外源钙对低温胁迫下梨生理特性的影响
2022-03-16武建强张文菲张熠可韩爱谦袁嘉玮
潘 琪,武建强,贾 慧,宁 芳,张文菲,张熠可,韩爱谦,袁嘉玮
(1.太原市农业技术推广服务中心,山西 太原 030027;2.山西农业大学 棉花研究所,山西 运城 044000)
我国是梨属植物的主要原产地,梨树种植面积及产量均位居世界第一。目前,我国北方梨树种植区饱受春季花期冻害的困扰,当梨盛花期温度低于-2.0 ℃时以及梨始花期温度低于-3.9 ℃,将会发生明显冻害[1]。因此,短时间内冷空气的突袭会使梨树花器官、叶片遭受冻害,导致果树减产甚至绝产,损失严重。目前,花期冻害成为限制梨产业发展的主要因素之一[2]。
丙二醛(MAD)含量是评价植物膜系统在非生物胁迫下受损程度的重要指标[3]。超氧化物歧化酶(SOD)是抵抗活性氧的第一道门槛,其作用主要是清除引起细胞结构和功能破坏的生物体内特异超氧阴离子自由基,SOD水平与生物体的衰老和死亡直接相关。过氧化氢酶(CAT)与过氧化物酶(POD)可将逆境胁迫下细胞内产生的过氧化氢和超氧阴离子自由基转换成水和氧气,从而减少逆境对膜系统的破坏,通常情况下抗逆性强的品种在逆境条件下CAT 和POD 的活性较强[4]。光合作用是植物获取能量及合成有机物的重要生理生化过程,净光合速率(Pn)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)、气孔导度(Gs)可反映逆境对植物光合作用的影响[5-6]。因此,利用MAD、SOD、POD、CAT 等植物抗逆性指标和 Pn、Ci、Tr、Gs 等植物光合作用的生理指标,结合隶属函数分析进行综合评价,可反映不同钙处理下梨花器官及叶片的抗寒性差异[7]。
钙在调控植物逆境生理及生长发育上发挥着重要作用[8]。植物体在低温胁迫条件下通过激活钙通道调控钙库的释放,以提高Ca2+水平来诱导抗冷相关基因的表达,从而提高植物体的抗寒性[9]。MONROY 等[10]研究表明,通过化学处理阻断Ca2+通道,拮抗钙调素作用或是抑制蛋白激酶,可使植株的抗寒性显著降低。在低温胁迫下施加一定浓度Ca2+可显著提高植株抗寒性,目前在降香黄檀(Dalbergia odoriferaT. Chen)[11]、尾巨桉(Eucalyptus urophylla×E. grandis)[12]、豆梨(Pyrus calleryana)[13]等中都有一定研究。有关梨的研究,张超等[14]研究发现,相同低温处理时间对不同梨品种花器官损伤不同;肖坤等[15]研究发现,叶面喷施15 mmol/LCaCl2可提高枝条中的脯氨酸和可溶性糖含量以提升植物抗寒性;周君等[16-17]研究发现,叶面喷施不同钙肥可提升梨叶片光合生理特性。但针对通过外源施加Ca2+能否降低花器官的冻害率以及不同浓度、不同种类的Ca2+对提高梨树抗寒性是否有差异尚未见报道。
本研究以山西省主栽梨树品种玉露香梨为试材,分析施用外源钙对低温胁迫处理下梨花器官、叶片抗寒生理及光合生理的影响,以期提高梨抗寒性,为梨逆境耐受性研究及品质改良和高产栽培研究提供理论依据。
1 材料和方法
1.1 试验材料
供试玉露香梨采集于运城市盐湖区泓芝驿镇,选择长势整齐一致、生长健壮、无病虫害的15年生树 30 株,于 2020 年 3 月 20 日从树冠 4 个方向采集1 年生 10 cm 花枝,于 2020 年 4 月 20 日从树冠外围采集长势一致的生长枝。
1.2 试验方法
供试材料采集后立即放入人工气候室进行外源钙喷施处理,共设置CK(不喷钙)、T1(14 mmol/L CaCl2)、T2(21 mmol/L CaCl2)、T3(28 mmol/L CaCl2)、T4(14 mmol/L Ca(NO3)2)、T5(21 mmol/L Ca(NO3)2)、T6(28 mmol/L Ca(NO3)2)7 个处理。各处理均为每2 d喷施一次,喷至花器官、叶片完全湿润,培养6 d后进行低温胁迫处理,-2 ℃条件下胁迫3 h。采集低温处理后的花器官、叶片待测。
1.3 测定指标及方法
MDA 含量和SOD、POD、CAT 活性测定参考史树德等[18]的方法进行;Pn、Ci、Tr、Gs 等光合指标采用英国ADC 公司生产的LCpro T 光合仪进行测定。
1.4 数据统计
隶属函数分析参考文献[19-20]的方法进行;利用Office 2019和SAS 8.2软件进行数据分析。
2 结果与分析
2.1 生理指标统计分析
从表1、2可以看出,不同处理的变异系数存在着较大差异,其中,花器官SOD活性、POD活性及叶片SOD 活性、POD 活性、气孔导度的变异系数比较大 ,分 别 是 19.48%、29.78%、16.67%、22.59%、26.10%,属于中等强度变异,其余指标的变异系数比较低。表明SOD 活性、POD 活性、气孔导度这3 个性状指标数据存在较大差异,受低温胁迫处理的影响明显。
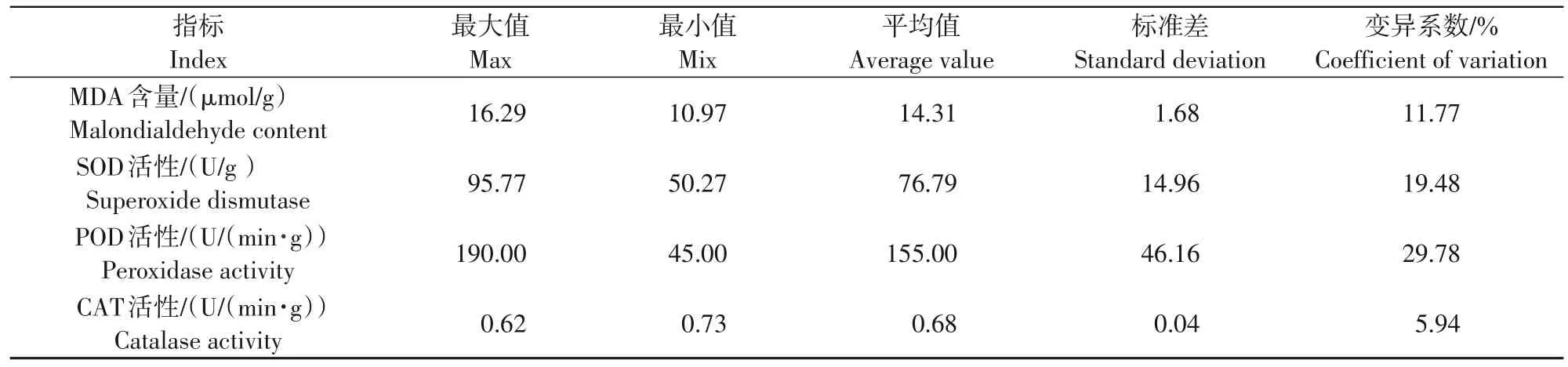
表1 低温处理下梨花器官抗寒生理指标统计分析Tab.1 Statistical analysis of cold-resistant physiological indexes of pear flower under low temperature treatment

表2 低温处理下梨叶片生理指标统计分析Tab.2 Statistical analysis of physiological indexes of pear leaves under low temperature treatment
2.2 不同外源钙对低温胁迫下梨花器官抗寒生理指标的影响
由表3可知,不同外源钙各浓度处理均降低了低温胁迫下梨花器官的MDA 含量,提高了POD 活性及 CAT 活性。MDA 含量除 T1、T3 处理外,其他处理均与CK 间呈显著性差异(P<0.05),其中,T2处理较 CK 降低了9.9%,硝酸钙T4、T5、T6 处理与CK 相比MDA 含量分别降低了14.9%、32.7%、18.8%;氯化钙与硝酸钙各浓度处理均在21 mmol/L处理时最低,其中T5 处理达最低。除T4、T6 处理外,各外源钙处理与CK 相比均显著提升了SOD 活性(P<0.05),其中氯化钙T1、T2、T3处理分别提高了13.4%、24.7%、38.0%,T5处理提高了41.1%;随氯化钙各处理随浓度升高,SOD 活性相应升高,表明施用CaCl2提高梨花器官SOD活性的最适浓度不在设置的浓度梯度范围内。各外源钙处理与CK相比均显著提升了POD 活性(P<0.05),其中氯化钙T1、T2、T3 处理分别是CK 的3.44、3.94、4.00 倍,硝酸钙T4、T5、T6 处理分别是 CK 的 3.89、4.22、3.61 倍;与SOD 活性结果类似,CaCl2处理的POD 活性未达到峰值,其最适浓度不在设置的浓度梯度范围内,T5处理POD 活性最高,但与T2、T3、T4 处理间无显著性差异。除T1、T4处理CAT活性与CK间无显著差异外,其余处理与CK 相比均存在显著性差异(P<0.05),其中,氯化钙T1、T2、T3 处理CAT 活性较CK分别提高1.6%、14.5%、11.3%,硝酸钙T4、T5、T6 处理CAT 活性较CK 分别提高4.8%、17.7%、14.5%。CaCl2、Ca(NO3)2处理 CAT 活性均在 21 mmol/L 时最高,其中21 mmol/L Ca(NO3)2处理达最高。
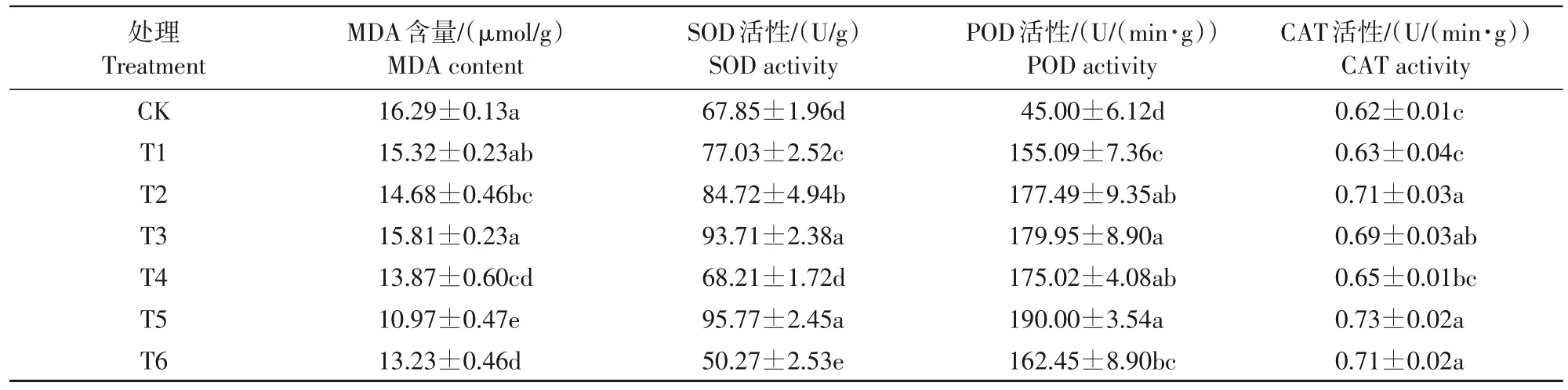
表3 不同外源钙对低温胁迫下梨花器官抗寒生理指标的影响Tab.3 Effects of different exogenous calcium on cold-resistant physiological indexes of pear flower under low temperature stress
2.3 不同外源钙对低温胁迫下梨叶片抗寒生理指标的影响
由表4可知,不同外源钙各浓度处理均降低了低温胁迫下梨叶片的MDA含量,提高了SOD活性、POD活性及CAT活性。MDA含量除T3处理外其余处理均与CK间呈显著性差异(P<0.05),其中硝酸钙各浓度处理整体表现优于氯化钙处理,硝酸钙T4、T5、T6 处理较 CK 分别降低了 20.8%、25.4%、20.0%。各外源钙处理SOD活性均显著高于CK(P<0.05),其中T2 处理显著高于其余处理(P<0.05),较CK 提高了59.5%;T1 处理显著高于除T2 处理外的其余处理(P<0.05),较CK提高了48.8%;硝酸钙各浓度处理间无显著性差异,T4、T5、T6 处理较CK分别提高了22.2%、25.5%、25.2%。与SOD相似,各外源钙处理POD活性均显著高于CK,其中T2处理显著高于其他处理,较CK 提高了122.0%。硝酸钙各浓度处理间无显著性差异,T4、T5、T6 处理较CK分别提高了66.5%、78.9%、76.8%。除T4 处理外其余处理CAT 活性均显著高于CK(P<0.05),T2处理显著高于除T3 处理外的其余处理(P<0.05),T2、T3处理CAT活性较CK分别提高46.1%、34.8%;硝酸钙各浓度处理间无显著性差异,T4、T5、T6 处理较CK分别提高了13.5%、23.6%、21.3%。
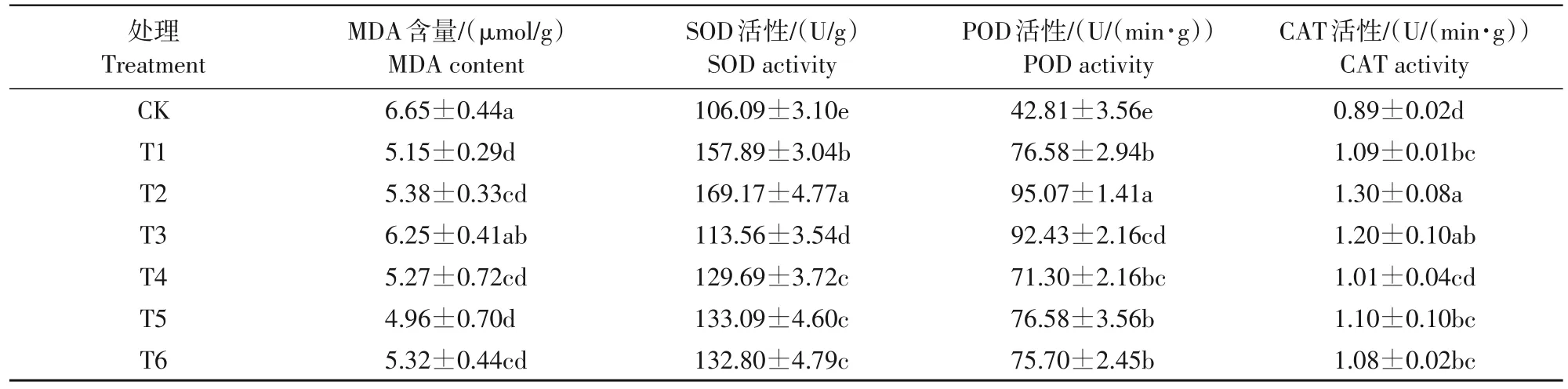
表4 不同外源钙对低温胁迫下梨叶片抗寒生理指标的影响Tab.4 Effects of different exogenous calcium on cold-resistant physiological indexes of pear leaves under low temperature stress
2.4 不同外源钙对低温胁迫下梨叶片光合生理指标的影响
从表5可以看出,不同外源钙各浓度处理均提高了低温胁迫下梨叶片的净光合速率和蒸腾速率,且不同外源钙的净光合速率、蒸腾速率、胞间二氧化碳浓度3 个光合生理指标值均随浓度的增加而升高,表明施用外源钙处理提高梨叶片净光合速率、蒸腾速率、胞间二氧化碳浓度3 个光合生理指标值的最适浓度不在设置的浓度梯度范围内。除T4、T5 处理外其余外源钙处理净光合速率均显著高于CK(P<0.05),其中,T3处理显著高于除T6处理外的其余处理(P<0.05),T3、T6 处理分别较CK提升了48.8%、39.0%。与净光合速率相似,除T4、T5处理外其余外源钙处理蒸腾速率均显著高于CK(P<0.05),其中,T3 处理蒸腾速率最高,T6 处理蒸腾速率次之,二者分别较CK提升了50.0%、37.1%。胞间二氧化碳浓度除T3 处理显著高于CK、T1、T4处理外(P<0.05),其余处理间无显著性差异,T3处理胞间二氧化碳浓度较CK 提高了18.7%。T2、T3处理气孔导度显著高于除T6 处理外的其余处理(P<0.05),T2、T3、T6 处理气孔导度分别较CK 提高了71.4%、71.4%、42.9%。

表5 不同外源钙对低温胁迫下梨叶片光合生理指标的影响Tab.5 Effects of different exogenous calcium on photosynthetic physiological indexes of pear leaves under low temperature stress
2.5 叶片抗寒生理与光合生理相关性分析
如表6所示,在对低温胁迫下各外源钙浓度处理的梨叶片抗寒生理指标和光合生理指标进行相关性分析发现,POD活性、CAT活性、Gs互呈显著正相关关系(P<0.05),其中POD 活性与CAT 活性呈极显著正相关关系(P<0.01);Pn、Tr、Ci 互呈显著正相关关系(P<0.05),其中Pn与Tr呈极显著正相关关系(P<0.01);POD活性、Pn、Gs、Tr互呈显著正相关关系(P<0.05)。表明外源钙可以通过调控光合性能促进植株抗寒能力的提升。
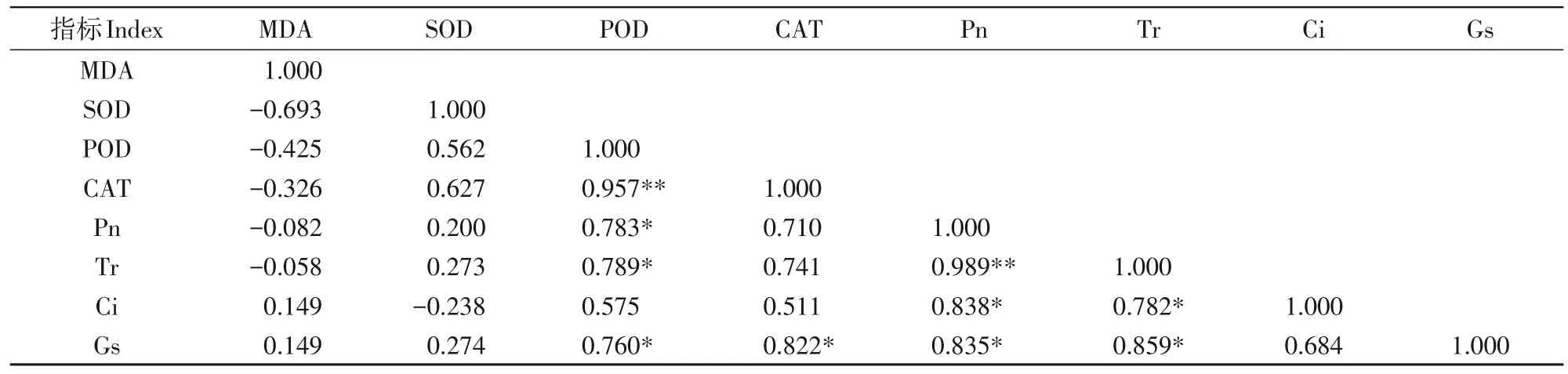
表6 叶片抗寒生理与光合生理相关性分析Tab.6 Correlation analysis of leaf cold-resistant physiology and photosynthetic physiology
2.6 隶属函数分析
鉴于不同指标测定的最适宜浓度并非完全相同,因此利用平均隶属函数分析进行梨花器官4个抗寒性指标的综合评价,结果表明(表7),不同处理梨花器官抗寒性强弱表现为21 mmol/LCa(NO3)2>21 mmol/L CaCl2>28 mmol/L CaCl2>28 mmol/L Ca(NO3)2>14 mmol/L Ca(NO3)2>14 mmol/L CaCl2;利用平均隶属函数分析进行梨叶片的抗寒及光合指标进行综合评价,结果表明(表8),不同处理梨叶片生理特性强弱表现为21 mmol/LCaCl2>28 mmol/L CaCl2>28 mmol/L Ca(NO3)2>14 mmol/L CaCl2>21 mmol/LCa(NO3)2>14 mmol/L Ca(NO3)2。 由 此可知,不同浓度下的 CaCl2、Ca(NO3)2处理均可提高梨花器官的抗寒性及叶片的生理特性,其中,在21 mmol/L Ca(NO3)2处理后梨花器官抗寒性最强,在21 mmol/L CaCl2处理后梨叶片抗寒性及光合性能最强。

表7 低温胁迫下梨花器官抗寒性综合评价Tab.7 Comprehensive evaluation of cold resistance of pear flower organs under low temperature stress
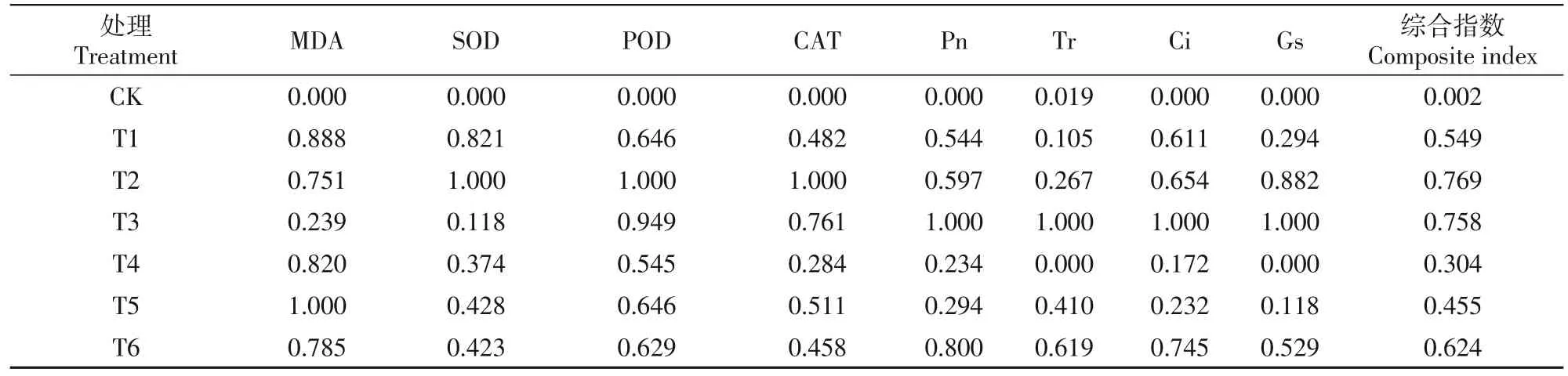
表8 低温胁迫下梨叶片抗寒性综合评价Tab.8 Comprehensive evaluation of cold resistance of pear leaves under low temperature stress
3 结论与讨论
MDA 是植物细胞质膜受到损伤后的过氧化物产物,因此其可作为反映植物受损程度的量化指标[21-22]。本研究表明,不同浓度的 CaCl2、Ca(NO3)2处理均可不同程度降低低温胁迫下梨花、叶片MDA 的含量,说明外源钙处理可降低低温胁迫下梨花、叶的损伤程度。同时,研究表明,过高的Ca2+也会降低其对于植物损伤的缓解效果,这与王小媚等[23]的研究结果一致。
POD、SOD、CAT 均是评价植物体衰老、萎蔫程度的重要生理指标[24]。本研究中,CaCl2处理的SOD活性、POD 活性仅呈现上升趋势,表明其峰值不在本研究浓度梯度设置范围内。但由于在该浓度范围下,CaCl2处理 MDA 含量、CAT 活性均呈现峰值,且Ca(NO3)2处理各指标也均遵循该趋势规律,因此本研究浓度梯度范围设置合理。然而,本研究发现,不同指标的测定结果中最适宜浓度并不相同,说明不能依据单一指标判断不同外源钙各浓度处理对于梨花器官的抗寒性能,这与周福平等[25]针对低温胁迫下高粱幼苗生理特性变化的研究结论相一致。Pn、Ci、Tr、Gs是评价植物光合作用能力的重要生理指标[26]。本研究表明,适宜的外源钙种类及浓度可显著提升低温胁迫下梨叶片的光合作用能力,并通过调控光合性能促进植株抗寒能力的提升。宋俏博[27]研究认为,外源Ca2+可以通过有效激发植物生长发育和光化学活性,维持叶片中非结构型碳水化合物有效外运,降低碳水化合物和ROS 过度积累,从而缓解低温下植株的光抑制现象,与本研究结果一致。本研究通过隶属函数分析进行抗寒性指标及光合指标的综合评价,结果表明,21 mmol/LCa(NO3)2处理后梨花器官抗寒性最强,21 mmol/L CaCl2处理后梨叶片抗寒性及光合性能最强。本研究中,相同浓度的CaCl2、Ca(NO3)2处理对指标的影响存在差异,而其本身Ca2+浓度是相同的,其原因可能是由于氯离子相较硝酸根离子与钙离子结合更紧密,有助于提升植株细胞吸收钙的效率,提高抗逆性,其机制还有待进一步研究。
本研究结果表明,在响应低温胁迫的抗寒生理指标和光合生理指标中,SOD、POD、Gs生理指标更易进行人为调控。相关性分析发现,POD 活性、CAT 活性、Gs 互呈显著正相关关系,Pn、Tr、Ci 互呈显著正相关关系,POD 活性、Pn、Gs、Tr 互呈显著正相关关系,表明外源钙可通过调控光合性能促进植株抗寒能力的提升。在梨花器官、叶片上喷施适宜外源钙浓度可显著抑制MDA 的生成,提升SOD、POD、CAT 等抗氧化相关酶的活性,提高梨叶片净光合速率、蒸腾速率、气孔导度等光合性能指标,进而提升梨产量及品质。研究结果表明,Ca(NO3)2为梨树花期喷施钙肥的最适种类,21 mmol/L 为Ca(NO3)2最适浓度;CaCl2为梨树果实膨大期喷施钙肥的最适种类,21 mmol/L为CaCl2最适浓度。
