Efficacy of probiotics on the modulation of gut microbiota in the treatment of diabetic nephropathy
2022-03-23NozomiNagaseYukaIkedaAiTsujiYasukoKitagishiSatoruMatsuda
lNTRODUCTlON
Diabetic nephropathy(DN)is a chronic disorder occurring in nearly 40% of patients with diabetes[1].DN is an important cause of end-stage renal disease and a micro-vascular complication of diabetes mellitus(DM)[2,3].Some dietary factors might be involved in the increase in renal failure in association with DM,showing that the number of patients with DN and/or DM has been increasing in Asian countries because of westernization of dietary lifestyle[2,3].Pathogenesis of DN may be multifactorial and complex.Early DN has no noticeable clinical symptoms,however,hyperglycemia may be a significant risk factor for DN and/or DM[4].Sustained elevated blood glucose could lead to changes in the downstream transcription factors and/or gene expression in kidney glomerular cells[5].Kidney fibrosis and albu-minuria are key pathological processes of the advanced stage of DN[6],but oxidative stress and/or inflammation may also be important mechanisms for the pathogenesis of DN[7].In general,oxidative stress and inflammatory responses are almost not distinct,because one reaction would intensify the other pathogenesis.Both DM and chronic kidney disease(CKD)may have a common pathophysiological mechanism within a chronic inflammatory state and/or oxidative stresses[8].Among them,high levels of reactive oxygen species(ROS)could induce inflammatory cytokines in the kidney[9],which might accelerate the development of DN.Inflammation of the kidneys can lead to proteinuria and/or persistent hypertension,which can proceed to renal failure.Hence,successful treatment of the microcirculation in patients with DN has become a superior strategy for the prevention of DN.This reasonable treatment should be discovered immediately.Recently,it has been shown that pathogenesis of DN is associated with certain gut microbiota[10].The importance of probiotics is widely recognized in various diseases.Besides,studies have shown that crosstalk between host and microbiota might be relevant pathologically in patients with DN[11].For example,alterations in the gut microbiota are associated with the development of proteinuria[12],and type 2 DM[13].Changes to the gut microbiota have also been reported in DM and DN[14].The gut microbiota might well communicate with the kidneys,and the collapse of this relationship might result in the development of renal dysfunction.Accordingly,the gut microbiota could be an important defense against the pathogenesis of kidney disease.Dietary lifestyles have radically changed over the last century in developed countries,and are characterized by reduced dietary fiber and/or increased high-fat consumption[15].Hence,the changes could be linked to alteration of gut microbiota[16].Abnormal intestinal metabolites and disruption of the intestinal barrier owing to the gut dysbiosis might facilitate harmful substances produced in the gut entering the circulatory system[17].These situations allow us to hypothesize that dietary changes could lead to a microbiome that modifies positively the threshold and/or the speed of developing DN and/or DM.
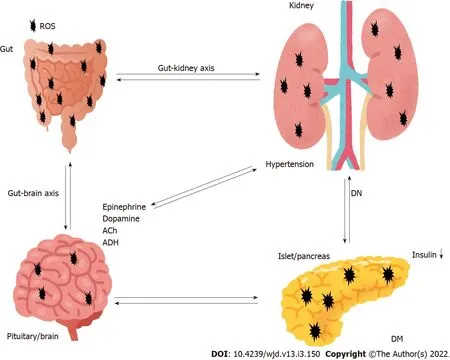
GUT-KlDNEY AXlS lN THE PATHOGENESlS OF DN
Although the significance of the gut microbiota has yet to be completely determined,it is obvious that an intricate symbiotic relationship might exist between host and microbe.In addition,the interaction has recently attracted interest in the study of the pathogenesis of various disorders.The human body holds numerous bacterial and/or microbial cells;the majority of which exist in the gut[18].The microbiota is a complex community of more than 100 trillion cells in healthy human intestines[19].The normal gut microbiota could protect the kidney,whereas gut dysbiosis of the microbiota could facilitate kidney disorders[20].Furthermore,alterations in the microbiota are gradually being linked to the development of various other diseases such as inflammatory bowel disease,cancer,psychiatric disorder,and cardiovascular disease[21].The gut-kidney axis could additionally affect metabolic and/or immune pathways in addition to the related diseases[22].The gut-kidney axis is largely mediated by metabolites produced by the gut microbiota,which might regulate physiological function of several organs including the brain,pancreas,adrenal glands,kidneys,
(Figure 1).For example,components of the immune system might have a key role with cytokines in communication between the gut and kidneys[23].Furthermore,crosstalk between the metabolic and immune pathways has a significant role in keeping a good balance in the kidneys[23].Intestinal responses to inflammation and/or infections are intricate.If microbiota-immune pathways overstimulate tolerance to some inflammation,greater inflammation may accelerate progression of renal disease and/or its complications.Accordingly,gut dysbiosis has frequently been associated with progression of many kidney diseases[24].In addition,accumulation of uremic toxins,which are derived from dietary metabolism in the gut and/or liver,has distinct effects on the kidneys.For example,the increase in urea increases its influx into the bowel lumen from epithelial cells,where it is hydrolyzed by gut microbiota urease to ammonia[25].Subsequently,ammonia byproducts may increase the bowel pH,leading to the severe mucosal damage[26].Accumulation of the uremic toxins in combination with inflammation may also increase the risk of renal disease[27].Therefore,key factors in kidney disease are function of the gut microbiota and/or the action of gut dysbiosis.Inflammatory bowel disease and DM are indeed multifactorial diseases,and both are chronic diseases associated with increased risk of various diseases including cardiovascular disease,which indicates that the gut is associated with host physiological functions[28].Interestingly,the prevalence of inflammatory bowel disease in adults with type 1 DM is higher compared to that of nondiabetic controls[29].It is plausible that the gut-kidney axis might be involved in the pathogenesis of inflammatory bowel disease and DM.Similarly,the gut microbiota may be involved in the damage of other organs,hence targeting the gut microbiota could represent a future therapeutic approach in various diseases.However,the potential impact of gastrointestinal-related disorders on the development and/or progression of DN remains to be elucidated.
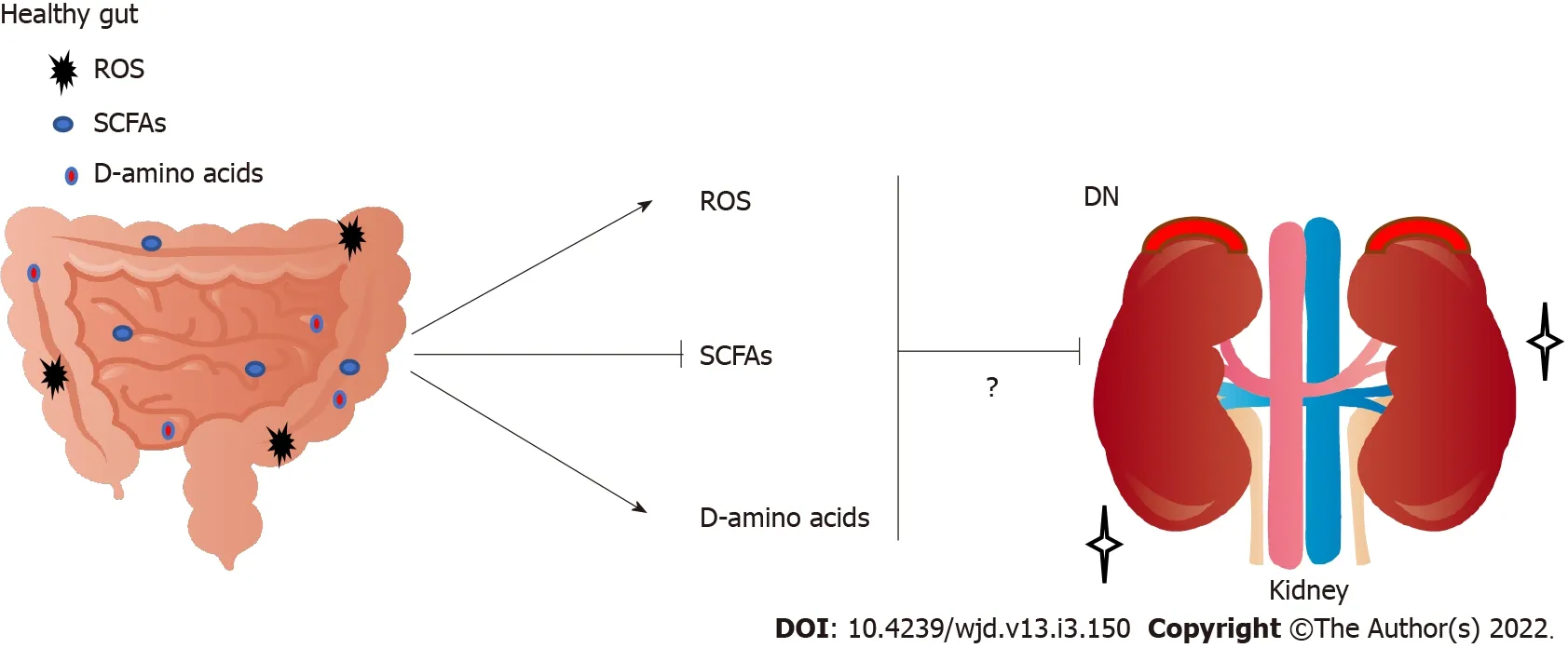
LEVELS OF SHORT-CHAlN FATTY AClDS,ROS,AND D-AMlNO AClDS MAY BE lNVOLVED lN THE DEVELOPMENT OF DN
Diabetic model mice fed with a high-fiber-diet are less likely to develop DN compared with diabetic control mice fed with a no-fiber diet[30].High-fiber diet might decrease the expression of genes encoding inflammatory cytokines related to DN[30].In general,fibers positively improve the dysbiosis of microbiota with promoting the production of short chain fatty acids(SCFAs)(including butyrate,acetate and propionate)in gut microbiota[31],which might also increase the production/release of cytokines and/or chemokines[32].In addition,SCFAs are able to inhibit intestinal inflammation and/or oxidative stress[33].Major SCFAs(acetate,propionate and butyrate)are derived through glycolysis of glucose to pyruvate or acetyl-CoA.The SCFAs regularly induce glucagon-like peptide 1 secretion through stimulation of a G-protein-coupled receptor(GPCR)[34].Gut microbiota in older people may weaken SCFA production[35].Those SCFAs have various effects on endocrine cells in gut
the GPCRs such as G-protein-coupled receptor(GPR)43 or GPR109A[36].SCFA-treated diabetic mice have been shown to be protected from nephropathy,suggesting that SCFAs protect renal cells from injury by oxidative stress in DN[37]
It has been shown that butyrate,one of the SCFAs produced by gut microbiota,plays a protective role in DN,which contributes in various physiological processes predominantly by inhibiting histone deacetylases(HDACs)[38].In addition,providing sodium butyrate has been shown to protect renal cells from oxidative damage and/or apoptosis in type 2 DN mice[39].Consistently,sodium butyrate has inhibited high-glucose-induced apoptosis of tubular epithelial cells in normal kidneys[40].Sodium butyrate also lowers plasma glucose and nuclear factor-B expression in the kidneys and attenuates kidney injury[41].In experimental mice,suppression of HDACs by sodium butyrate may explain the decrease in apoptosis in the kidneys[42].HDACs can regulate cell proliferation,migration and apoptosis,which are organized by a family of enzymes important for chromatin remodeling,keeping a dynamic balance with histone acetyltransferases in expression of several genes[43].Valproate,an HDAC inhibitor,has also been shown to decrease renal injury and/or renal fibrosis[44].
The signaling pathways triggered by hyperglycemia appear to have a pivotal role in diabetic complications due to the production of ROS and/or additional oxidative stress,which finally leads to apoptotic cell death in various tissues[45].ROS includes superoxide anions,hydroxyl free radicals,and hydrogen peroxide[46].The mitochon-drial electron transport chain is considered a major endogenous source of ROS[47].Production of excess ROS leads to increased membrane permeability and serious cellular damage[48].Such overproduction of ROS links to the pathological condition of altered metabolic pathways in the kidneys and disturbed renal function known as nephropathy[49].Once ATP synthesis is dysregulated in this hyperglycemic situation,it can result in excess production of ROS,which leads to kidney failure[50].Furthermore,high glucose exposure with excessive ROS can lead to renal podocyte apoptosis in experimental DN[51].Antioxidants including ubiquinone(also termed coenzyme Q10),ascorbic acid,and resveratrol have been tested in animal models of kidney diseases with some evidence of therapeutic benefits[52].Epidemiological studies have also found an association between high levels of ROS and risk of DN[53].Therefore,downregulation of ROS and/or oxidative stress might have a crucial role in regulating diabetic complications.Besides,ROS have been revealed to function as second messengers in several signal transduction pathways[54,55].
Studies have shown the clinical significance of D-amino acids in several kidney diseases[56].For example,the combination of blood level and urinary dynamics of D-serine effectively separates CKD from non-CKD[57].D-amino acids in body fluids are also a promising early detection marker for kidney disease[58].However,excess D-serine can cause kidney damage in rats[59].In this case,it has been shown that D-serine administration can initiate extensive necrosis in renal proximal tubules[59].In contrast,administration of D-alanine does not induce kidney injury[60].Furthermore,protective effects of low-dose D-serine have likely been shown to suppress renal damage,which may promote the hypoxia-mediated proliferation of tubular epithelial cells[61].In addition,D-cysteine administration can also protect the kidneys from ischemia-reperfusion injury,which might be useful to treat various renal diseases[62].D-aspartate plays a role during development and neurogenesis[63].D-aspartate treatment might produce favorable effects during demyelination and remyelination in the nervous system[64].Furthermore,the ovary-inducing activity of D-tryptophan is more effective than that of L-tryptophan[65].These data suggest that D-amino acids have both beneficial and harmful effects on tissue development and/or tissue-protection(Figure 2).
This lasted until they reached the avenue of orange trees, where were statues holding flaming torches, and when they got nearer to the palace they saw that it was illuminated from the roof to the ground, and music sounded softly from the courtyard
GUT MlCROBlOTA COULD CONTRlBUTE TO HEALTHY KlDNEYS
But, unfortunately, while I was still with her she became seriously ill, and though she presently recovered, her beauty is entirely11 gone, so that she hates the very sight of herself, and is in despair
This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers.It is distributed in accordance with the Creative Commons Attribution NonCommercial(CC BYNC 4.0)license,which permits others to distribute,remix,adapt,build upon this work non-commercially,and license their derivative works on different terms,provided the original work is properly cited and the use is noncommercial.See:https://creativecommons.org/Licenses/by-nc/4.0/
There are rooms and halls in it, but we do not enter them, weremain in the kitchen, where it is warm and light, clean and tidy; thecopper utensils13 are shining, the table as if polished with beeswax;the sink looks like a freshly scoured14 meatboard
The authors declare that they have no competing financial interests.
CONCLUSlON
When everything was tight closed the little hare turned to Big Lion and said Now! and Big Lion bounded out of the ditch and tore the other animals in pieces
Carbohydrates are metabolized by gut bacteria into monosaccharides and oligosaccharides,and they could be fermented into SCFAs.As shown above,SCFAs are one of the primary end products of gut fermentation that have considerable effects on host physiology.SCFAs can act as signaling molecules between the gut microbiota and host,and may have a protective effect on the renal function of patients with CKD.In particular,butyrate improves the intestinal barrier and reduces lipopolysaccharide influx into the blood,which could attenuate progression of DN[66].We provide here a perspective of gutkidney axis applied in search of renal disease management associated with the gut microbiome,which may theoretically be beneficial for future treatment of DN.Diet is known to be an essential regulator of gut microbiomes[67].Many studies have confirmed the association between nutrition and the human microbiome in maintaining human health,suggesting significant roles of bacterial metabolites in both health and disease[68].Trillions of bacteria present in the intestinal and colon lumina constitute the human gut microbiota[69].Dietary intake could control microbiota whose fermentation may produce various metabolites including SCFAs[70].The metabolites might additionally regulate the growth of pathogens by competing for of nutrients.For example,parenteral nutrition has been associated with a change in the microbiota,altering SCFA production,and inducing gut mucosal atrophy[71].The SCFAs made by the healthy gut microbiota have anti-inflammatory properties,including proliferation of regulatory T cells[72,73].In addition,a significant role for regulatory T cells has been revealed in type 2 diabetes for protection against DN[74].In addition,SCFAs have favorable effects on β cells,potentiating glucose-stimulated insulin release and/or maintaining β-cell mass through inhibiting apoptosis[75].Furthermore,propionate,has been shown to prevent adipogenic differentiation of specific stem cells[76].
Japan
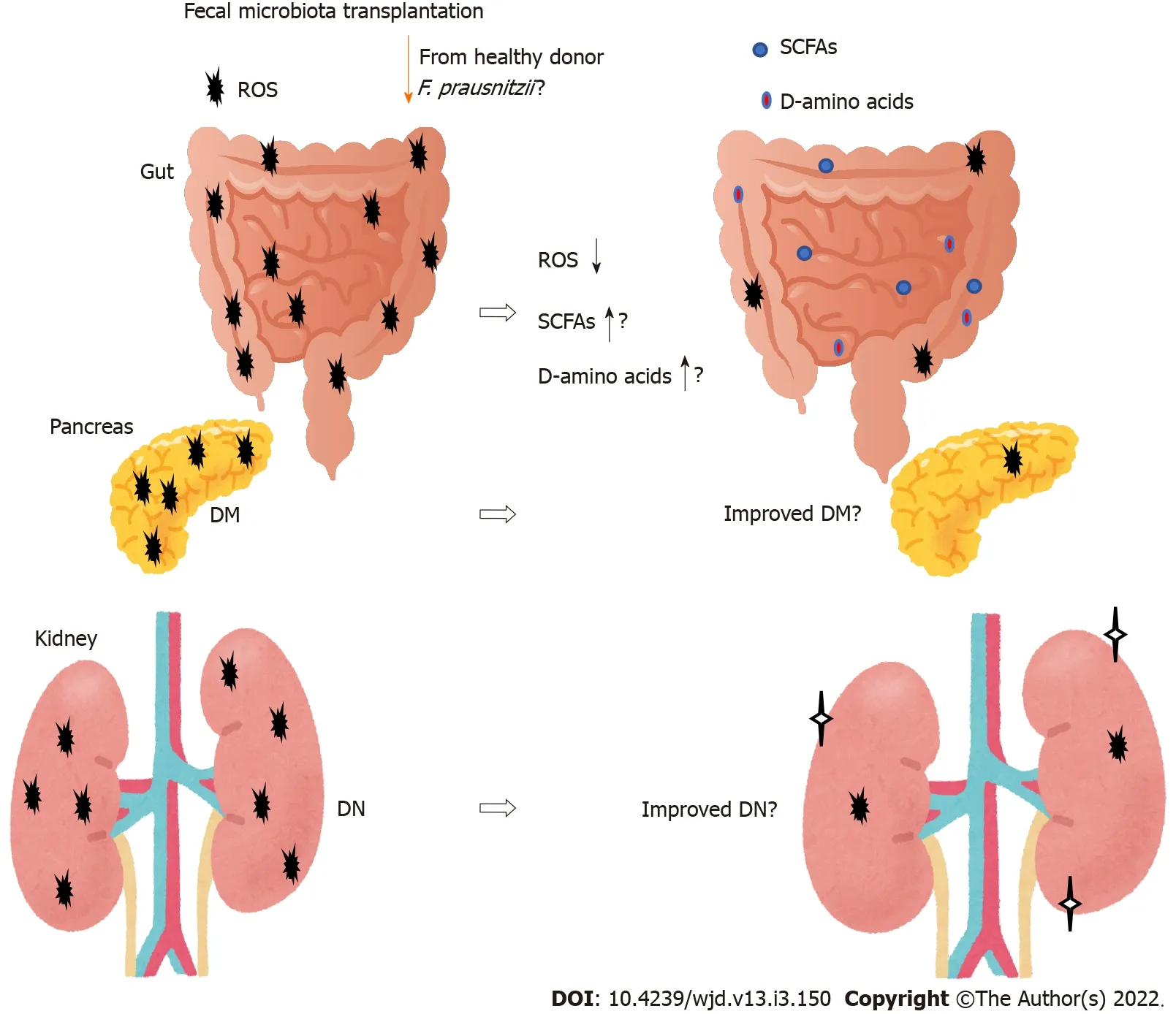
New therapies for DN are emerging.One method that may affect the gut microbiota composition is fecal microbiota transplantation(FMT)(Figure 3).The beneficial effects of the transplantation are dependent on the host responses,however,which may provide a potential treatment strategy for type 2 diabetes[93].In particular,transplantation of
(
)could restore the intestinal structure,which might be used as a potential therapeutic approach against inflammation as well as diabetes[94-96].Furthermore,
may serve as a diagnostic and therapeutic biomarker for the use of FMT[97].The potential role of the gut microbiota has been hypothesized to modulate renal function in experimental DN murine models[98].Through FMT,the role of the gut microbiota and its SCFA production have been verified in the treatment of DN.Therefore,administration of prebiotics and/or probiotics should individually be tailor-made to prevent and/or cure chronic diseases such as DN.For example,acetate produced by certain gut microbiota reprogramming has been shown to contribute to the tubulointerstitial injury of DN,suggesting that gut microbiota might be a new strategy for DN treatment[99].Furthermore,FMT from healthy donors considerably attenuates glomerular injury with podocyte improvement in diabetic rats[100].
FOOTNOTES
Each author has participated sufficiently in the work of drafting the article and/or revising the article for important rational content;all authors give final approval of the version to be submitted.
Finally,the gut microbiota have the largest genetic capacity to metabolize D-amino acids that are utilized as nutrients to support bacterial growth to regulate spore germination[86].Therefore,one possible source of D-amino acids in mammals may be their gut microbiota.In general,many bacterial species encode racemases that convert L-amino acids to D-amino acids[87].For example,D-alanine production is associated with a relative abundance of bacterial species with racemases such as those of
and
in the gut microbiota[88].Different bacterial species may produce distinct profiles of D-amino acids[89].Higher D-amino acids levels have been related to the gut microbial mass[90].Oral intake of a peptide containing specific D-amino acids may reverse the diabetes-associated pathological alterations in the kidneys[91](Figure 2).Noteworthy differences in the microbiota composition have been discovered in patients with kidney disease compared with healthy controls[92].Consequently,treatment options for DN should include dietary therapy affecting the gut microbiota.Therapeutic interventions would nevertheless represent a potential target of the microbiota for prevention and/or treatment of DN.
Many studies have emphasized the relationship between the gut microbiota and oxidative stress[77].In general,ROS production has a defense mechanism that could elicit cytotoxicity against several pathogens then reduce the burden of infection[78].Redox signaling is also found in response to microbial signals
the gut epithelial NADPH oxidase 1[79].Therefore,microbial ROS might rigorously control signaling processes for appropriate immunity and/or the gut barrier[80].Numerous bacterial species of the microbiota can reduce mitochondrial ROS production[81].For example,microbial products can upregulate the activity of superoxide dismutase,which results in reduced ROS levels and then decreased cellular apoptosis[82].In addition,microbial excess ROS might disturb other important pathways of host cells,suggesting that ROS-mediated signaling can regulate various cellular processes in order to keep the host healthy[83].Epithelial cells may also exhibit increased ROS production in response to several harmful bacteria[84].In the gut,epithelial appropriate ROS production in response to the gut bacteria may play a signaling role in the host[85].It is likely that there are many ROS-sensitive important enzymes that could be affected by alterations in the gut redox conditions.
The above-mentioned topics are only just being explored in preclinical research,suggesting that further studies are required.Owing to a lack of treatments,DN has been a public health concern.Although it is untimely to draw definitive conclusions about the clinical usefulness of microbiota-based treatment strategies for DN,modulation of gut microbiota is an exciting frontier in kidney research.It is clear that intensive evaluation of preclinical studies is necessary to find further insights.In addition,long-term studies are also necessary to clarify the detailed effects of probiotic treatment in the management of DN.A healthy lifestyle with a balanced familiar diet is now one of the main recommendations.
He was struck with astonishment69 at the sight of the chariot, and was gazing at it, when the Enchanter strode up to him, exclaiming: Shake hands, Cloverleaf, old fellow! Don t you know me? No, I can t say I do, replied the King, somewhat embarrassed
Nozomi Nagase 0000-0003-3665-5714;Yuka Ikeda 0000-0003-4805-1758;Ai Tsuji 0000-0003-1619-7592;Yasuko Kitagishi 0000-0002-6906-7444;Satoru Matsuda 0000-0003-4274-5345.
Wang JJ
Kerr C
Wang JJ
1 Du X,Liu J,Xue Y,Kong X,Lv C,Li Z,Huang Y,Wang B.Alteration of gut microbial profile in patients with diabetic nephropathy.
2021;73:71-84[PMID:33905112 DOI:10.1007/s12020-021-02721-1]
2 van den Berg E,Hospers FA,Navis G,Engberink MF,Brink EJ,Geleijnse JM,van Baak MA,Gans RO,Bakker SJ.Dietary acid load and rapid progression to end-stage renal disease of diabetic nephropathy in Westernized South Asian people.
2011;24:11-17[PMID:20872351 DOI:10.5301/jn.2010.5711]
3 Alicic RZ,Johnson EJ,Tuttle KR.Inflammatory Mechanisms as New Biomarkers and Therapeutic Targets for Diabetic Kidney Disease.
2018;25:181-191[PMID:29580582 DOI:10.1053/j.ackd.2017.12.002]
4 Vergès B.Cardiovascular disease in type 1 diabetes:A review of epidemiological data and underlying mechanisms.
2020;46:442-449[PMID:32998054 DOI:10.1016/j.diabet.2020.09.001]
5 Nordquist L,Friederich-Persson M,Fasching A,Liss P,Shoji K,Nangaku M,Hansell P,Palm F.Activation of hypoxiainducible factors prevents diabetic nephropathy.
2015;26:328-338[PMID:25183809 DOI:10.1681/ASN.2013090990]
6 Christou GA,Kiortsis DN.The role of adiponectin in renal physiology and development of albuminuria.
2014;221:R49-R61[PMID:24464020 DOI:10.1530/JOE-13-0578]
7 Navarro-González JF,Mora-Fernández C,Muros de Fuentes M,García-Pérez J.Inflammatory molecules and pathways in the pathogenesis of diabetic nephropathy.
2011;7:327-340[PMID:21537349 DOI:10.1038/nrneph.2011.51]
8 Tanase DM,Gosav EM,Neculae E,Costea CF,Ciocoiu M,Hurjui LL,Tarniceriu CC,Maranduca MA,Lacatusu CM,Floria M,Serban IL.Role of Gut Microbiota on Onset and Progression of Microvascular Complications of Type 2 Diabetes(T2DM).
2020;12[PMID:33276482 DOI:10.3390/nu12123719]
9 Justin Rucker A,Crowley SD.The role of macrophages in hypertension and its complications.
2017;469:419-430[PMID:28251313 DOI:10.1007/s00424-017-1950-x]
10 Tesch GH.Diabetic nephropathy - is this an immune disorder?
2017;131:2183-2199[PMID:28760771 DOI:10.1042/CS20160636]
11 Fernandes R,Viana SD,Nunes S,Reis F.Diabetic gut microbiota dysbiosis as an inflammaging and immunosenescence condition that fosters progression of retinopathy and nephropathy.
2019;1865:1876-1897[PMID:30287404 DOI:10.1016/j.bbadis.2018.09.032]
12 Yoshifuji A,Wakino S,Irie J,Tajima T,Hasegawa K,Kanda T,Tokuyama H,Hayashi K,Itoh H.Gut Lactobacillus protects against the progression of renal damage by modulating the gut environment in rats.
2016;31:401-412[PMID:26487672 DOI:10.1093/ndt/gfv353]
13 Wen L,Duffy A.Factors Influencing the Gut Microbiota,Inflammation,and Type 2 Diabetes.
2017;147:1468S-1475S[PMID:28615382 DOI:10.3945/jn.116.240754]
14 Yang J,Dong H,Wang Y,Jiang Y,Zhang W,Lu Y,Chen Y,Chen L.Cordyceps cicadae polysaccharides ameliorated renal interstitial fibrosis in diabetic nephropathy rats by repressing inflammation and modulating gut microbiota dysbiosis.
2020;163:442-456[PMID:32592781 DOI:10.1016/j.ijbiomac.2020.06.153]
15 Maslowski KM,Mackay CR.Diet,gut microbiota and immune responses.
2011;12:5-9[PMID:21169997 DOI:10.1038/ni0111-5]
16 Andoh A,Kuzuoka H,Tsujikawa T,Nakamura S,Hirai F,Suzuki Y,Matsui T,Fujiyama Y,Matsumoto T.Multicenter analysis of fecal microbiota profiles in Japanese patients with Crohn's disease.
2012;47:1298-1307[PMID:22576027 DOI:10.1007/s00535-012-0605-0]
17 Yang G,Wei J,Liu P,Zhang Q,Tian Y,Hou G,Meng L,Xin Y,Jiang X.Role of the gut microbiota in type 2 diabetes and related diseases.
2021;117:154712[PMID:33497712 DOI:10.1016/j.metabol.2021.154712]
18 Sender R,Fuchs S,Milo R.Revised Estimates for the Number of Human and Bacteria Cells in the Body.
2016;14:e1002533[PMID:27541692 DOI:10.1371/journal.pbio.1002533]
19 De Sordi L,Khanna V,Debarbieux L.The Gut Microbiota Facilitates Drifts in the Genetic Diversity and Infectivity of Bacterial Viruses.
2017;22:801-808.e3[PMID:29174401 DOI:10.1016/j.chom.2017.10.010]
20 Mahmoodpoor F,Rahbar Saadat Y,Barzegari A,Ardalan M,Zununi Vahed S.The impact of gut microbiota on kidney function and pathogenesis.
2017;93:412-419[PMID:28654798 DOI:10.1016/j.biopha.2017.06.066]
21 Li DY,Tang WHW.Contributory Role of Gut Microbiota and Their Metabolites Toward Cardiovascular Complications in Chronic Kidney Disease.
2018;38:193-205[PMID:29602401 DOI:10.1016/j.semnephrol.2018.01.008]
22 Evenepoel P,Poesen R,Meijers B.The gut-kidney axis.
2017;32:2005-2014[PMID:27848096 DOI:10.1007/s00467-016-3527-x]
23 Yang T,Richards EM,Pepine CJ,Raizada MK.The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease.
2018;14:442-456[PMID:29760448 DOI:10.1038/s41581-018-0018-2]
24 Afsar B,Vaziri ND,Aslan G,Tarim K,Kanbay M.Gut hormones and gut microbiota:implications for kidney function and hypertension.
2016;10:954-961[PMID:27865823 DOI:10.1016/j.jash.2016.10.007]
25 Vaziri ND,Yuan J,Khazaeli M,Masuda Y,Ichii H,Liu S.Oral activated charcoal adsorbent(AST-120)ameliorates chronic kidney disease-induced intestinal epithelial barrier disruption.
2013;37:518-525[PMID:23689670 DOI:10.1159/000351171]
26 Figura N.Helicobacter pylori factors involved in the development of gastroduodenal mucosal damage and ulceration.
1997;25 Suppl 1:S149-S163[PMID:9479642 DOI:10.1097/00004836-199700001-00025]
27 Wu PH,Lin YT,Chiu YW,Baldanzi G,Huang JC,Liang SS,Lee SC,Chen SC,Hsu YL,Kuo MC,Hwang SJ.The relationship of indoxyl sulfate and p-cresyl sulfate with target cardiovascular proteins in hemodialysis patients.
2021;11:3786[PMID:33589722 DOI:10.1038/s41598-021-83383-x]
28 Torkamani A,Topol EJ,Schork NJ.Pathway analysis of seven common diseases assessed by genome-wide association.
2008;92:265-272[PMID:18722519 DOI:10.1016/j.ygeno.2008.07.011]
29 Lu S,Gong J,Tan Y,Liu D.Epidemiologic Association between Inflammatory Bowel Diseases and Type 1 Diabetes Mellitus:a Meta-Analysis.
2020;29:407-413[PMID:32919423 DOI:10.15403/jgld-798]
30 Li YJ,Chen X,Kwan TK,Loh YW,Singer J,Liu Y,Ma J,Tan J,Macia L,Mackay CR,Chadban SJ,Wu H.Dietary Fiber Protects against Diabetic Nephropathy through Short-Chain Fatty Acid-Mediated Activation of G Protein-Coupled Receptors GPR43 and GPR109A.
2020;31:1267-1281[PMID:32358041 DOI:10.1681/ASN.2019101029]
31 Bai Y,Li Y,Marion T,Tong Y,Zaiss MM,Tang Z,Zhang Q,Liu Y,Luo Y.Resistant starch intake alleviates collageninduced arthritis in mice by modulating gut microbiota and promoting concomitant propionate production.
2021;116:102564[PMID:33203617 DOI:10.1016/j.jaut.2020.102564]
32 Rutting S,Xenaki D,Malouf M,Horvat JC,Wood LG,Hansbro PM,Oliver BG.Short-chain fatty acids increase TNFαinduced inflammation in primary human lung mesenchymal cells through the activation of p38 MAPK.
2019;316:L157-L174[PMID:30407866 DOI:10.1152/ajplung.00306.2018]
33 Huang W,Guo HL,Deng X,Zhu TT,Xiong JF,Xu YH,Xu Y.Short-Chain Fatty Acids Inhibit Oxidative Stress and Inflammation in Mesangial Cells Induced by High Glucose and Lipopolysaccharide.
2017;125:98-105[PMID:28049222 DOI:10.1055/s-0042-121493]
34 Zhou D,Chen YW,Zhao ZH,Yang RX,Xin FZ,Liu XL,Pan Q,Zhou H,Fan JG.Sodium butyrate reduces high-fat dietinduced non-alcoholic steatohepatitis through upregulation of hepatic GLP-1R expression.
2018;50:1-12[PMID:30510243 DOI:10.1038/s12276-018-0183-1]
35 Rampelli S,Candela M,Turroni S,Biagi E,Collino S,Franceschi C,O'Toole PW,Brigidi P.Functional metagenomic profiling of intestinal microbiome in extreme ageing.
2013;5:902-912[PMID:24334635 DOI:10.18632/aging.100623]
36 Moniri NH,Farah Q.Short-chain free-fatty acid G protein-coupled receptors in colon cancer.
2021;186:114483[PMID:33631190 DOI:10.1016/j.bcp.2021.114483]
37 Andrade-Oliveira V,Amano MT,Correa-Costa M,Castoldi A,Felizardo RJ,de Almeida DC,Bassi EJ,Moraes-Vieira PM,Hiyane MI,Rodas AC,Peron JP,Aguiar CF,Reis MA,Ribeiro WR,Valduga CJ,Curi R,Vinolo MA,Ferreira CM,Câmara NO.Gut Bacteria Products Prevent AKI Induced by Ischemia-Reperfusion.
2015;26:1877-1888[PMID:25589612 DOI:10.1681/ASN.2014030288]
38 Felizardo RJF,de Almeida DC,Pereira RL,Watanabe IKM,Doimo NTS,Ribeiro WR,Cenedeze MA,Hiyane MI,Amano MT,Braga TT,Ferreira CM,Parmigiani RB,Andrade-Oliveira V,Volpini RA,Vinolo MAR,Mariño E,Robert R,Mackay CR,Camara NOS.Gut microbial metabolite butyrate protects against proteinuric kidney disease through epigenetic- and GPR109a-mediated mechanisms.
2019;33:11894-11908[PMID:31366236 DOI:10.1096/fj.201901080R]
39 Dong W,Jia Y,Liu X,Zhang H,Li T,Huang W,Chen X,Wang F,Sun W,Wu H.Sodium butyrate activates NRF2 to ameliorate diabetic nephropathy possibly
inhibition of HDAC.
2017;232:71-83[PMID:27799462 DOI:10.1530/JOE-16-0322]
40 Du Y,Tang G,Yuan W.Suppression of HDAC2 by sodium butyrate alleviates apoptosis of kidney cells in db/db mice and HGinduced NRK52E cells.
2020;45:210-222[PMID:31746362 DOI:10.3892/ijmm.2019.4397]
41 Khan S,Jena G.Sodium butyrate,a HDAC inhibitor ameliorates eNOS,iNOS and TGF-β1-induced fibrogenesis,apoptosis and DNA damage in the kidney of juvenile diabetic rats.
2014;73:127-139[PMID:25158305 DOI:10.1016/j.fct.2014.08.010]
42 Kim SW,Hooker JM,Otto N,Win K,Muench L,Shea C,Carter P,King P,Reid AE,Volkow ND,Fowler JS.Wholebody pharmacokinetics of HDAC inhibitor drugs,butyric acid,valproic acid and 4-phenylbutyric acid measured with carbon-11 labeled analogs by PET.
2013;40:912-918[PMID:23906667 DOI:10.1016/j.nucmedbio.2013.06.007]
43 Choudhary C,Kumar C,Gnad F,Nielsen ML,Rehman M,Walther TC,Olsen JV,Mann M.Lysine acetylation targets protein complexes and co-regulates major cellular functions.
2009;325:834-840[PMID:19608861 DOI:10.1126/science.1175371]
44 Khan S,Jena G,Tikoo K.Sodium valproate ameliorates diabetes-induced fibrosis and renal damage by the inhibition of histone deacetylases in diabetic rat.
2015;98:230-239[PMID:25576297 DOI:10.1016/j.yexmp.2015.01.003]
45 Sávio-Silva C,Soinski-Sousa PE,Simplício-Filho A,Bastos RMC,Beyerstedt S,Rangel ÉB.Therapeutic Potential of Mesenchymal Stem Cells in a Pre-Clinical Model of Diabetic Kidney Disease and Obesity.
2021;22[PMID:33557007 DOI:10.3390/ijms22041546]
46 Ahmad R,Ahsan H.Singlet oxygen species and systemic lupus erythematosus:a brief review.
2019;40:343-349[PMID:31116079 DOI:10.1080/15321819.2019.1616555]
47 Auger C,Vinaik R,Appanna VD,Jeschke MG.Beyond mitochondria:Alternative energy-producing pathways from all strata of life.
2021;118:154733[PMID:33631145 DOI:10.1016/j.metabol.2021.154733]
48 Bonora M,Patergnani S,Ramaccini D,Morciano G,Pedriali G,Kahsay AE,Bouhamida E,Giorgi C,Wieckowski MR,Pinton P.Physiopathology of the Permeability Transition Pore:Molecular Mechanisms in Human Pathology.
2020;10[PMID:32635556 DOI:10.3390/biom10070998]
49 Jha JC,Banal C,Chow BS,Cooper ME,Jandeleit-Dahm K.Diabetes and Kidney Disease:Role of Oxidative Stress.
2016;25:657-684[PMID:26906673 DOI:10.1089/ars.2016.6664]
50 Badal SS,Danesh FR.New insights into molecular mechanisms of diabetic kidney disease.
2014;63:S63-S83[PMID:24461730 DOI:10.1053/j.ajkd.2013.10.047]
51 Susztak K,Raff AC,Schiffer M,Böttinger EP.Glucose-induced reactive oxygen species cause apoptosis of podocytes and podocyte depletion at the onset of diabetic nephropathy.
2006;55:225-233[PMID:16380497]
52 Huang SS,Ding DF,Chen S,Dong CL,Ye XL,Yuan YG,Feng YM,You N,Xu JR,Miao H,You Q,Lu X,Lu YB.Resveratrol protects podocytes against apoptosis
stimulation of autophagy in a mouse model of diabetic nephropathy.
2017;7:45692[PMID:28374806 DOI:10.1038/srep45692]
53 Khan SR.Is oxidative stress,a link between nephrolithiasis and obesity,hypertension,diabetes,chronic kidney disease,metabolic syndrome?
2012;40:95-112[PMID:22213019 DOI:10.1007/s00240-011-0448-9]
54 Chiarugi P,Buricchi F.Protein tyrosine phosphorylation and reversible oxidation:two cross-talking posttranslation modifications.
2007;9:1-24[PMID:17115885 DOI:10.1089/ars.2007.9.1]
55 Linnane AW,Kios M,Vitetta L.Healthy aging:regulation of the metabolome by cellular redox modulation and prooxidant signaling systems:the essential roles of superoxide anion and hydrogen peroxide.
2007;8:445-467[PMID:17415678 DOI:10.1007/s10522-007-9096-4]
56 Kaimori JY,Maehara K,Hayashi-Takanaka Y,Harada A,Fukuda M,Yamamoto S,Ichimaru N,Umehara T,Yokoyama S,Matsuda R,Ikura T,Nagao K,Obuse C,Nozaki N,Takahara S,Takao T,Ohkawa Y,Kimura H,Isaka Y.Histone H4 lysine 20 acetylation is associated with gene repression in human cells.
2016;6:24318[PMID:27064113 DOI:10.1038/srep24318]
57 Hesaka A,Sakai S,Hamase K,Ikeda T,Matsui R,Mita M,Horio M,Isaka Y,Kimura T.
-Serine reflects kidney function and diseases.
2019;9:5104[PMID:30911057 DOI:10.1038/s41598-019-41608-0]
58 Kimura T,Hamase K,Miyoshi Y,Yamamoto R,Yasuda K,Mita M,Rakugi H,Hayashi T,Isaka Y.Chiral amino acid metabolomics for novel biomarker screening in the prognosis of chronic kidney disease.
2016;6:26137[PMID:27188851 DOI:10.1038/srep26137]
59 Ganote CE,Peterson DR,Carone FA.The nature of D-serine--induced nephrotoxicity.
1974;77:269-282[PMID:4447130]
60 Maekawa M,Okamura T,Kasai N,Hori Y,Summer KH,Konno R.D-amino-acid oxidase is involved in D-serineinduced nephrotoxicity.
2005;18:1678-1682[PMID:16300376 DOI:10.1021/tx0500326]
61 Nakade Y,Iwata Y,Furuichi K,Mita M,Hamase K,Konno R,Miyake T,Sakai N,Kitajima S,Toyama T,Shinozaki Y,Sagara A,Miyagawa T,Hara A,Shimizu M,Kamikawa Y,Sato K,Oshima M,Yoneda-Nakagawa S,Yamamura Y,Kaneko S,Miyamoto T,Katane M,Homma H,Morita H,Suda W,Hattori M,Wada T.Gut microbiota-derived D-serine protects against acute kidney injury.
2018;3[PMID:30333299 DOI:10.1172/jci.insight.97957]
62 Kimura H.The physiological role of hydrogen sulfide and beyond.
2014;41:4-10[PMID:24491257 DOI:10.1016/j.niox.2014.01.002]
63 van den Pol AN,Obrietan K,Cao V,Trombley PQ.Embryonic hypothalamic expression of functional glutamate receptors.
1995;67:419-439[PMID:7545794 DOI:10.1016/0306-4522(95)96912-w]
64 de Rosa V,Secondo A,Pannaccione A,Ciccone R,Formisano L,Guida N,Crispino R,Fico A,Polishchuk R,D'Aniello A,Annunziato L,Boscia F.D-Aspartate treatment attenuates myelin damage and stimulates myelin repair.
2019;11[PMID:30559305 DOI:10.15252/emmm.201809278]
65 Kobayashi K,Maezawa T,Tanaka H,Onuki H,Horiguchi Y,Hirota H,Ishida T,Horiike K,Agata Y,Aoki M,Hoshi M,Matsumoto M.The identification of D-tryptophan as a bioactive substance for postembryonic ovarian development in the planarian Dugesia ryukyuensis.
2017;7:45175[PMID:28338057 DOI:10.1038/srep45175]
66 Sabatino A,Regolisti G,Cosola C,Gesualdo L,Fiaccadori E.Intestinal Microbiota in Type 2 Diabetes and Chronic Kidney Disease.
2017;17:16[PMID:28271466 DOI:10.1007/s11892-017-0841-z]
67 Chen PB,Black AS,Sobel AL,Zhao Y,Mukherjee P,Molparia B,Moore NE,Aleman Muench GR,Wu J,Chen W,Pinto AFM,Maryanoff BE,Saghatelian A,Soroosh P,Torkamani A,Leman LJ,Ghadiri MR.Directed remodeling of the mouse gut microbiome inhibits the development of atherosclerosis.
2020;38:1288-1297[PMID:32541956 DOI:10.1038/s41587-020-0549-5]
68 Cotillard A,Kennedy SP,Kong LC,Prifti E,Pons N,Le Chatelier E,Almeida M,Quinquis B,Levenez F,Galleron N,Gougis S,Rizkalla S,Batto JM,Renault P;ANR MicroObes consortium,Doré J,Zucker JD,Clément K,Ehrlich SD.Dietary intervention impact on gut microbial gene richness.
2013;500:585-588[PMID:23985875 DOI:10.1038/nature12480]
69 Guo Y,Kitamoto S,Kamada N.Microbial adaptation to the healthy and inflamed gut environments.
2020;12:1857505[PMID:33382358 DOI:10.1080/19490976.2020.1857505]
70 Shanahan F,van Sinderen D,O'Toole PW,Stanton C.Feeding the microbiota:transducer of nutrient signals for the host.
2017;66:1709-1717[PMID:28663354 DOI:10.1136/gutjnl-2017-313872]
71 Ríos-Covián D,Ruas-Madiedo P,Margolles A,Gueimonde M,de Los Reyes-Gavilán CG,Salazar N.Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health.
2016;7:185[PMID:26925050 DOI:10.3389/fmicb.2016.00185]
72 Papatriantafyllou M.T cells:maintaining T cell homeostasis.
2013;13:546-547[PMID:23868219 DOI:10.1038/nri3504]
73 Papatriantafyllou M.Regulatory T cells:distilling regulatory T cell inducers.
2013;13:546[PMID:24046840 DOI:10.1038/nri3506]
74 Abouzeid S,Sherif N.Role of alteration in Treg/Th17 cells' balance in nephropathic patients with Type 2 diabetes mellitus.
2015;7:1613-1618[PMID:26816588 DOI:10.19082/1613]
75 Pingitore A,Chambers ES,Hill T,Maldonado IR,Liu B,Bewick G,Morrison DJ,Preston T,Wallis GA,Tedford C,Castañera González R,Huang GC,Choudhary P,Frost G,Persaud SJ.The diet-derived short chain fatty acid propionate improves beta-cell function in humans and stimulates insulin secretion from human islets in vitro.
2017;19:257-265[PMID:27761989 DOI:10.1111/dom.12811]
76 Iván J,Major E,Sipos A,Kovács K,Horváth D,Tamás I,Bay P,Dombrádi V,Lontay B.The Short-Chain Fatty Acid Propionate Inhibits Adipogenic Differentiation of Human Chorion-Derived Mesenchymal Stem Cells Through the Free Fatty Acid Receptor 2.
2017;26:1724-1733[PMID:28992793 DOI:10.1089/scd.2017.0035]
77 Kong Y,Olejar KJ,On SLW,Chelikani V.The Potential of
spp.for Modulating Oxidative Stress in the Gastrointestinal Tract.
2020;9[PMID:32664392 DOI:10.3390/antiox9070610]
78 Ghosh S,Dai C,Brown K,Rajendiran E,Makarenko S,Baker J,Ma C,Halder S,Montero M,Ionescu VA,Klegeris A,Vallance BA,Gibson DL.Colonic microbiota alters host susceptibility to infectious colitis by modulating inflammation,redox status,and ion transporter gene expression.
2011;301:G39-G49[PMID:21454446 DOI:10.1152/ajpgi.00509.2010]
79 Neish AS.Redox signaling mediated by the gut microbiota.
2013;47:950-957[PMID:23937589 DOI:10.3109/10715762.2013.833331]
80 Patel RM,Myers LS,Kurundkar AR,Maheshwari A,Nusrat A,Lin PW.Probiotic bacteria induce maturation of intestinal claudin 3 expression and barrier function.
2012;180:626-635[PMID:22155109 DOI:10.1016/j.ajpath.2011.10.025]
81 Lobet E,Letesson JJ,Arnould T.Mitochondria:a target for bacteria.
2015;94:173-185[PMID:25707982 DOI:10.1016/j.bcp.2015.02.007]
82 Liu TF,Vachharajani VT,Yoza BK,McCall CE.NAD+-dependent sirtuin 1 and 6 proteins coordinate a switch from glucose to fatty acid oxidation during the acute inflammatory response.
2012;287:25758-25769[PMID:22700961 DOI:10.1074/jbc.M112.362343]
83 Belizário JE,Faintuch J,Garay-Malpartida M.Gut Microbiome Dysbiosis and Immunometabolism:New Frontiers for Treatment of Metabolic Diseases.
2018;2018:2037838[PMID:30622429 DOI:10.1155/2018/2037838]
84 Ha EM,Oh CT,Bae YS,Lee WJ.A direct role for dual oxidase in Drosophila gut immunity.
2005;310:847-850[PMID:16272120 DOI:10.1126/science.1117311]
85 Neish AS,Jones RM.Redox signaling mediates symbiosis between the gut microbiota and the intestine.
2014;5:250-253[PMID:24637602 DOI:10.4161/gmic.27917]
86 Cava F,Lam H,de Pedro MA,Waldor MK.Emerging knowledge of regulatory roles of D-amino acids in bacteria.
2011;68:817-831[PMID:21161322 DOI:10.1007/s00018-010-0571-8]
87 Radkov AD,Moe LA.Bacterial synthesis of D-amino acids.
2014;98:5363-5374[PMID:24752840 DOI:10.1007/s00253-014-5726-3]
88 Gilmore MS,Skaugen M,Nes I.Enterococcus faecalis cytolysin and lactocin S of Lactobacillus sake.
1996;69:129-138[PMID:8775973 DOI:10.1007/BF00399418]
89 Lam H,Oh DC,Cava F,Takacs CN,Clardy J,de Pedro MA,Waldor MK.D-amino acids govern stationary phase cell wall remodeling in bacteria.
2009;325:1552-1555[PMID:19762646 DOI:10.1126/science.1178123]
90 Ketting D,Wadman SK,Spaapen LJ,Van der Meer SB,Duran M.Gas chromatography method for the separation of amino acids enantiomers in plasma and urine.Application in a case of short bowel syndrome.
1991;204:79-86[PMID:1819475 DOI:10.1016/0009-8981(91)90219-3]
91 Chai Z,Wu T,Dai A,Huynh P,Koentgen F,Krippner G,Ren S,Cooper ME.Targeting the CDA1/CDA1BP1 Axis Retards Renal Fibrosis in Experimental Diabetic Nephropathy.
2019;68:395-408[PMID:30425061 DOI:10.2337/db18-0712]
92 Vaziri ND,Wong J,Pahl M,Piceno YM,Yuan J,DeSantis TZ,Ni Z,Nguyen TH,Andersen GL.Chronic kidney disease alters intestinal microbial flora.
2013;83:308-315[PMID:22992469 DOI:10.1038/ki.2012.345]
93 Wang H,Lu Y,Yan Y,Tian S,Zheng D,Leng D,Wang C,Jiao J,Wang Z,Bai Y.Promising Treatment for Type 2 Diabetes:Fecal Microbiota Transplantation Reverses Insulin Resistance and Impaired Islets.
2019;9:455[PMID:32010641 DOI:10.3389/fcimb.2019.00455]
94 Ganesan K,Chung SK,Vanamala J,Xu B.Causal Relationship between Diet-Induced Gut Microbiota Changes and Diabetes:A Novel Strategy to Transplant Faecalibacterium prausnitzii in Preventing Diabetes.
2018;19[PMID:30467295 DOI:10.3390/ijms19123720]
95 Xu J,Liang R,Zhang W,Tian K,Li J,Chen X,Yu T,Chen Q.Faecalibacterium prausnitzii-derived microbial antiinflammatory molecule regulates intestinal integrity in diabetes mellitus mice
modulating tight junction protein expression.
2020;12:224-236[PMID:31503404 DOI:10.1111/1753-0407.12986]
96 Björkqvist O,Rangel I,Serrander L,Magnusson C,Halfvarson J,Norén T,Bergman-Jungeström M.Faecalibacterium prausnitzii increases following fecal microbiota transplantation in recurrent Clostridioides difficile infection.
2021;16:e0249861[PMID:33836037 DOI:10.1371/journal.pone.0249861]
97 Chen HT,Huang HL,Xu HM,Luo QL,He J,Li YQ,Zhou YL,Nie YQ,Zhou YJ.Fecal microbiota transplantation ameliorates active ulcerative colitis.
2020;19:2650-2660[PMID:32256746 DOI:10.3892/etm.2020.8512]
98 Li Y,Su X,Gao Y,Lv C,Gao Z,Liu Y,Wang Y,Li S,Wang Z.The potential role of the gut microbiota in modulating renal function in experimental diabetic nephropathy murine models established in same environment.
2020;1866:165764[PMID:32169506 DOI:10.1016/j.bbadis.2020.165764]
99 Hu ZB,Lu J,Chen PP,Lu CC,Zhang JX,Li XQ,Yuan BY,Huang SJ,Ruan XZ,Liu BC,Ma KL.Dysbiosis of intestinal microbiota mediates tubulointerstitial injury in diabetic nephropathy
the disruption of cholesterol homeostasis.
2020;10:2803-2816[PMID:32194836 DOI:10.7150/thno.40571]
100 Lu J,Chen PP,Zhang JX,Li XQ,Wang GH,Yuan BY,Huang SJ,Liu XQ,Jiang TT,Wang MY,Liu WT,Ruan XZ,Liu BC,Ma KL.GPR43 deficiency protects against podocyte insulin resistance in diabetic nephropathy through the restoration of AMPKα activity.
2021;11:4728-4742[PMID:33754024 DOI:10.7150/thno.56598]
杂志排行

World Journal of Diabetes的其它文章
- Beyond diabetes remission a step further:Post bariatric surgery hypoglycemia
- Free fatty acids,glucose,and insulin in type 2 diabetes mellitus
- Sodium-glucose co-transporter 2 inhibitors induced euglycemic diabetic ketoacidosis within four days of initiation
- Age at diagnosis of type 2 diabetes and cardiovascular risk factor profile:A pooled analysis
- Hemoglobin within normal range is negatively related to hemoglobin A1c in a nondiabetic American population aged 16 years and older
- Functional annotation and enrichment analysis of differentially expressed serum proteins in patients with type 2 diabetes after dapagliflozin