白及根腐病植株根际土壤微生物群落组成特征分析
2022-03-10赵林艳官会林向萍李泽诚柏雨龙宋洪川孙世中徐武美
赵林艳 官会林 向萍 李泽诚 柏雨龙 宋洪川 孙世中 徐武美
(1. 云南师范大学 高原特色中药材种植土壤质量演变退化与修复云南省野外科学观测研究站,昆明 650500;2. 西南林业大学环境修复与健康研究院,昆明 650224;3. 昆明英武农业科技有限公司,昆明 650500)
白及(Bletilla striata)为兰科白及属多年生草本植物,具有收敛止血、消肿生肌等功效,常用于咳血吐血、外伤止血、疮疡肿毒、皮肤皲裂、烫灼伤、肛裂等疾病[1]。我国有4种白及属植物,其分布区北起江苏、河南,南至我国台湾,东起浙江,西至西藏东南部。白及人工种植区主要位于云南、贵州、四川等地,其中云南种植面积最大,约占全国种植面积的42%[2-3]。随着白及种植年限的增加,土壤致病菌的积累也相应增加,植物病害较为严重,特别是根腐病,以植物地下部分腐烂为特征,具体表现为块茎呈水渍状腐烂,随后扩散至根部,使其发黑坏死,叶片出现褐色长型枯斑,最终导致全株枯死[4],严重影响白及产量与经济效益。
根腐病是根茎类药用植物栽培过程中的一类高发病害,其致病因素较为复杂。病原真菌被认为是导致根腐病的主要原因之一[5]。廖作清等[6]研究发现,尖孢镰刀菌(Fusarium oxysporum)、腐皮镰刀菌(F.solani)、立枯丝核菌(Rhizoctonia solani)、恶疫霉病菌(Phytophthora cactorum)等病原真菌可引起三七根腐病;毕武等[7]发现尖孢镰刀菌、人参锈腐病菌(Cylindrocarpon destructans)等会导致西洋参根腐病。已有研究证实有丝核菌属(Rhizoctonia)和腐皮镰刀菌(Fusarium solani)可导致白及根腐病[8-9],但尚未发现更多与白及根腐病相关的病原微生物。
根际微生物群落组成特征对作物营养、生长与抗病害等均具有重要影响[10-11]。根际微生物多样性、相对丰度与植物病害有着密切关联性[12-13],如黄芪根际土壤细菌群落多样性降低会加重根腐病害,且在发病黄芪根际土壤中有益微生物丰度也呈下降趋势[14]。同样,大蒜根腐病发病土壤中细菌多样性降低,镰刀菌属(Fusarium)和匍柄霉菌属(Stemphylium)等病原真菌相对丰度增加,而硝化螺旋菌属(Nitrospira)、芽孢杆菌属(Bacillus)、木霉属(Trichoderma)等有益微生物相对丰度降低[15]。Wu等[16]研究发现,根腐三七根际土壤微生物群落与健康植株相比有明显差异,其中患病三七根际土壤中放线菌、革兰氏阳性菌、丛枝菌根真菌等的相对丰度显著降低。此外,研究还发现西洋参根腐病株根际土壤真菌丰度显著低于健康植株,且红游动菌属(Rhodoplanes)、Kaistobacter、鞘脂菌属(Sphingobium)等的相对丰度与西洋参根腐病害呈正相关[17-18]。然而,针对白及根腐病植株根际土壤微生物群落组成特征相关研究还未见报道。因此,本研究利用高通量测序技术分析了白及根腐病与健康植株根际土壤微生物群落组成与变化特征,并对土壤理化性质与酶活性进行了定量分析,旨在为了解白及根腐病发病机理,科学应对白及根腐病害,提高白及产量与经济效益提供理论依据。
1 材料与方法
1.1 材料
白及根际土壤样品采自位于昆明市呈贡新区的昆明英武农业科技有限公司科研基地,土壤类型为旱地红壤,白及连作8年。白及品种为广泛栽种的三叉白及,产量高、具有较强的耐寒性,其田间种植密度为50株/m2。按照随机均衡原则,在科研基地内选择已移栽2年的根腐发病和健康白及各5株,利用无菌试管采集粘附在根茎上的土壤,随即保存于-80℃超低温冰箱,用于微生物群落分析,并采集距离根茎3 cm内的土样,去除石砾、残根等杂物,自然风干用于土壤理化性质与酶活性分析。
1.2 方法
1.2.1 土壤DNA的提取与高通量测序 称取1 g白及根际土壤,用DNA提取试剂盒(NucleoSpin Kit,Macherey-Nagel GmbH,Germany)提取样品总DNA。使 用 引 物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′) 对细菌16S rRNA V3-V4区进行PCR扩增,使用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′) 和ITS2(5′-GCTGCGTTCTTCATCGATGC-3′)对 真 菌ITS片段进行扩增。PCR扩增产物使用AMPure XP试 剂 盒(Beckman Coulter Life Sciences,USA)和QuantiFluorTM-ST荧光定量系统(Promega,USA)进行纯化和定量,利用Illumina HiSeq 2500 PE250进行高通量测序。
1.2.2 土壤理化性质与酶活性的测定 土壤自然风干后研磨过20目和100目筛,分别用于理化性质和酶活性的测定。土壤理化性质的测定方法参照《土壤农化分析》[19]:土壤pH使用pH计(雷磁PHS-25)测定;电导率使用EC计(COMBI 5000)测定;氨态氮(NH4+-N)和硝态氮(NO3--N)含量使用连续流动分析仪(Seal Analytical AA3)测定;有效磷(AP)含量使用钼锑抗比色法,结合紫外可见分光光度计(元析UV-8000)测定;有效钾(AK)含量使用连续流动火焰分光光度计(AA3,Model 410)测定;有机质含量使用浓硫酸-重铬酸钾溶液消解法,用硫酸亚铁标准溶液进行滴定。土壤脲酶和蔗糖酶活性,分别用3,5-二硝基水杨酸比色法与靛酚蓝比色法进行测定[20];磷酸酶活性用磷酸苯二钠比色法进行测定[21];蛋白酶活性用茚三酮比色法进行测定[22]。
1.2.3 数据分析 利用FLASH V1.2.7[23]和Trimmomatic[24]软件对Illumina HiSeq双端测序数据进行拼接与过滤,使用UCHIME v4.2[25]去除嵌合体,并用UPARSE[26]软件按97%的序列相似性获得OTU(operational taxonomic units)。使用RDP软件以细菌16S rRNA Silva数 据 库(http://www.arb-silva.de)和真菌ITS UNITE数据库(https://unite.ut.ee/)为参照对不同OTU的代表性序列进行分类学注释。使用MOTHUR[27]软件计算各土样微生物OTU丰富度、Chao1与Shannon多样性指数。
基于Bray-Curtis距离指数在OTU分类水平进行不同微生物群落的UPGMA聚类分析,并利用冗余分析(RDA)探索健康与根腐白及根际土壤微生物群落的分化。利用独立样本t检验分析不同土壤因子的差异显著性。以上分析利用R软件包“vegan”[28-29]与SPSS 16.0统计软件(SPSS Inc.,Chicago,IL)进行。
2 结果
2.1 根腐病对白及根际微生物群落组成的影响
门水平上,Ascomycota和Mortierellomycota为白及根际土壤优势真菌类群,在发病组中的占比为73.43%和11.78%(表1),Ascomycota相对丰度显著高于健康组(40.92%),而Mortierellomycota相对丰度显著低于健康组(42.50%)。Proteobacteria和Bacteroidetes为优势细菌类群,在发病组中的占比分别为24.71%和28.24%,在健康组中的占比分别为28.36%和22.57%,无显著差异。属水平上,发病组Fusarium、Erysiphe等真菌的相对丰度显著高于健康组,而Microscypha、Hannaella等真菌的相对丰度显著降低(图1)。发病组中,Akkermansia、IMCC26134与Larkinella等细菌的相对丰度显著低。
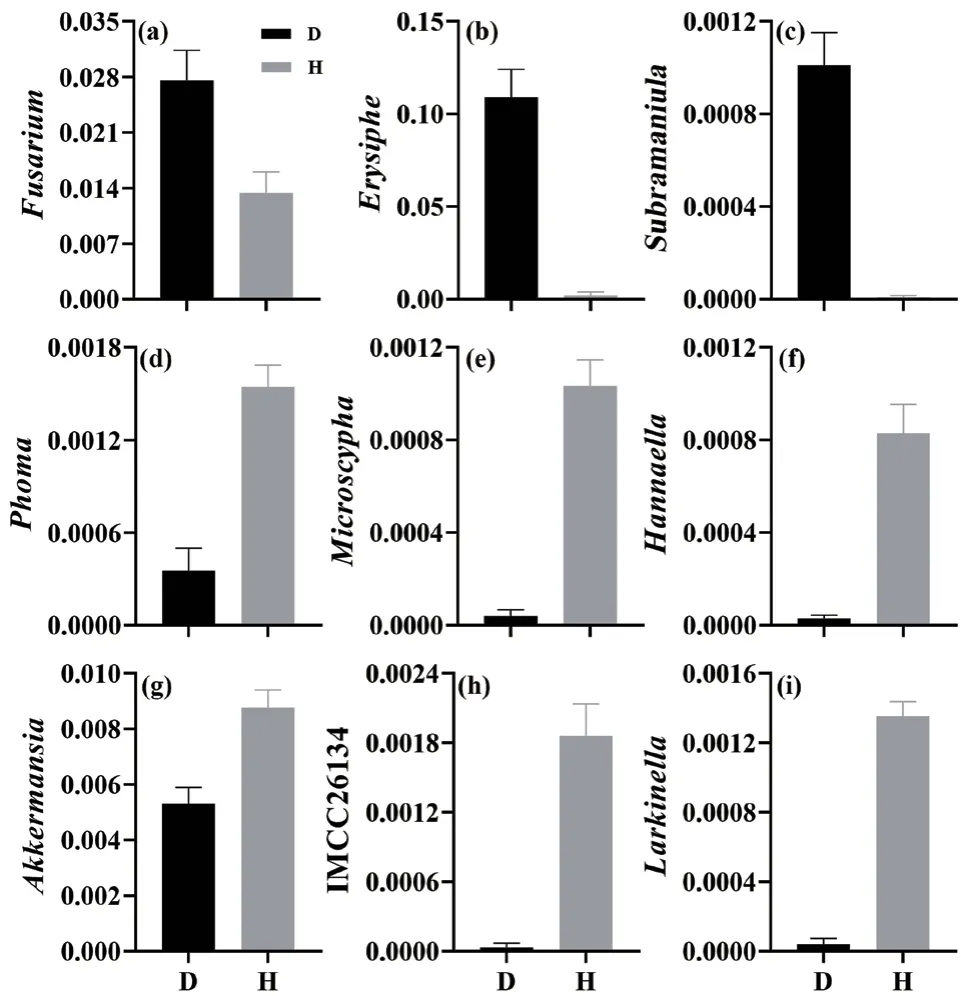
图1 白及根腐病和健康植株根际土壤微生物属分类水平的相对丰度Fig. 1 Relative abundance of rhizospheric microorganisms at the genus level in the rhizospheric soils of root-rot and healthy B. striata
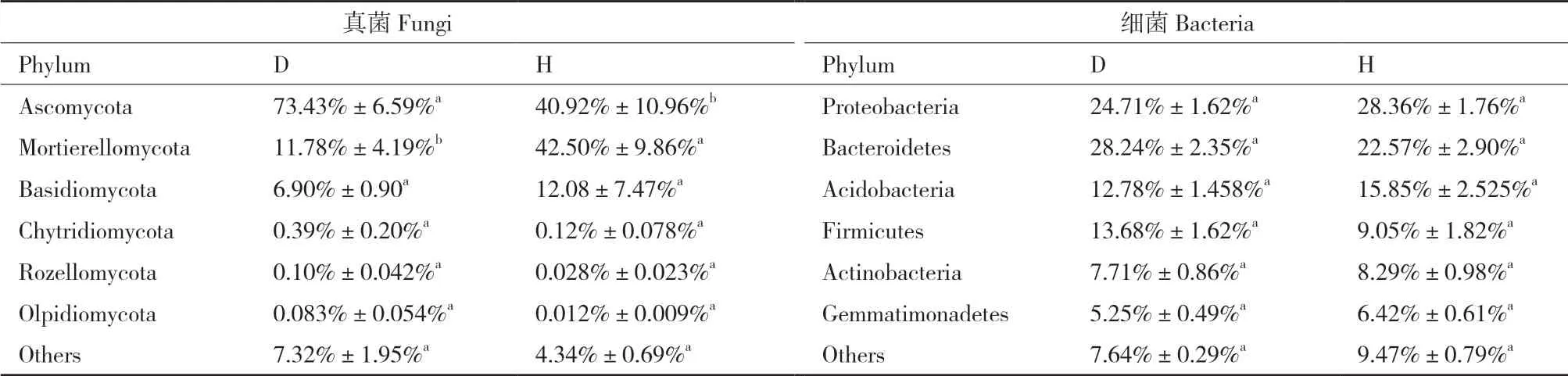
表1 白及根腐病和健康植株根际土壤微生物门水平的相对丰度Table 1 Relative abundance of microorganisms at the phylum level in the rhizospheric soils of root-rot and healthy B. striata
2.2 根腐病对白及根际微生物群落alpha多样性的影响
原始测序数据经拼接、过滤与嵌合体检测,并按97%的序列相似性进行划分,共得到真菌OTU 924个,隶属于9门、24纲、60目、120科、207属与214种,得到细菌OTU 1 690个,隶属于25门、65纲、139目、227科、451属与476种。根腐白及根际土壤中特有真菌和细菌的OTU数为 82和1个,而健康植株根际土壤特有真菌和细菌的OTU数为18和2个(图2),表明与健康植株相比较,根腐白及根际土壤真菌群落差异较大,而细菌群落组成差异相对较小。根腐白及根际土壤真菌OTU丰富度显著高于健康植株,而细菌OTU丰富度无显著差异,表明白及患根腐病后,真菌alpha多样性呈升高趋势(表2)。

表2 白及根腐病和健康植株根际土壤微生物群落alpha多样性指数Table 2 Alpha diversity of microbial community in the rhizospheric soils of root-rot and healthy B. striata
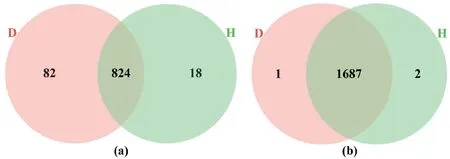
图2 白及根腐病和健康植株根际土壤共有和特有的OTU数量Fig. 2 Numbers of shared and unique OTUs in the rhizospheric soils of root-rot and healthy B. striata
2.3 根腐病对白及根际微生物群落beta多样性的影响
UPGMA聚类分析表明,发病组与健康组土壤真菌和细菌群落具有明显聚类特征(图3-a,c),冗余分析表明,发病组与健康组真菌群落在第二排序轴具有明显的分化特征(图3-b),而细菌群落在第一排序轴具有明显的分化特征(图3-d);与细菌群落相比较,真菌群落分化更明显。
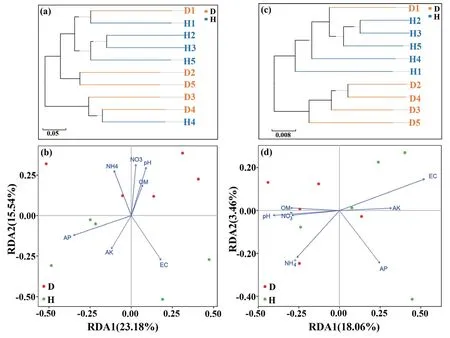
图3 基于聚类与冗余分析的白及根腐病和健康植株根际土壤微生物群落差异特征Fig. 3 Differences in the rhizospheric soil microbial community of root-rot and healthy B.striata based on clustering and redundancy analyses
2.4 白及根腐病和健康植株根际土壤理化性质与酶活性
与健康植株相比较,白及根腐病植株根际土壤有效钾含量较低,pH、有机质、铵态氮与硝态氮含量较高(P<0.05),而有效磷含量无显著差异(P>0.05);土壤脲酶、蛋白酶和蔗糖酶的活性较低。因此,土壤养分失衡、酶活性降低可能促进了白及根腐病的发生(表3)。
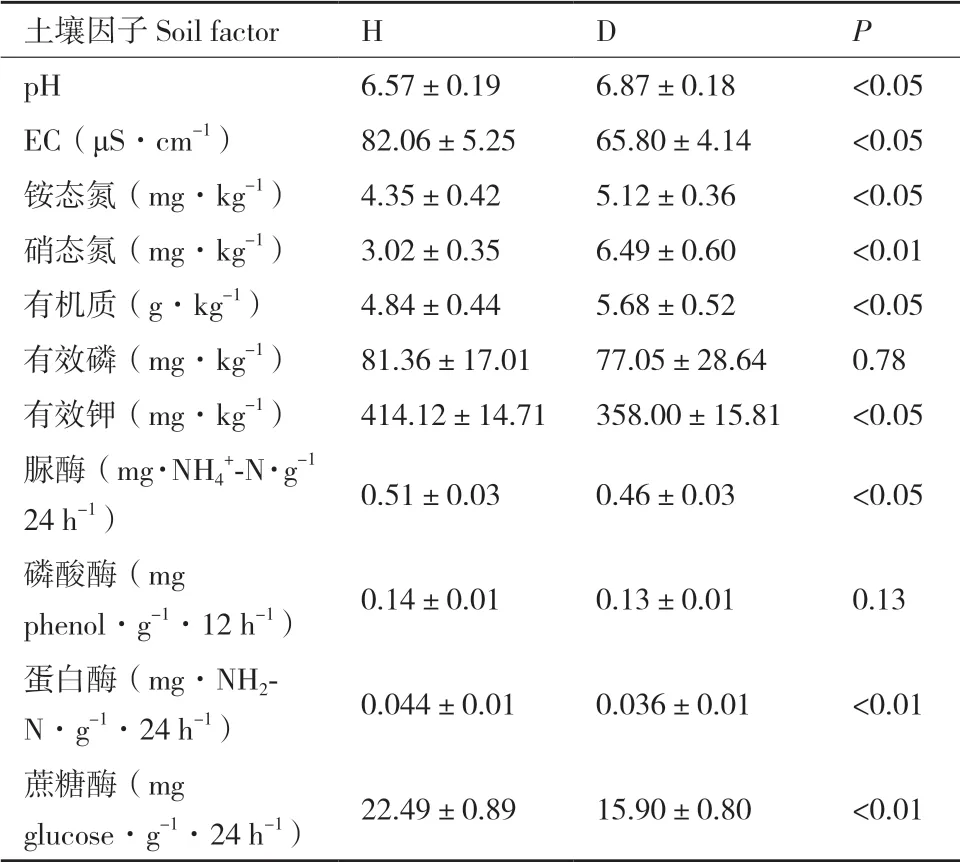
表3 白及根腐病和健康植株根际土壤理化性质与酶活性Table 3 Physicochemical properties and enzyme activities in the rhizospheric soils of root-rot and healthy B.striata
3 讨论
土壤微生物在农业生态系统养分循环和肥力维持等方面发挥着重要作用,且对植物健康生长具有直接或间接的影响[30-31]。土壤微生物群落组成特征在一定程度上反映了土壤健康状况,当土壤中病原微生物大量生长,有益微生物数量减少时,常引起作物病害的发生[32-33]。在本研究中,白及根腐病植株根际土壤Fusarium、Erysiph等菌群相对丰度显著增加,且与健康植株相比较,真菌和细菌群落具有明显的分化特征,表明白及根腐病的发生与土壤微生物群落组成变化密切相关。
土壤中存在着大量植物病原真菌,Fusarium可引起白及、三七、西洋参等根茎类药用植物发生根腐病[6-9],且这些药用植物根腐病的发生伴随着土壤Fusarium丰度的快速增加。如Dong等[34]研究发现,三七根腐病致病菌F. oxysporum丰度与三七死亡率呈显著正相关,Zhao等[35]研究发现太子参连作后,根腐病发病土壤中F. oxysporum丰度显著高于健康组。本研究发现,白及发病土壤中Fusarium丰度显著升高,可能是引起其根腐病的重要原因。此外,Erysiph丰度也显著升高,其已证实是引起多种植物白粉病的关键致病菌[36-37]。同样,土壤中还存在着许多具有良好生防潜力的有益微生物,如Hannaella coprosmaensis、Subramaniula cristata等可作为生防菌,抑制植物病原真菌的生长[38-39],Larkinella bovis具有促进植物生长的功能[40]。此外,生防菌在土壤中的相对丰度与植物发病率呈显著负相关[41],Liu等[42]研究发现,在西洋参连作土壤中施加有益微生物丛枝菌根真菌可显著降低F. oxysporum、F. solani等病原真菌的丰度,并有效促进西洋参生长。Zhang等[43]在三七根腐病株根部接种益生菌后,根际土壤中病原菌的相对丰度显著降低,并降低了三七的死亡率。在本研究中,白及根腐病植株根际土壤Microscypha、Hannaella等真菌,以及Akkermansia,IMCC26134,Larkinella等细菌的相对丰度显著降低,但其与白及根腐病的关联性尚不清楚。
本研究发现健康白及根际土壤脲酶、蛋白酶与蔗糖酶活性和有效钾含量较高,pH、有机质、铵态氮与硝态氮含量较低。已有研究表明,随着白及种植年限的增加,土壤pH升高,有机质含量增加,土壤脲酶和蔗糖酶活性降低[44],这与本研究结果相似,表明白及连作可能会导致土壤养分失衡,酶活性降低。土壤养分失衡会影响微生物群落组成,营养元素富集可促使病原微生物数量增加,从而导致植物病害率上升[33,45]。在本研究中,冗余分析表明根腐病组与健康组微生物群落结构具有明显分化特征,根腐病组具有更高的NH4+-N和NO3--N含量。研究表明,土壤中NH4+-N含量是影响植物根际微生物群落组成的重要因素,常与病原菌丰度呈正相关[46]。当土壤中氮含量较低时,小麦根腐病发病率显著降低,且与镰刀菌数量的减少密切相关[47]。Zhao等[48]研究发现,施用烟秆炭改善了白及连作土壤理化性质与微生物群落结构,促进了连作地白及的生长。因此,土壤养分失衡与酶活性降低也可能促进白及根腐病害的发生。本研究为理解白及根腐病发病机理并针对性地开展防治措施提供了科学依据。
4 结论
病原菌积累、微生物群落结构变化、土壤养分失衡与酶活性降低可能是导致白及根腐病害的主要原因,可采取有效措施抑制镰刀菌、白粉菌等致病菌群的生长,改良土壤理化环境与微生物群落结构,从而消减白及根腐病害。