林和成熟林所需时间居中且相同(图1)。

图1 不同树龄新疆杨单木叶生物量Fig.1 Leaf biomass of Populus alba var. pyramidalis individual plant with tree-age
2)单木年均叶生物量模型。由新疆杨单木不同径级年均叶生物量与胸径的关系(图2),建立新疆杨单木年均叶生物量模型,经过线性、对数、反向、二次、三次、复合、幂、S、增长、指数和logist等回归方程筛选,得出幂函数异速生长方程拟合度最高,为
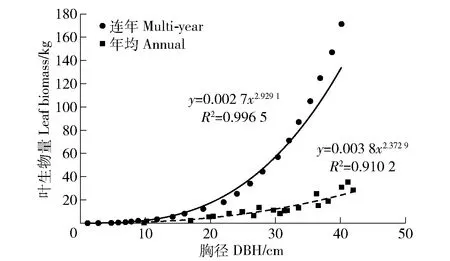
图2 不同径级新疆杨年均和连年叶生物量Fig.2 Annual and multi-year leaf biomass of Populus alba var. pyramidalis with DBH
(5)
式中:W1为叶生物量,kg;D1.3为胸径,cm。R2=0.910 2,经检验P<0.01,回归关系非常显著。
同理,由胸径、树高、冠幅因子和年均叶生物量建模得到绿洲防护林带单株年均叶生物量多元模型,经过线性等回归方程筛选,得出年均叶生物量模型(表1)。
3.2 单木连年叶生物量及模型
1)单木连年叶生物量。根据树干解析得出年轮与胸径,将胸径代入表1中年均叶生物量模型,得到年均叶生物量,累加计算得到21年相应连年叶生物量。幼龄林单木连年叶生物量为(2.677 4±0.664 1)kg,年增长量为0.382 5 kg/a;中龄林单木叶连年累积生物量增长5.7倍,所需时间仅为4年,叶生物量增速最快,耗时最短,年增量为3.112 8 kg/a。12~16年中龄林进入到近熟林这个过程需要5年,这5年单木叶连年累积生物量增长3.3倍,增速有所减缓,所需时间变长,近熟林单木年增量为7.093 1 kg/a。17~21年由近熟林进入成熟林这个过程需要5年,5年单木叶连年累积生物量增长3.1倍,增速明显减缓,所需时间较长,成熟林年增量为21.718 5 kg/a。说明连年叶生物量随林龄的增长呈增长趋势,涨幅呈减小趋势;连年叶生物量年增量随林龄的增长也呈增大趋势,而且增长的幅度较大(图1)。

表1 新疆杨年均叶生物量模型Tab.1 Growth models of annual leaf biomass of Populus alba var. pyramidalis
2)单木连年叶生物量模型。由胸径和连年叶生物量关系,建模得到胸径单因子的连年叶生物量模型,经筛选幂方程拟合度最高,R2=0.906 5,经检验回归关系非常显著。同理由林带结构因子胸径和树高、树高、南北冠幅、东西冠幅、冠幅乘积与连年叶生物量建模(表2)。

表2 新疆杨连年叶生物量增长模型Tab.2 Growth models of leaf biomass of Populus alba var. pyramidalis for multi-years
3.3 年均和连年叶生物量比较
3.3.1 单木年均和连年叶生物量比较 将连年叶生物量与年均叶生物量作比较如图3所示,幼龄林、中龄林、近熟林和成熟林连年叶生物量是年均叶生物量的比例分别为182.54%、323.10%、436.21%和515.79%,均>100%,且比例增量逐渐增加;中龄林、近熟林和成熟林连年叶生物量与年均叶生物量比例增速分别为1.77、1.35和1.18,逐渐减小(图3)。

图3 不同树龄单木连年叶生物量与年均之比Fig.3 Ratio of multi-year leaf biomass to annual for a different tree-age individual plant
3.3.2 林带年均和连年叶生物量比较 440 m(主林带300 m×副林带140 m)的新疆杨林带年均叶生物量幼龄林、中龄林、近熟林和成熟林分别为(0.312 4±0.176 4)、(1.524 5±0.049 7)、(2.562 5±0.364 2)和(6.331 4±1.535 2)t,而各林龄连年叶生物量分别是年均叶的2.4、2.7、5.2和6.4倍。表明随林龄增长林带年均和连年叶生物量亦呈增长趋势;各林龄林带连年叶生物量总是大于年均,随着林龄的增长差距越来越大(图4)。
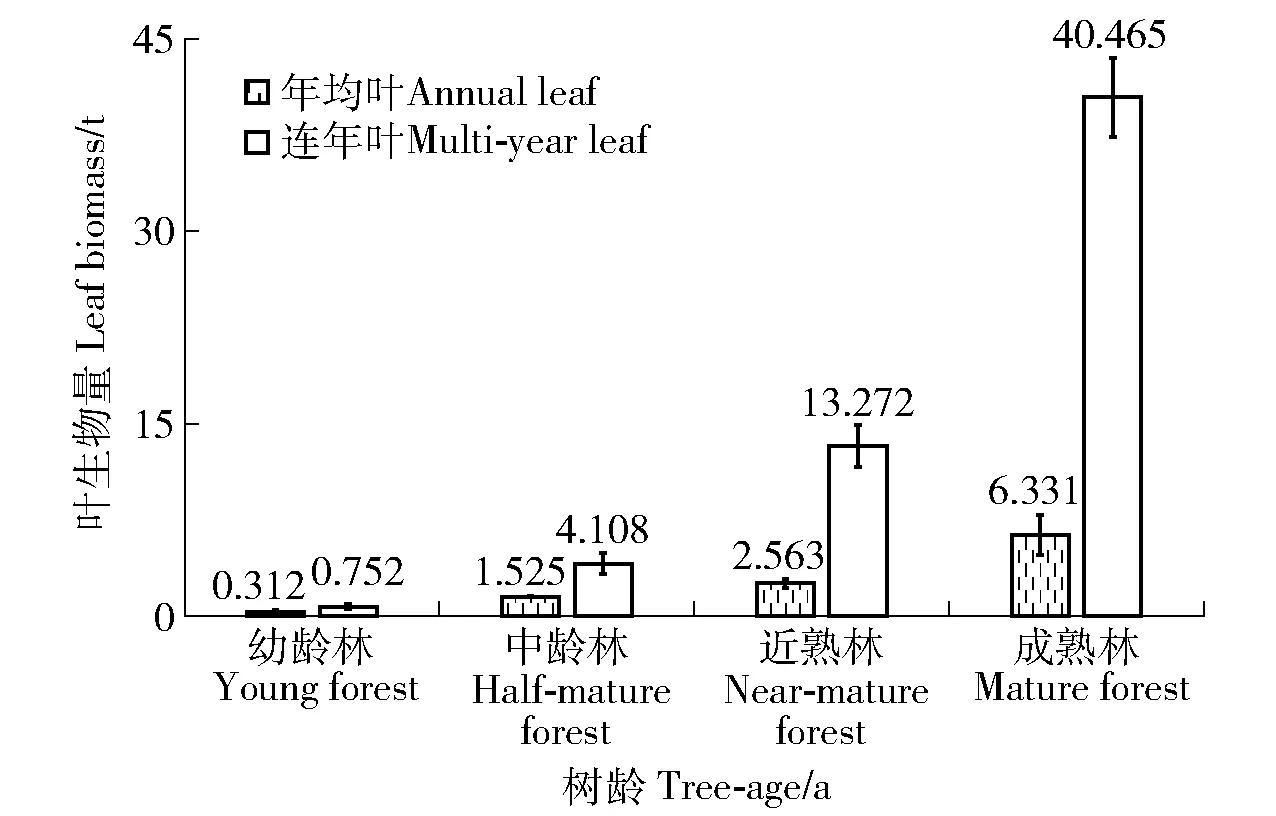
图4 不同林龄林带连年叶生物量与年均比较Fig.4 Ratio of multi-year leaf biomass to annual in different tree-age forest belts
3.3.3 林网年均和连年叶生物量比较 如表3所示,造林密度为68(2行3 m×7 m)、102(2行2 m×2 m)、137(2行1.5 m×3 m)和205株/hm2(2行1 m×2 m)的幼龄林单位面积林网年均叶生物量分别为(0.074 4±0.042 0)、(0.107 0±0.060 5)、(0.147 7±0.083 4)和(0.213 2±0.120 4)t/hm2,中龄林分别是幼龄林的5.0、4.6、4.8和4.5倍,近熟林分别是中龄林的1.7、1.6、1.7和1.6倍,成熟林分别是近熟林的2.5、2.3、2.4和2.0倍,增长倍数均在1.5倍以上,但涨幅呈先增大后减小再略增大的趋势。4种造林密度均长势良好,未出现抽稍现象。4种造林密度由小到大的幼龄林单位面积林网年均叶生物量净生产力分别为(0.010 6±0.006 0)、(0.015 3±0.008 6)、(0.021 1±0.011 9)和(0.030 5±0.017 2)t/(hm2·a),中龄林分别是幼龄林的3.2、2.9、3.1和2.9倍,近熟林分别时中龄林的1.2、1.1、1.1和1.2倍,成熟林分别是近熟林的1.9、1.7、1.9和1.5倍,增长倍数均>1,说明随林龄增长林网年均叶生物量净生产力呈增大趋势,但涨幅随林龄呈先增大后减小在略增大的趋势,中龄林涨幅出现极值;随着造林密度的增加林网年均叶生物量净生产力呈略减小趋势,但减幅不大。
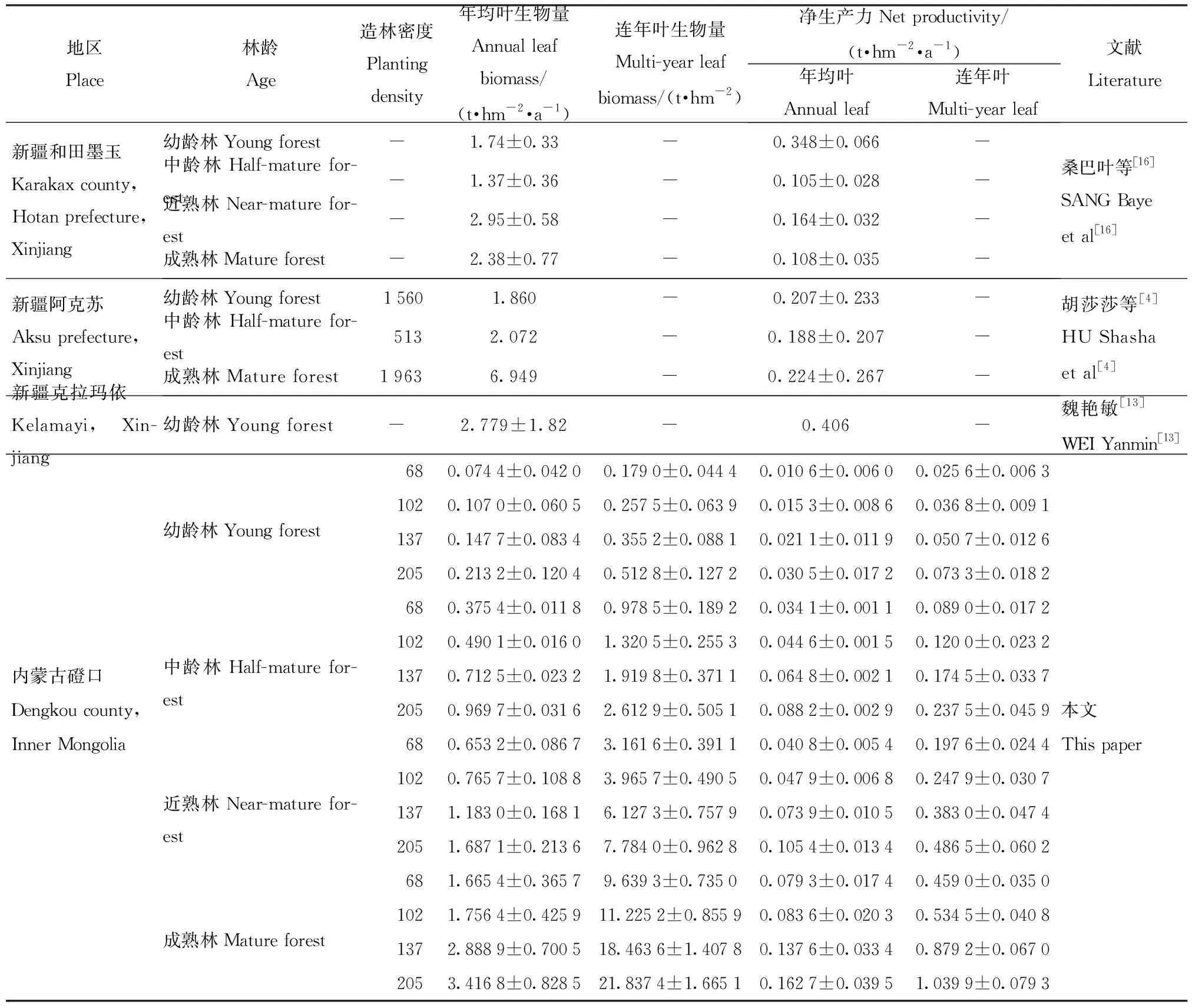
表3 林网年均和连年叶生物量及净生产力比较Tab.3 Comparison of leaf biomass and net productivity between annual forest network and multi-year
4种造林密度递增序列的幼龄林单位面积林网连年叶生物量分别为(0.179 0±0.044 4)、(0.257 5±0.063 9)、(0.355 2±0.088 1)和(0.512 8±0.127 2)t/hm2,中龄林分别是幼龄林的5.5、5.1、5.4和5.1倍,近熟林分别是中龄林的3.2、3.0、3.2和3.0倍,成熟林分别是近熟林的3.0、2.8、3.0、2.8,增长倍数均大于2.5倍,涨幅随林龄呈先增大后减小的趋势,说明随着林龄的增长林网年均叶生物量亦呈增大趋势;涨幅随着造林密度增加涨幅呈缩小的趋势。4种造林密度渐增排列的幼龄林单位面积林网连年叶生物量净生产力分别为(0.025 6±0.006 3)、(0.036 8±0.009 1)、(0.050 7±0.012 6)和(0.073 3±0.018 2)t/(hm2·a),中龄林分别是幼龄林的3.5、3.3、3.4和3.2倍,近熟林分别时中龄林的2.2、2.1、2.2和2.0倍,成熟林分别是近熟林的2.3、2.2、2.3和2.1倍,增长倍数均>2.0,说明随着林龄的增长林网连年叶生物量净生产力呈增大趋势,但涨幅随林龄呈先增大后减小再略增大的趋势,中龄林涨幅出现极值;随着造林密度的增加林网年均叶生物量净生产力呈略减小趋势,但减幅不大。
本研究4种造林密度幼龄林、中龄林、近熟林的年均叶生物量和净生产力均比以往研究[4,13-14,16]分别在新疆和田墨玉、新疆阿克苏、新疆克拉玛依的试验中得出的叶生物量低,但笔者取造林密度为137和205株/hm2,成熟林年均叶生物量和净生产力均高于桑巴叶等[16]得出的研究结果,年均叶生物量分别高出1.2和1.4倍,净生产力分别高出1.3和1.5倍。本研究与刘静[17]在该区域结构相近的林带得出基本相同的结果,更说明本文模型的可靠性。
本研究4种造林密度各林龄连年叶生物量目前并无研究无法横向比较,只能与年均叶生物量进行比较,除幼龄林和中龄林68和102株/hm2外,中龄林137和205株/hm2和近熟林、成熟林的连年生叶生物量均高于桑巴叶等[16]、胡莎莎等[4,14]、魏艳敏[13]的年均叶生物量。
4 结论
单株、林带、林网年均和连年叶生物量和净生产力随年龄和胸径的增长均呈增大趋势,涨幅均呈先增大后减小再略增大趋势,中龄林涨幅出现极值;4种林带结构在造林密度68~205株/hm2范围内,均生长良好,年均和连年叶生物量和净生产力均随着造林密度增加呈递增趋势,但涨幅呈减小的趋势。连年叶生物量及其涨幅均远大于年均叶生物量,且随年龄的增长,这种差距越来越大;由单株、林带、林网,随着空间尺度增大连年和年均叶生物量差距越来越大。
5 讨论
1)魏艳敏[13]在新疆得出相同形式胸径单因子叶生物量模型,但系数与本文模型相差较大,可能是因为区域气候水土条件差异造成的,虽模型相关系数更大但毕竟样本量太少,不具代表性,而本文样本量更大,模型对本区更具有指导意义。胡莎莎[14]在新疆墨玉得出相同形式胸径、树高二元叶生物量模型,但同样存在样本量较少和相关系数较低的问题,系数差异较大可能是由于气候水土条件、林分结构、样本量与本试区差异较大造成的。陈章水等[15]在新疆疏勒等所得模型与本文差异较大可能是区域和林分结构差异造成的。笔者取胸径22.85 cm的标准木年均叶生物量为8.343 kg与刘静[17]在临河相近胸径21.34~22.71得出的叶生物量8.980 kg相接近,说明本文的研究模型可靠。
2)年均叶生物量中龄林年均叶生物量增速最大可能是幼龄林树木根系生长完善;年均叶生物量生长后期增幅减小可能是树木生长彼此竞争加剧所致;成熟林叶生物量年增量最大可能与树木解锁新的营养空间有关。4种造林密度幼龄林、中龄林、近熟林的年均叶生物量和净生产力均比胡莎莎[4,14]、魏艳敏[13]、桑巴叶等[16]得出的叶生物量低,可能是本区农田防护林造林密度远低于新疆,因数量少所以幼龄林、中龄林、近熟林叶生物量和净生产力低于新疆,但成熟林本区叶生物量和净生产力远大于新疆,可能是由于本区低密度造林,充分利用了边行效应,营养空间较大,水分充足,所以生长后期叶生物量和净生产力竟然超过了新疆片林或多行林带高密度造林。
3)大部分学者叶生物量的研究均是年均叶生物量的研究,缺少连年叶生物量的研究,而本文是以林带结构参数为自变量因子的叶连年生物量及模型研究,这对于估算区域乃至全球尺度森林连年叶生物量产量、土壤碳储量、光合作用的累积、防护林的滞尘都有重要的参考价值。
