农作物籽粒分割研究现状综述
2022-03-11彭顺正岳延滨冯恩英黄安林
彭顺正 岳延滨 冯恩英 黄安林

摘要 籽粒分割是籽粒计数的关键环节,也是种子精选、育种评价的重要研究内容之一。从机器视觉和深度学习角度分析籽粒和颗粒分割现状,总结分析了各种分割方法特点。并对视觉方法和深度学习方法在图像分割等方面进行了比对分析,体现了深度学习在分割方面的优越性。
关键词 籽粒;图像分割;机器视觉;深度学习
中图分类号 TP 391.4文献标识码 A
文章编号 0517-6611(2022)04-0013-04
doi:10.3969/j.issn.0517-6611.2022.04.004
开放科学(资源服务)标识码(OSID):
A Review of the Research Status of Crop Grain Segmentation
PENG Shun-zheng, YUE Yan-bin, FENG En-ying et al (Institute of Science and Technology Information, Guizhou Academy of Agricultural Sciences, Guiyang, Guizhou 550006)
Abstract Grain segmentation is a key part of grain counting, and it is also one of the important research contents of seed selection and breeding evaluation. From the perspective of machine vision and deep learning, the current situation of grain and granule segmentation is analyzed, and the characteristics of various segmentation methods are summarized and analyzed. And compared and analyzed the visual methods and deep learning methods in image segmentation and other aspects, which reflects the superiority of deep learning in segmentation.
Key words Grain;Image segmentation;Machine vision;Deep learning
基金项目 贵州省2020年科技支撑计划项目(黔科合支撑〔2020〕1Y019)。
作者简介 彭顺正(1989—) ,男,贵州兴义人,助理研究员,硕士,从事农产品品质检测、图像算法研究。
收稿日期 2021-05-20
农业种子工程战略的实施是支撑我国农业产业发展的一项基本举措。育种创新、种子精选等是农业工程的重要内容。种子精选和育种评价需要对籽粒性状进行评价,千粒重就是一种重要的农艺性状参数[1-2]。基于图像方法检测千粒重等参数,其原理是对籽粒进行分割计数,然后检测重量。籽粒分割好坏直接关系到籽粒计数是否准确。研究籽粒分割方法对于籽粒农艺性状参数检测具有重要意义。因此,笔者基于机器视觉和深度学习的角度,概述籽粒分割研究现状。由于颗粒分割和籽粒分割具有同质性,也对颗粒的分割研究进行分析总结,并展望农作物籽粒分割的发展方向。
1 基于机器视觉的传统分割研究现状
基于机器视觉的方法分割作物种子籽粒应用比较广泛、涉及的类型比较多,不同作物籽粒性状、外观色泽、空间立体姿态不同,导致空间分布排列存在差异。这些客观性因素导致目前没有一种通用的作物籽粒分割算法。一般情况下,只讨论籽粒在空间分布上是单层的情况,重叠分布的情况不作讨论,这种情况下视觉分割方法适应性差,无法对遮挡部分进行准确的分割。籽粒空间分布主要存在2种分布:单籽粒与粘连籽粒。一般来说,单籽粒的分割比较简单,不同分割方法都可以获取比较满意的分割效果。然而对于粘连籽粒的分割方法就相对复杂很多,主要是粘连籽粒经过图像预处理之后,其边缘形态、颜色均会发生一定程度的变化。对于籽粒轮廓边缘识别和黏结位置的定位分割是籽粒分割关键技术难点,因此衍生了各种分割算法。从分割原理角度来看,总体上分为:基于颜色的分割、基于形状的分割,基于颜色+几何形状的分割3种类型[3-5]。
适当地增强目标和背景对比度可以提高籽粒分割的准确度。例如,在RGB、Lab、HSV等颜色空间模型中,利用分量图和灰度图特征比较或重叠,以及各通道颜色之间的变换或差分等运算以增强目标和背景的对比度为分割打下了基础[3-5]。
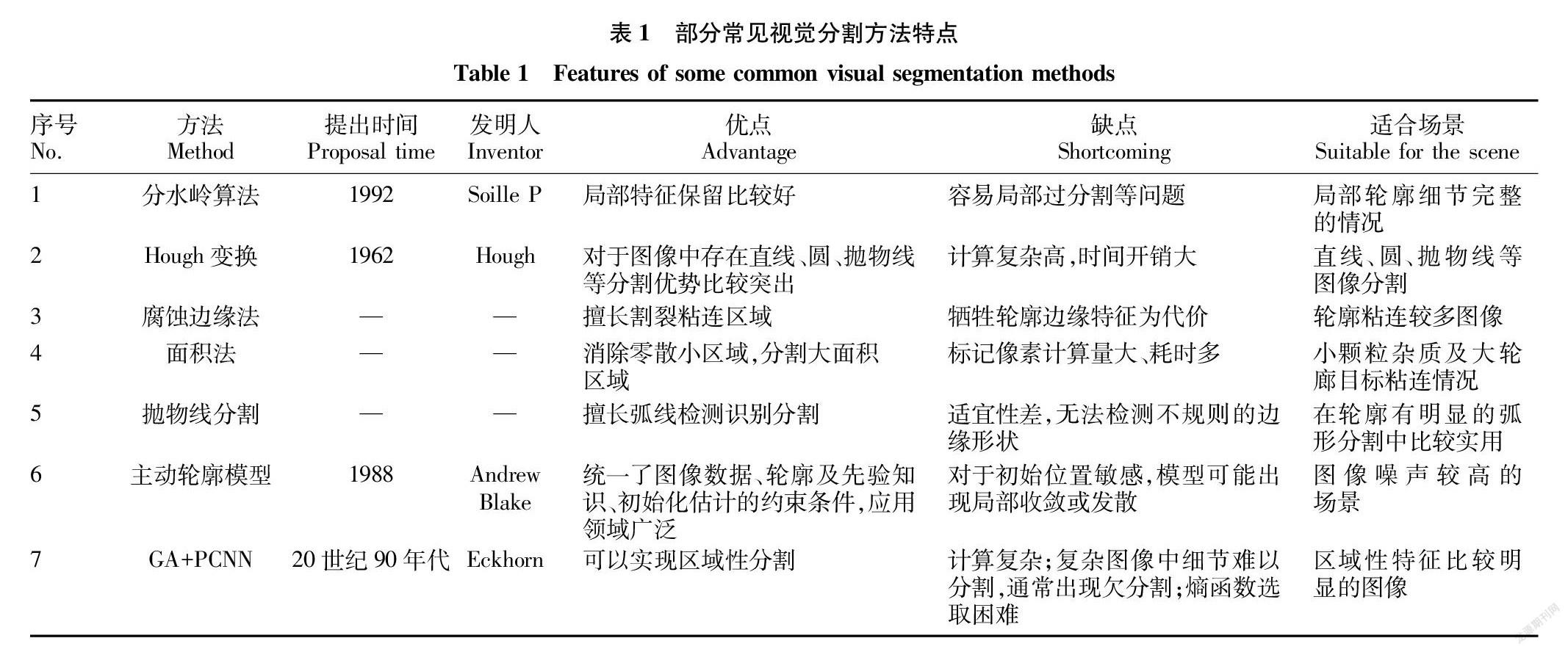
籽粒分割就是将其边缘轮廓准确的分割成独立的整体。目前轮廓提取方面常见的方法主要有分水岭算法(包括改进的分水岭算法)、Hough变换方法、腐蚀边缘法、面积法、抛物线分割、主动轮廓模型、遗传算法(GA)与改进脉冲耦合神经网络(PCNN)相结合等分割方法[6-24]。
由表1可知,①分水岭算法依赖于局部像数值的变化,判断局部极值所在位置,进而实现目标轮廓的方法。然而,根据相关研究经验,分水岭算法在部分情况下容易出现过分割现象,因此有学者也对分水岭算法进行了多种尝试和改进[6-14]。②Hough变换方法主要是检测图像中直线或曲线轮廓,从而达到轮廓识别定位分割的目的[15]。③腐蚀边缘法原理是通过形态学腐蚀方法剥离掉连通区域的重叠部分,从而实现黏结部分的分割[16-18]。④面积法通常采用标记像素来实现目标区域分割[19-20],为了减少对目标像数遍历和计算量,部分学者提出采用“跳跃”扫描方式,而后根据邻域像数单元检测填充断档区域实现目标区域分割[21]。⑤抛物线分割方法[22]事实上比较倾向于依赖先验知识。比如谷物籽粒类似椭圆、油菜籽粒类似圆,这些籽粒的形状都具有比较明显的空间几何特征,类似于先验知识。抛物线分割原理主要是检测定位3个关键点来拟合抛物线作为模拟轮廓。事实上,抛物线分割优缺点也比较明显,对于弧形籽粒抛物线分割比较好,但无法对残缺籽粒进行完整识别分割。⑥主动轮廓模型[23]分割通过寻找轮廓最尖锐位置,计算弧线曲率大小和方向判断籽粒是否粘连,从而定位分割点。但对于轮廓局部尖锐层度变化比較敏感,容易受到轮廓毛刺干扰,通常需要对轮廓平滑处理。⑦遗传算法(GA)与改进脉冲耦合神经网络(PCNN)相结合[24]的分割方法原理是:采用PCNN网络先激活图像中所有像素,产生迭代阈值,不断循环,直到存在像数值小于迭代阈值时,产生抑制脉冲,引起邻近类似像数神经元抑制,获取一系列脉冲序列。其中第N次离散时间所得序列作为目标图像,而后通过GA算法优化PCNN模型参数进行图像分割。
综上所述,增强图像的对比度可以提高籽粒分割的准确性;综合考虑籽粒颜色、空间几何形状等特征有助于分割方法的选取。目前还没有通用的分割方法适合任何籽粒的分割。因此,分割方法选取是籽粒分割的一个关键环节。深入了解各种分割方法的特点和优缺点有助于选取合适的分割方式,对于提高籽粒分割效率和准确性具有重要意义。
2 基于深度学习的主流分割研究现状
深度学习是一种含有多层感知神经网络结构[25]。目前,基于深度学习的籽粒图像分割相对较少,国内相关研究主要集中在检测方向[26-38],还有关于预测还原残缺籽粒的相关报道[39]。由于具有分割速度快、精度高、基于像数级定位分类分割的特点,深度学习在图像分割方面得到广泛运用。
目前,国内相关学者基于深度学习方法分割作物籽粒。尤其以谷物、小麦、大豆等最为常见。深度卷积结构模型是影响分割性能主要因素之一,网络结构的选择和设计非常关键。有学者采用Mask R-CNN网络和FCN网络对麦穗图像进行分割,发现Mask R-CNN网络处理速度比FCN网络快8.5倍[26];在类似大豆籽粒分类研究中,也证实网络结构选择和设计直接影响网络模型的分割准确率和实时性[27]。深度学习分割本质其实就是对像数级图像特征进行分类,从而实现目标分割识别。如采用U-Net网络分割水稻籽粒图像,就是通过训练各类别的像数特征实现分割[28]。某些情况下,综合考虑图像分辨率、籽粒各部位尺寸悬殊对语义分割网络设计具有重要的作用。例如,针对稻米垩白识别的G-Chalk网络改进和分割识别准确度提高就说明了其重要性[29]。
在籽粒检测分类识别方面,目前涉及玉米、小麦、大米、花生等。对网络卷积核、各层特征的可视化分析,有助于构建实例的分类识别体系。例如,基于VGG16网络对玉米品种分类识别[30]。基于Faster R-CNN模型实现玉米品质精选的研究[31]。这些研究说明,可以采用多个检测模型对同一问题开展研究,这有助于构建最优的检测模型。向日葵种子识别就采用多个检测模型进行试验比对,以确立最优检测识别模型[32]。目前的检测模型结构由2个主体部分构成,经典网络模型(如AlexNet、GoogleNet、VGG等)及特殊功能的块结构(如残差块、Dropout技术、BN算法等)模式。小麦识籽粒完整性检测识别网络ResNet模型结构就是典型代表[32-33]。事实上,影响检测模型检测识别精度除网络结构本身外,还存在其他因素。比如,自然图像数据维度高,常常影响图像识别的推理速度,针对这个问题,可采用种子图像降低数据维度,在网络卷积层加入注意力机制模块,以提高模型特征描述性能,进而提高目标识别精度[34]。另外,为弥补卷积网络模型特征利用率低下的不足,有研究结合SVM提高特征复用性,改善检测模型的识别精度[34-35]。深度学习训练需要大量数据样本,有利于模型训练,为了减少作物籽粒图像采集量,一般采用数据增广(如翻转、旋转、对比度调整、图像拉伸、不同尺度的剪切等操作)扩充数据样本量。模型训练比较耗时,收敛速度慢是深度学习目前面临比较多的问题,目前,有学者采取激活函数优化和池化等操作提高有效特征提取效率或数据降维以缩短模型训练周期[36]。还有学者通过调整学习参数提高模型收敛速度,如花生颗粒分类的CNN+SqueezeNet模型[37]。总之,可以结合数据集构成、过拟合弱化、训练收敛速度、网络結构简化等方面优化检测模型来提高籽粒识别分类[38]。
当前,有学者还尝试了采用深度学习对残缺籽粒或遮挡部分进行复原,并取得比较理想的效果[39]。各卷积网络的特点见表2。
综上所述,深度学习在籽粒分割、检测等方面,其模型选择设计除了考虑籽粒形状、图像分辨率以外,还需要考虑网络结构本身。无论是分割还是检测,不同的网络结构对不同种子籽粒分割和检测都具有一定的适应性。因此,一般需要试验多种不同的网络结构模型,确立最优模型。在模型训练和性能提升方面,可以结合数据集、卷积层、学习参数、激活函数、优化器、池化等方面优化网络,缩短模型收敛时间、提高准确度。目前,对花生、大豆等籽粒的研究可以为其他作物籽粒分割检测方面提供借鉴。
3 非农作物颗粒分割研究现状
考虑到籽粒分割与其他非作物颗粒分割具有一定的同质性。因此,其他非作物颗粒物分割对于作物籽粒分割具有一定的参考意义。
3.1 机器视觉分割研究方面 目前主要包括岩石、矿石、煤、金刚石、炉渣、雨滴、细胞等颗粒状物体分割。针对颗粒粘连分割采取的方法包括距离变换+分水岭+形态学、BP神经网络+归一化算法、大津法+形态学+分水岭、图像邻域的密度统计、中值滤波和小波变换+分割算法、凸包分析的图像二次分割算法、基于直方图累积矩的动态优化算法、大津法+形态学+连通标记、大津法双阈值算法、灰度直方图配准+双峰直方图、链码模型等[40-54]。其中分水岭、BP神经网络、形态学、大津法在作物籽粒中也有应用,说明籽粒和颗粒分割比较类似,其余算法在籽粒分割中应用较少,说明颗粒和籽粒分割有其各自的特殊性。一般来说,同种籽粒形状比较类似,规律性较强。而颗粒形状、大小是随机变量,没有多少可行。但空间排布上,二者无非是单粒和粘连两种常见类型(重叠类型除外)。
3.2 深度学习分割方面 目前涉及矿物微粒、未溶碳化物颗粒、化学混合物粒子、烟雾颗粒、纳米颗粒、岩石颗粒、混合颗粒等方面,分割模型主要包括MASK R-CNN、U-Net、RPN、CNN、FCNN。针对颗粒分割,这些分割模型重点学习颗粒轮廓像数特征,提升颗粒定位精度实现颗粒部分像数级分割[55-62],这些分割模型在作物籽粒中也有运用,说明深度学习网络模型具有很好的通用性。从分割对象来看,深度学习分割已经涉及微粒子、化合物等更加微小的颗粒,足以说明其分割精度和优越性不言而喻,未来应用的前景会更加广阔,也将是未来微粒图像分割发展的风向标。
4 小结
该研究论述了图像分割对作物籽粒的重要性,系统总结和分析了图像在颗粒状分割方面的研究现状,并对视觉方法和深度学习2个领域中常见且具有代表性的算法进行了分析总结,明确了各算法分割机理及其优缺点,揭示了各算法的优缺点和适用场景。另外还对颗粒和籽粒的分割方法进行了分析比对,研究表明:无论是视觉方法还是深度学习方法,二者互有重叠,说明二者分割机制具有互通性。
机器视觉分割方法较多依赖于籽粒/颗粒的形状、颜色、纹理等特征。分割方法众多,但通用性差,且无法达到像素级分割,对于微型颗粒识别分割精细程度较弱,这是导致视觉方法在微粒分割方面应用比较少的主要原因。
深度学习分割和检测机理是对每个像素特征进行分类识别,从而达到检测识别分割的目的。基于像素级的分割,分割结果的精准度和精细化程度高,是其在化合物粒子、微粒子等肉眼无法识别的情况下,分割识别应用比较突出主要原因。深度学习通过提取像素特征,将像素特征输入到分类网络分类识别,并将每个像素归类到特定的类别集合中,最终将不同类别的像素归类到不同的集合,从而达到识别分割效果。
从分割对象角度来看,深度学习应用范围比视觉方法应用范围广阔,从块状到微粒子均有涉及,且图像数据处理量大、检测速度快、精度高、理论技术新颖,是目前图像分割技术发展的一座里程碑,应用前景广阔。
参考文献
[1]
杨国浪,张晓强,徐长虹,等.灰色关联度分析在西藏河谷农区甘蓝型油菜育种中的应用[J].种子,2020,39(2):113-117.
[2] 费锦宗,沈金雄,郭彦丽,等.中国油菜区试新品种产量和品质性状分析[J].湖北农业科学,2019,58(22):33-40.
[3] 王金星,李玉风,江浩,等.玉米籽粒内部组分低对比度透射图像精确分割研究[J].农业机械学报,2020,51(6):238-247.
[4] 燕红文,崔清亮.基于分水岭分割算法对燕麦籽粒识别中粘连区域的分割[J].计算机时代,2020(1):23-25.
[5] 杨涛,肖衡,杨博雄,等.基于图像的玉米籽粒粘连的分割方法研究[J].科學技术创新,2019(36):87-89.
[6] 吕少中,杜文亮,陈震,等.基于机器视觉的荞麦剥壳性能参数在线检测方法[J].农业机械学报,2019,50(10):35-43.
[7] 吕少中. 基于机器视觉的荞麦剥壳性能参数在线检测方法研究[D].呼和浩特:内蒙古农业大学,2019:65-80.
[8] 柳冠伊,刘平义,魏文军,等.玉米果穗粘连籽粒图像分割方法[J].农业机械学报,2014,45(9):285-290.
[9] 王刚. 基于机器视觉的玉米千粒重快速检测仪的研制[D].长春:吉林大学,2012:18-39.
[10] 杨蜀秦. 农作物籽粒的图像处理和识别方法研究[D].杨凌:西北农林科技大学,2012:15-37.
[11] 王传宇,郭新宇,温维亮,等.基于计算机视觉的玉米籽粒形态测量[J].农机化研究,2011,33(6):141-144.
[12] 李永锋,周德祥,邢超,等.基于形态分水岭算法的粘连大米籽粒分割研究[J].计算机与信息技术,2010(10):42-45.
[13] 匡芳君,徐蔚鸿,王艳华.基于改进分水岭算法的粘连大米图像分割[J].粮食与饲料工业,2010(8):5-8.
[14] 时玉强. 基于机器视觉的大豆品质的研究[D].哈尔滨:东北农业大学,2009:21-28.
[15] 杨冬平.基于梯度改进Hough变换的油菜籽图像分割方法[J].粮油食品科技,2016,24(4):80-82.
[16] 李永锋,周德祥,邢超,等.基于数学形态学的粘连大米籽粒分割研究[J].农业网络信息,2010(10):18-21.
[17] 周德祥,毋桂萍,杨红卫,等.基于数学形态学粘连粮食籽粒图像分割算法的改进[J].农机化研究,2010,32(7):49-52.
[18] 荀一,鲍官军,杨庆华,等.粘连玉米籽粒图像的自动分割方法[J].农业机械学报,2010,41(4):163-167.
[19] 王康,梁秀英,曾优,等.玉米籽粒大小特征检测方法研究[J].湖南农业大学学报(自然科学版),2017,43(3):329-335.
[20] 臧晶,郭倩倩,李成华,等.复杂条件下多玉米籽粒识别与统计方法研究[J].沈阳农业大学学报,2014,45(5):633-636.
[21] 奚德君.面向定向种植的玉米粒群批量整列与分选系统的研制[D].哈尔滨:东北农业大学,2018:48.
[22] 杜世伟,李毅念,姚敏,等.基于小麦穗部小穗图像分割的籽粒计数方法[J].南京农业大学学报,2018,41(4):742-751.
[23] 杨蜀秦,宁纪锋,何东健.一种基于主动轮廓模型的连接米粒图像分割算法[J].农业工程学报,2010,26(2):207-211.
[24] 张新伟,易克传,刘向东,等.玉米自动化考种过程的粘连籽粒图像分割[J].中国农业大学学报,2018,23(10):144-151.
[25] 深度学习[EB/OL].[2021-01-15].https://baike.so.com/doc/7000002-7222884.html.
[26] 谢元澄,于增源,姜海燕,等.小麦麦穗几何表型测量的精准分割方法研究[J].南京农业大学学报,2019,42(5):956-966.
[27] 朱荣胜,闫学慧,陈庆山.基于图像识别和卷积神经网络的大豆优良籽粒筛选研究[J].大豆科学,2020,39(2):189-197.
[28] 陈进,韩梦娜,练毅,等.基于U-Net模型的含杂水稻籽粒图像分割[J].农业工程学报,2020,36(10):174-180.
[29] 孙志恒. 基于深度学习的稻米垩白识别算法研究与应用[D].成都:电子科技大学,2019:24-39.
[30] 张芳. 基于深度学习的玉米果穗分类识别[D].南昌:江西农业大学,2019:23-30.
[31] 王建宇. 基于卷积神经网络的玉米籽粒精选系统研制[D].哈尔滨:东北农业大学,2019:41-64.
[32] 栾争光. 基于卷积神经网络的种子智能分选算法研究[D].郑州:中原工学院,2020:8-46.
[33] 张博. 基于深度学习的小麦外观品质机器视觉检测研究[D].杨凌:西北农林科技大学,2019:21-43.
[34] 祝诗平,卓佳鑫,黄华,等.基于CNN的小麦籽粒完整性图像检测系统[J].农业机械学报,2020,51(5):36-42.
[35] 于重重,周兰,王鑫,等.基于CNN神经网络的小麦不完善粒高光谱检测[J].食品科学,2017,38(24):283-287.
[36] 林萍,张华哲,何坚强,等.可见光谱图与深度神经网络的垩白大米检测方法[J].光谱学与光谱分析,2020,40(1):233-238.
[37] 朱江波. 基于嵌入式平台的颗粒状农作物图像分类算法研究[D].哈尔滨:哈尔滨工业大学,2019:34-62.
[38] 赵志衡,宋欢,朱江波,等.基于卷积神经网络的花生籽粒完整性识别算法及应用[J].农业工程学报,2018,34(21):195-201.
[39] 马志宏. 基于深度学习的水稻粒穗复杂性状图像分析方法[D].上海:上海交通大学,2018:33-55.
[40] 蔡改贫,刘占,汪龙,等.基于形态学优化处理的标记符分水岭矿石图像分割[J].科学技术与工程,2020,20(23):9497-9502.
[41] 陈然. 基于双门限阈值的爆堆岩块图像分割技术及图像识别系统开发[D].衡阳:南华大学,2020:19-48.
[42] 刘晓敏,茅健.非均匀光照金刚石切割线颗粒提取方法[J].轻工机械,2019,37(6):64-70.
[43] 孙国栋,林凯,高媛,等.基于改进亲和度图的矿石颗粒图像分割研究与实现[J].仪表技术与传感器,2019(12):114-118.
[44] 谢雅君,张国英.混配矿石图像的分割优化及级配检测算法[J].有色金属(矿山部分),2019,71(6):110-115.
[45] 黄友亮,孙广彤,战胜,等.基于自适应中值滤波高炉渣颗粒小波去噪[J].矿业研究与开发,2019,39(9):141-145.
[46] 孙颖. 机器视觉用于燃料粒度在线检测研究[D].太原:山西大學,2019:5-22.
[47] 郭挺. 基于视频图像的降雨识别方法研究[D].南京:南京信息工程大学,2019:12-41.
[48] 张庆杭. 基于图像处理的重叠类圆颗粒物的计数及APP的研究开发[D].镇江:江苏大学,2019:17-72.
[49] 展彦彤. 基于多尺度特征分析的矿石粒度分布方法研究[D].北京:中国矿业大学(北京),2019:21-35.
[50] 刘小燕,吴鑫,孙炜,等.基于形态学重建和GMM的球团颗粒图像分割[J].仪器仪表学报,2019,40(3):230-238.
[51] 张文晔. 基于机器视觉的金刚砂线颗粒检测技术的研究与应用[D].常州:江苏理工学院,2018:20-27.
[52] 焦慧君. 拍摄深度限定的溶液晶体粒度分布图像识别方法研究[D].济南:山东大学,2018:11-34.
[53] 王晓迎. 细胞追踪的灰度图像准确分割方法研究[D].徐州:中国矿业大学,2014:25-36.
[54] 张瑞华. 医学显微细胞图像分割研究[D].武汉:武汉科技大学,2014:38-83.
[55] 谢涛. 基于深度学习的微细粒矿物识别研究[D].徐州:中国矿业大学,2020:23-48.
[56] 陈园园. 基于深度学习的GCr15轴承钢金相组织图像分割技术研究[D].兰州:兰州理工大学,2020:37-46.
[57] HEFFERNAN S T,LY N C,MOWER B J,et al. Identifying surface morphological characteristics to differentiate between mixtures of U3O8 synthesized from ammonium diuranate and uranyl peroxide[J]. Radiochimica acta,2019,108(1):29-36.
[58] XIANG D,CAI D,HU X N,et al.A deep learning algorithm for particle segmentation of aerosol images[J]. Journal of flow visualization and image processing,2019,26(3):239-252.
[59] 吴玥. 基于MaskR-CNN分割的TEM纳米颗粒参数测量方法研究[D].天津:天津工业大学,2019:34-41.
[60] 蔡杨,苏明旭,蔡小舒.基于卷积神经网络的混合颗粒分类法研究[J].光学学报,2019,39(7):123-132.
[61] 张芳,吴玥,肖志涛,等.基于U-Net卷积神经网络的纳米颗粒分割[J].激光与光电子学进展,2019,56(6):137-143.
[62] 徐江川. 基于深度卷积神经网络的熟料颗粒方法研究[D].合肥:中国科学技术大学,2018:18-52.
安徽农业科学,J.Anhui Agric.Sci. 2022,50(4):17-20
安徽农业科学 2022年
氮代谢相关酶的研究进展
徐洪超1,商 靖1,刘铭荟1,张 飞2*,李玥莹1*
(1.沈阳师范大学生命科学学院,辽宁沈阳 110034;2.辽宁省农业科学院高粱研究所,辽宁沈阳 110161)
摘要 氮代谢是植物体内的基本生理代谢过程之一,包括氮素同化、积累和蛋白质合成等过程,与植物的生长发育、产量和品质的联系非常密切。氮代谢的生理过程在酶的催化下完成,与氮代谢的生理过程密切相关的酶有:硝酸还原酶、谷氨酰胺合成酶、转氨酶(谷氨酸草酰乙酸转氨酶、谷氨酸丙酮酸转氨酶)、谷氨酸合酶、谷氨酸脱氢酶等关键酶。这5种酶在植物氮代谢过程中具有重要的作用,简要介绍这几种酶的基本结构与特性、对作物生长发育的调控等,为进一步探究氮代谢研究机制提供有益的参考。
关键词 硝酸还原酶;谷氨酰胺合成酶;谷氨酸合酶;转氨酶;谷氨酸脱氢酶
中图分类号 Q 945.1文献标识码 A
文章编号 0517-6611(2022)04-0017-04
doi:10.3969/j.issn.0517-6611.2022.04.005
开放科学(资源服务)标识码(OSID):
Research Progress of Enzymes Related to Nitrogen Metabolism
XU Hong-chao,SHANG Jing,LIU Ming-hui et al (College of Life Sciences, Shenyang Normal University,Shenyang,Liaoning 110034)
Abstract Nitrogen metabolism is one of the basic physiological metabolic processes in plants, including processes such as nitrogen assimilation, accumulation and protein synthesis. It is closely related to plant growth, yield and quality. The physiological process of nitrogen metabolism is completed under the catalysis of enzymes. The enzymes closely related to the physiological process of nitrogen metabolism are: nitrate reductase, glutamine synthetase, transaminase (glutamate oxaloacetate transaminase, glutamate pyruvate transaminase ), glutamate synthase, glutamate dehydrogenase and other key enzymes. These five enzymes play an important role in the process of plant nitrogen metabolism. The basic structure and characteristics of these enzymes and the regulation of crop growth and development are briefly introduced to provide a useful reference for further exploring the research mechanism of nitrogen metabolism.
Key words Nitrate reductase;Glutamine synthetase;Glutamate synthase;Transaminase;Glutamate dehydrogenase
氮素是植物代谢过程所需要的重要营养元素之一,植物吸收的氮主要分為有机氮化物和无机态氮两类。植物不能直接把空气中的氮气转化为自身利用,它们一般吸收利用土壤或者环境中的NO3-或者NH4+,在相关酶的作用下合成谷氨酰胺和谷氨酸,然后经过转氮作用形成不同的氨基酸,再合成蛋白质。氮代谢与植物的生长发育以及产量等方面有着密切的联系。在植物体内,与氮代谢密切相关的酶主要有5种,这5种酶对作物的生长发育、氮素同化与积累、应对逆境等情况都起着调控作用,因此,笔者针对植物氮代谢过程中的几种酶的研究进展进行概述,以期为进一步探究氮代谢研究机制提供有益的参考。
1 硝酸还原酶
1.1 硝酸还原酶的结构与性质
硝酸还原酶(nitrate reductase,NR)是一种氧化还原酶,存在于细胞质内或细胞膜外。该酶的活性最适pH为7.9左右,最适温度为20~25 ℃。该酶发生作用的平衡常数为1025~1040,反应进行的方向倾向于生成亚硝态氮,所以将硝酸盐还原为亚硝酸盐的反应几乎是单向的[1]。
NR属于二聚体结构,组成NR的每个亚基含有3个辅助因子,即血红素、FAD和钼辅因子,3个区域通过铰链区域连接[2]。不同生物中,NR的分子量是有很大不同的。植物体内与NR的相关基因主要有Nia1和Nia2两个基因。在衣藻属中已证实NR可以将电子从NAD(P)H转移到ARC[3],除此之外,NAD(P)H-NR还可以将电子转移到THB1上,THB1通过其双加氧酶活性催化NO转化为硝酸盐[4]。
NR属于多结构域蛋白中的一种。由于NR蛋白产量低、体积大、结构灵活和复杂等特性,通过X射线衍射分析来了解NR蛋白结构一直没有实现。经过长时间的研究试验,玉米NR的NADH/FAD和血红素结构域以及烟草的血红素结构域蛋白质结构模型被成功建立[5]。
NR还是一种光合自养生物体内的诱导酶,它的活性与植物体内代谢过程的关系十分密切[6]。NR的活性受到环境因素和生长调节物质等因素的影响,比如光照、硝酸盐、生物节律等因素都能影响NR的表达效果。其中光和硝酸盐浓度对NR的影响尤为明显。目前,NR已相继从茶树[7]、菠菜[8]、甜菜[9]、水稻[10]等多种植物中被克隆出来。
1.2 NR对作物生长发育的影响
NR是植物氮素同化过程中的第一个酶,也是整个硝态氮同化过程的关键酶、限速酶,在植物代谢调控中具有非常重要的地位[11]。该酶可以通过硝酸盐来影响氮代谢,并且与碳代谢也会有一定的联系。
王雷山等[12]研究表明,推迟棉花的播种期,棉花初花期后叶片NR活性与产量呈正相关。杨忠良等[13]对水稻的研究表明,蛋白质含量高的品种幼苗NR活性高于蛋白质含量低的品种,这表明NR活性可以影响作物的质量。张浩玮等[14]对草地早熟禾的研究中发现,氮积累量与NR活性呈显著正相关。在施肥条件相同的情况下,蔬菜中的NR活性提高了肥料的利用率,同时蔬菜的产量也有所增加[15]。这些研究均表明,高活性的NR对植物的生长发育及产量等都有着积极的作用。
1.3 NR对植物抗逆性的调控
植物受到胁迫时,NO可以作为第二信使作出响应,减少伤害。NR可以催化亚硝酸盐还原成NO,从而在植物逆境中发生响应。
俞珊等[16]对苦草进行研究发现,在较低浓度铅胁迫下,较高的NR的活性植物表现出对重金属胁迫的耐性。潘玲玲[17]将黄瓜的NR基因转入到烟草植株内,获得转基因的烟草植株,与野生型相比较,转基因植株可以减少逆境胁迫下产生的生理伤害、提高抗氧化酶活性,缓解NO3-胁迫。谢寅峰等[18]对针叶树幼苗进行研究发现,在干旱胁迫下,NR活性较强的树种抗旱能力也比较强。以上研究均表明NR涉及一些生物和非生物胁迫的响应,可以提高作物的抗性。
2 谷氨酰胺合成酶
2.1 谷氨酰胺合成酶的结构与性质
谷氨酰胺合成酶(glutamine synthetase,GS)是“GS-GOGAT循环”的第一酶[19],广泛存在于高等植物、动物、微生物中。该酶主要的作用是催化谷氨酰胺的合成,是氮代谢的关键酶。谷氨酰胺是植物体内氮的运输形式,可以起到解除氨毒害的作用。在ATP发挥活性的条件下,谷氨酸在GS的催化下可以合成谷氨酰胺[20]。GS还能够与谷氨酸合成酶相偶联,将氨基供给α-氨基酸,所以说GS与植物蛋白质的合成也有着重要的联系。
根据分子量、基因序列等的不同,GS同工酶分为3类,分别为GS Ⅰ、GS Ⅱ和GS Ⅲ,3类同工酶的分布范围有所不同[21]。高等植物的器官中分布着多种GS的同工酶(属GS Ⅱ类),分为GS1、GS2、GSx 3类。其中GS1、GS2发挥的作用比较多,而GSx含量较少,GS1与储存氮源、氮素的转移再利用有关,GS2主要参与硝酸还原产生的氨的同化以及光呼吸过程[22]。GS同工酶的亚基组成的数量和结构有所差别,种类和功能也有所不同。
GS基因的表达受氮素水平、植物发育程度、光照、胁迫因子及氮源形式等多种因素的影响,其中氮素形态及水平的影响是比较重要的[23]。GS基因已经从甜瓜[24]、甜菜[25]、小麦[26]等作物中被克隆。
2.2 GS对作物逆境的调控 氮素是作物生长发育所需要的主要营养元素之一,GS是氮素代谢过程中重要的关键酶。GS可以对逆境中的作物进行调控,提高抗性。
逆境条件可以刺激植物体内GS的活性和相应基因的表达。孙辉等[27]以水稻为研究对象,将GS1和GS2基因整合到转基因水稻的基因组中发现,转基因水稻的抗逆性有所增加。黄其满等[28]在对转基因小麦的研究中发现,GS基因在小麦植株中有效表达,可以使其表现出抗草胺膦特性,相比于对照植株叶片逐渐变黄,转基因小麦植株能正常结实。
2.3 GS对作物氮素同化的调控作用 GS与作物氮的同化吸收和再利用关系密切,该酶可以提高植物对氮素的利用效率,从而影响植株的产量。
旗叶是小麦的主要营养器官,可以进行氮储存和氮同化,与籽粒的产量与品质密切相关,王小纯等[29]对不同专用型小麦进行研究时发现,小麦旗叶GS活性与籽粒蛋白质含量呈显著正相关。Cai等[30]在水稻中的试验发现,在温室中培养超表达GS1的转基因水稻,转基因植株体内可溶蛋白和氮含量均提高。Cruz等[31]在以拟南芥为试验材料进行研究时发现,GS1基因家族中2个基因可以对NH4+刺激作出响应,提高植物根中氮的同化利用效率。
3 谷氨酸合酶
3.1 谷氨酸合酶的结构与性质
谷氨酸合酶(glutamate synthase,GOGAT)是氨同化途径中关键酶,能够与GS偶联,共同完成氨同化的过程。此酶可催化谷氨酰胺的氨基还原转移于α-酮戊二酸,生成谷氨酸。根据电子供体的不同,GOGAT主要分为3种类型:NADH-GOGAT型、Fd-GOGAT型和NADPH-GOGAT型[32]。在高等植物中,前两种形式是比较普遍的。NADH-GOGAT主要與氮类物质的转移运输相关联。
GOGAT基因的表达受氮素、光照、作用部位等的影响[33]。陈阳等[34]对草地早熟禾的研究发现,氮浓度较低时,NADH-GOGAT基因的表达量更多。NADH-GOGAT还受水分、环境等因素的影响。Fd-GOGAT在叶绿体中的活性比较高。Fd-GOGAT基因已经从水稻[35]、马铃薯[36]、大豆[37]等作物中克隆出来。
3.2 GOGAT对氮素积累起调控作用
GOGAT是GS/GOGAT循环同化途径的关键酶之一,GOGAT活性还与氮素积累密切相关。张玮琦[38]在对水稻的研究中发现,在分蘖期,氮素积累量与GOGAT活性呈显著正相关。朱静等[39]对灵芝进行试验研究,推测灵芝GOGAT的缺失是通过影响胞内ROS的稳态来影响灵芝三萜的生物合成,从而影响氮源调控灵芝次级代谢的分子机制。
3.3 GOGAT对作物产量与品质的作用
GOGAT对作物的产量和品质起着积极作用。陈展宇等[40]在对大豆根系的研究试验中发现,大豆苗期、鼓粒期、成熟期,GOGAT活性与产量呈正相关,盛花期和结荚期呈显著正相关。赵全志等[41]在对水稻叶色的研究中发现,GOGAT酶与叶色变化的关系相对密切,影响作物碳氮代谢的强度和产量的形成。刘业丽等[42]对不同基因型的大豆进行研究表明,在R4期的叶片与茎秆中,GOGAT的活性与籽粒蛋白质含量呈正相关。滕祥勇[43]在以甜菜作为研究材料时发现,甜菜叶片中随着Fd-GOGAT活性的提高,产量呈极显著的增加,并且GOGAT的活性与作物中的含糖率关系也十分紧密。
4 谷氨酸脱氢酶
4.1 谷氨酸脱氢酶的结构与性质
谷氨酸脱氢酶(glutamate dehydrogenase,GDH)是一种多聚酶,广泛存在于生物界中[44]。根据辅酶的特异性,可以将GDH分为NADP依赖型、NAD依赖型和非依赖型NAD(P)3类。GDH的相对分子量为255~270 kD,一般由6个亚基组成,分为α和β两种类型,并且含有多种形式的同工酶[45]。在GDH的催化下可以生成谷氨酸,GDH易受外部因素如温度、光照、pH、NO3-、NH4+等影响[46]。到目前为止,GDH基因已经在小麦[47]、玉米[48]、花生[49]等作物中被克隆出来。
4.2 GDH在作物抗逆条件下的响应 GDH是研究植物氮代谢的重要指标之一,可以在植物的衰老过程及逆境等状况下发挥作用。
张彩生[50]在研究水稻抗旱品种过程中,将稻瘟病菌GDH基因在水稻中进行表达时发现,该基因可以改变气孔的开放和增加脯氨酸等小分子的量,使水稻具有较高的抗旱能力。Kumar等[51]在对水稻进行盐胁迫试验时发现,耐盐水稻GDH的氨同化活性比较高。Valliyodan等[52]发现,GDH可以催化谷氨酸合成,使脯氨酸的含量增加,增强了植物的耐逆性。由此可见,GDH可以在逆境条件下发生响应。
4.3 GDH对作物氮素利用率的调控作用 GDH还与作物的氮素利用率关系非常紧密。周香城[53]将真菌埃伦柱孢GDH基因表达到水稻中发现,转基因水稻苗期的氮素利用率有所提高。Egami等[54]将构巢曲霉的NADP(H)-GDH基因在马铃薯中进行过量表达,发现马铃薯的块茎干物质增加,并且在低氮条件下,转基因马铃薯块茎的氮素利用率也有所提高。以上研究均表明,GDH对作物氮素利用率有积极的调控作用。
5 转氨酶
5.1 转氨酶的结构与性质
转氨基作用是植物进行氮代谢的重要过程之一,可以促进氨基酸的合成和分解。植物体内最普遍的转氨酶主要包括2种:谷氨酸草酰乙酸转氨酶(GOT)和谷氨酸丙酮酸转氨酶(GPT)。
GOT又称天门冬氨酸转氨酶,是天门冬氨酸合成的关键酶,在该酶的催化下,L-谷氨酸和草酰乙酸可以反应生成L-门冬氨酸和α-酮戊二酸。GOT是存在于所有真核细胞的胞内酶,该酶最适生长温度为37 ℃,徐凡等[55]对烟草的试验表明,Mn2+、Fe2+、Zn2+对GOT激活作用的程度不同,Ca2+对GOT有抑制作用。
GPT的辅酶为磷酸吡哆醛,是丙氨酸合成的关键酶。根据物种的不同,GPT可分为2~6个亚型,细胞质、线粒体和过氧化酶体中均可以发现其亚型的存在[56]。GPT可以催化L-谷氨酸和丙酮酸生成L-丙氨酸和α-酮戊二酸。郭旭舜等[57]对大豆下胚轴进行试验发现,GPT最适温度为35 ℃,最适pH 7.0,Co2+、Mg2+和Ca2+对其有激活效应,EDTA2-、尿素、Mn2+和Fe2+对其有抑制效应。
5.2 转氨酶在氮素固定方面的作用
转氨基作用可以促进丙酮酸、α-酮戊二酸与氨基酸的互变,对糖和蛋白质代谢产物的相互转变具有重要意义。转氨酶是一种转移酶,该酶可以催化转氨基反应,还能反映叶片氮同化、氨转运和贮藏状态,从而协调氮代谢与碳代谢。
江立庚等[58]对水稻研究发现,GPT活性的变化对氮素吸收利用的影响比较大。提高剑叶GPT活性可以显著增加氮素积累总量,并且硅肥可通过提高叶片中GPT酶活性,促进水稻对氮素的吸收作用。王显等[59]在对水稻的研究中发现,施用氮肥或者硅肥能提高叶片中转氨酶的活性,促进水稻氮代谢水平,提高蛋白质含量。稻米的种类不同,GPT、GOT活性的变化规律也不完全一致。另外,Farnham等[60]在以苜蓿根瘤菌作为试验材料时也发现,GOT在作物氮素固定和吸收方面起着重要的作用。
5.3 转氨酶在抵御非生物胁迫方面发挥的作用
转氨酶不仅在植物的碳氮代谢过程中有着不可或缺的作用,还可以在抵抗非生物胁迫方面发挥重要的调控作用。
Miyashita等[61]在拟南芥研究中发现,缺氧情况下会诱导植物中GPT基因的表达量增加。说明GPT可以在缺氧的情况下发挥调控作用。崔新等[62]在对茶树的试验中发现,高温胁迫下茶树中的GPT可以累积较多的丙氨酸。GOT还有多种同工酶,分布的位置不同使得其功能与活性也不相同。瞿炤珺[63]在以寒地粳稻为试验材料时发现,在冷水胁迫下,該植株根系的酶活性上升,且增幅逐渐增加。
6 结语
植物的氮代谢过程对植物的抗逆作用以及生长发育等方面有着非常重要的意义。植物体内的氮代谢过程是一个需要多种酶相互配合、协调的复杂过程。对氮代谢过程进行详细的探究可以了解植物体内的生理机制,从而培养出符合期望的理想品种。目前对氮代谢相关酶的研究是比较广泛的,并且在很多作物的研究中取得了很大进展。在许多作物中已经克隆出了相关的酶的基因,揭示其影响氮代谢的本质,有助于指导作物改良的过程。
但是仍有一些问题有待解决,例如转氨酶在作物中的研究报道相对较少,在氮代谢和抗逆过程的作用机理也有待深入研究。对于GDH而言,GDH多用于低等植物的研究,高等生物优质GDH的发掘和鉴定还有待研究,这也为下一步的研究提供了方向。
参考文献
[1]
洪华生,王玉珏,王大志.海洋浮游植物硝酸还原酶研究进展[J].海洋科学,2007,31(10):4-10.
[2] 田华,段美洋,王兰.植物硝酸还原酶功能的研究进展[J].中国农学通报,2009,25(10):96-99.
[3] CHAMIZO-AMPUDIA A,SANZ-LUQUE E,LLAMAS ,et al.A dual system formed by the ARC and NR molybdoenzymes mediates nitrite-dependent NO production in Chlamydomonas[J].Plant,cell & environment,2016,39(10):2097-2107.
[4] SANZ-LUQUE E,OCA A-CALAHORRO F,DE MONTAIGU A,et al.THB1,a truncated hemoglobin, modulates nitric oxide levels and nitrate reductase activity[J].The plant journal,2015,81(3):467-479.
[5] CAMPBELL W H.Nitrate reductase biochemistry comes of age[J].Plant physiology,1996,111(2):355-361.
[6] 汤玉玮,林振武,陈敬祥.硝酸还原酶活力与作物耐肥性的相关性及其在生化育种上应用的探讨[J].中国农业科学,1985,18(6):39-45.
[7] 周月琴,庞磊,李叶云,等.茶树硝酸还原酶基因克隆及表达分析[J].西北植物学报,2013,33(7):1292-1297.
[8] 程祖锌,何海华,黄志伟,等.菠菜硝酸还原酶基因的克隆与原核表达[J].江苏农业科学,2012,40(12):19-22.
[9] 曾彦达.甜菜亚硝酸还原酶基因克隆及生物信息学分析[D].哈尔滨:东北农业大学,2011.
[10] 胡静,吴文华.水稻硝酸还原酶基因5′上游序列的克隆与序列分析[J].华中师范大学学报(自然科学版),2009,43(2):284-288.
[11] 张华珍,徐恒玉.植物氮素同化过程中相关酶的研究进展[J].北方园艺,2011(20):180-183.
[12] 王雷山,KHAN AZIZ,宋兴虎,等.棉花主茎叶与根系硝酸还原酶活性分布对播期和密度的响应[J].棉花学报,2017,29(1):88-98.
[13] 杨忠良,刘海英,刘会,等.氮素对水稻幼苗氮代谢相关酶活性及相关基因表达的影响[J].黑龙江农业科学,2017(10):26-31.
[14] 張浩玮,白小明,陈辉,等.草地早熟禾(Poa pratensis)N积累、分配和硝酸还原酶活性对氮、磷肥的响应[J].中国沙漠,2019,39(5):222-231.
[15] MOROZKINA E V,ZVYAGILSKAYA R A.Nitrate reductases:Structure,functions,and effect of stress factors[J].Biochemistry (Moscow),2007,72(10):1151-1160.
[16] 俞珊,张光生,成小英.铅胁迫对苦草生理生化及氮、磷代谢关键酶的影响[J].安全与环境学报,2016,16(1):372-376.
[17] 潘玲玲.黄瓜CsNR基因的克隆、遗传转化及在硝酸盐胁迫下的功能分析[D].泰安:山东农业大学,2017.
[18] 谢寅峰,沈惠娟.水分胁迫下3种针叶树幼苗抗旱性与硝酸还原酶和超氧化物歧化酶活性的关系[J].浙江林学院学报,2000,17(1):24-27.
[19] FORDE B G,LEA P J.Glutamate in plants:Metabolism,regulation,and signalling[J]. Journal of experimental botany,2007,58(9):2339-2358.
[20] 刘芸菲,薛栋升,宫春杰.谷氨酰胺合成酶研究进展[J].山东化工,2021,50(5):97-99,101.
[21] 赵凯琴,罗延青,俎峰,等.谷氨酰胺合成酶及其在植物基因工程中应用研究进展[J].南方农业学报,2012,43(4):435-438.
[22] 王小纯,张同勋,李高飞,等.小麦谷氨酰胺合成酶基因克隆与其表达特性分析[J].河南农业大学学报,2012,46(5):487-492.
[23] 孙敏红,吴炼,谢深喜.铵硝营养对枳橙幼苗谷氨酰胺合成酶活性及相关基因表达的影响[J].植物生理学报,2018,54(11):1703-1710.
[24] 管闪青.甜瓜谷氨酰胺合成酶基因的克隆及其表达分析研究[D].上海:上海交通大学,2007.
[25] 王淑春.甜菜谷氨酰胺合成酶基因的克隆[D].哈尔滨:黑龙江大学,2007.
[26] 李高飞.小麦谷氨酰胺合成酶基因克隆及其苗期表达调控[D].郑州:河南农业大学,2011.
[27] 孙辉,黄其满,苏金.谷氨酰胺合成酶基因GS1和GS2的高效表达增强转基因水稻对氮素缺乏的耐性(英文)[J].植物生理与分子生物学学报,2005,31(5):492-498.
[28] 黄其满,刘伟华,孙辉,等.农杆菌介导的转谷氨酰胺合成酶基因小麦的抗除草剂特性研究(英文)[J].植物生态学报,2005,29(2):338-344.
[29] 王小纯,熊淑萍,马新明,等.不同形态氮素对专用型小麦花后氮代谢关键酶活性及籽粒蛋白质含量的影响[J].生态学报,2005,25(4):802-807.
[30] CAI H M,ZHOU Y,XIAO J H,et al.Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice[J].Plant cell reports,2009,28(3): 527-537.
[31] CRUZ C,BIO A F M,DOMNGUEZ-VALDIVIA M D,et al.How does glutamine synthetase activity determine plant tolerance to ammonium?[J].Planta,2006,223(5):1068-1080.
[32] OLIVER G,GOSSET G,SANCHEZ-PESCADOR R,et al.Determination of the nucleotide sequence for the glutamate synthase structural genes of Escherichia coli K-12[J].Gene,1987,60(1):1-11.
[33] 牛超,刘关君,曲春浦,等.谷氨酸合成酶基因及其在植物氮代谢中的调节作用综述[J].江苏农业科学,2018,46(9):10-16.
[34] 陈阳,孙华山,王玉书,等.草地早熟禾NADH-GOGAT基因的克隆及表达分析[J].草地学报,2019,27(2):459-465.
[35] 孙立亭.水稻类病斑基因Fd-GOGAT的功能分析和白条纹突变体K170的基因定位[D].南京:南京农业大学,2015.
[36] 赵艳菲,张嘉越,韩玉珠.马铃薯氮代谢途径中Fd-GOGAT基因的克隆及生物信息学分析[C]//金黎平,吕文河.马铃薯产业与美丽乡村(2020).哈尔滨:黑龙江科学技术出版社,2020:309-313.
[37] 陈丽华,刘丽君,刘页丽,等.不同基因型大豆Fd-GOGAT基因cDNA序列的克隆与分析[J].大豆科学,2011,30(3):374-378.
[38] 张玮琦.氮肥对水稻氮代谢酶活性和氮素利用的影响[D].沈阳:沈阳农业大学,2019.
[39] 朱静,岳思宁,陈琛,等.谷氨酸合酶在灵芝中生物学功能的研究[J].南京农业大学学报,2019,42(6):1073-1079.
[40] 陈展宇,房晓琨,孙帆,等.不同年代大豆品种根系氮代谢关键酶活性变化的研究[J/OL].吉林农业大学学报,2020-12-25[2021-05-25].https://kns.cnki.net/kcms/detail/22.1100.s.20201225.0934.002.html.
[41] 趙全志,陈静蕊,刘辉,等.水稻氮素同化关键酶活性与叶色变化的关系[J].中国农业科学,2008,41(9):2607-2616.
[42] 刘业丽,栾怀海,何琳,等.不同基因型大豆NADH-GOGAT活性动态规律研究[J].大豆科学,2014,33(2):199-202.
[43] 滕祥勇.甜菜谷氨酸合成酶(GOGAT)酶学特性及氮素对其酶活性的调控[D].哈尔滨:东北农业大学,2012.
[44] 龚茵茵,燕璐,林建中,等.低等生物谷氨酸脱氢酶基因用于作物遗传改良的研究进展[J].生命科学研究,2021,25(1):31-38.
[45] 黄国存,田波.高等植物中的谷氨酸脱氢酶及其生理作用[J].植物学通报,2001,36(4):396-401.
[46] 邱旭华.水稻氮代谢基础研究:谷氨酸脱氢酶作用的分子机理[D].武汉:华中农业大学,2009.
[47] 李冰.小麦氮利用效率相关基因TaGDH的克隆与功能分析[D].泰安:山东农业大学,2014.
[48] 曹爽.玉米抗冷基因ZmGDH2的功能分析及遗传转化的研究[D].长春:吉林大学,2017.
[49] 陈湘瑜,徐日荣,林栩松,等.花生谷氨酸脱氢酶基因AhGDH1的克隆与生物信息学分析[J].福建农业学报,2016,31(3):217-224.
[50] 张彩生.异源表达稻瘟病菌MgGDH基因对水稻干旱胁迫影响的研究[D].长沙:湖南大学,2015.
[51] KUMAR R G,SHAH K,DUBEY R S.Salinity induced behavioural changes in malate dehydrogenase and glutamate dehydrogenase activities in rice seedlings of differing salt tolerance[J].Plant science,2000,156(1):23-34.
[52] VALLIYODAN B,NGUYEN H T.Understanding regulatory networks and engineering for enhanced drought tolerance in plants[J].Current opinion in plant biology,2006,9(2):189-195.
[53] 周香城.异源表达埃伦柱孢谷氨酸脱氢酶基因CeGDH对水稻氮素利用的影响[D].长沙:湖南大学,2014.
[54] EGAMI T,WAKAYAMA M,AOKI N,et al.The effects of introduction of a fungal glutamate dehydrogenase gene (gdhA) on the photosynthetic rates,biomass,carbon and nitrogen contents in transgenic potato[J].Plant biotechnology,2012,29(1):57-64.
[55] 徐凡,吳媚.烟草谷草转氨酶纯化及部分酶学性质分析[J].广东农业科学,2013,40(10):146-148,175.
[56] LIEPMAN A H,OLSEN L J.Genomic analysis of aminotransferases in Arabidopsis thaliana[J].Critical reviews in plant sciences,2004,23(1):73-89.
[57] 郭旭舜,林裕添,贺敬东,等.大豆下胚轴谷丙转氨酶酶学性质分析[J].大豆科学,2021,40(1):82-88.
[58] 江立庚,曹卫星,甘秀芹,等.水稻氮素吸收、利用与硅素营养的关系[J].中国农业科学,2004,37(5):648-655.
[59] 王显,张国良,霍中洋,等.氮硅配施对水稻叶片光合作用和氮代谢酶活性的影响[J].扬州大学学报(农业与生命科学版),2010,31(3):44-49.
[60] FARNHAM M W,DEGENHART N R,VANCE C P,et al.Molecular and whole-plant responses to selection for enzyme activity in alfalfa root nodules:Evidence for molecular compensation of aspartate aminotransferase expression[J].Theoretical and applied genetics,1992,84(3/7):355-361.
[61] MIYASHITA Y,DOLFERUS R,ISMOND K P,et al.Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana[J].The plant journal,2007,49(6):1108-1121.
[62] 崔新,刘志薇,吴致君,等.茶树谷丙转氨酶基因的克隆及其表达分析[J].西北植物学报,2016,36(12):2361-2369.
[63] 瞿炤珺.孕穗期冷水胁迫对寒地粳稻根系生长及氮代谢的影响[D].哈尔滨:东北农业大学,2019.
