大气CO2浓度倍增和高温对玉米气孔特征及气体交换参数的影响
2022-03-09张运鑫郝立华马保国常志杰殷嘉伟刘媛媛郑云普
刘 亮,张运鑫,郝立华,马保国,常志杰,殷嘉伟,刘媛媛,郑云普
大气CO2浓度倍增和高温对玉米气孔特征及气体交换参数的影响
刘 亮1,2,张运鑫1,郝立华1,马保国1,常志杰1,殷嘉伟3,刘媛媛1,郑云普1,2※
(1. 河北工程大学水利水电学院,邯郸 056038;2. 河北省水资源高效利用工程技术研究中心,邯郸 056038;3. 河北工程大学园林与生态工程学院,邯郸 056038)
为深入了解未来大气CO2浓度升高背景下玉米气孔特征及气体交换过程对高温的响应机理,该研究利用人工气候室,探究在大气CO2浓度400mol/mol(C400)和800mol/mol(C800)下,不同温度处理(昼/夜)25/19 ℃、31/25 ℃和37/31 ℃对玉米气孔特征及气体交换参数的影响机理。结果表明:1)CO2浓度升高对玉米气孔密度的影响并不显著(> 0.05),增温却导致玉米不同轴面气孔密度均显著增加(< 0.001);不同轴面气孔密度的增加幅度均随温度升高而增大,叶片气孔密度对环境温度升高的响应呈现出非线性变化趋势。2)将环境温度由25/19 ℃增加到37/31 ℃导致C400和C800处理下玉米蒸腾速率(r)分别提高57%和84%,且不同轴面的气孔密度均与r之间存在较好的线性相关关系(近轴面2=0.69;远轴面2=0.71)。3)当温度从25/19 ℃升高到31/25 ℃,2个CO2浓度处理下玉米的n分别提高23%和21%,但环境温度提高到37/31 ℃却导致n分别降低24%和13%,说明高温环境(37/31 ℃)对光合反应位点造成生理伤害,而高浓度CO2缓解了高温对玉米造成的生理胁迫。同时,37/31 ℃条件下玉米叶片光合系统II()最大光化学效率(v/m)显著降低的结果也直接支持了上述结论。研究结果有助于从气孔特征的角度深入了解CO2浓度和温度升高对玉米叶片气体交换过程产生的影响,为未来气候变化背景下实现农作物绿色高效提质增产提供理论依据。
玉米;蒸腾;增温;CO2富集;光合;气孔特征
0 引 言
自19世纪西方工业革命以来,全球大气CO2浓度急剧升高,有关研究预计到21世纪末大气CO2浓度将由当前的400mol/mol增加到800mol/mol,同时还将导致地球表面的平均气温提高约6.0 ℃[1]。因此,全球农业生产力和粮食安全正在面临CO2浓度升高和高温胁迫带来的双重挑战[2]。作为决定作物生长发育过程的关键环境因子,大气CO2浓度和温度不仅改变植物叶片气孔特征[3-4]和气体交换过程[5-6],还协同调控作物的产量及其籽粒品质[7]。鉴于此,大气CO2浓度和温度升高对农作物粮食产量和品质产生影响的机理研究在当前气候变化背景下日益受到学者们的普遍关注[4-5],故深入了解农作物对CO2浓度和温度升高的生理响应机制将为实现农作物绿色高效提质增产提供重要理论依据[6-7]。
气孔是植物与大气环境进行CO2和水蒸气交换的重要通道[8],故植物的气孔数量、气孔开度及其分布状况均影响叶片气体交换效率[8-9]。然而,植物气孔形态极易受环境温度[5]、大气CO2浓度[7]、空气湿度[10]、土壤水分[8]、紫外线[11]等影响,进而导致植物叶片气体交换效率的变化[12]。以往研究已证实,温度升高不仅对单个气孔形态特征产生影响,而且改变叶片表面气孔和表皮细胞的比例[5]。然而,目前有关气孔形态特征及其分布状况响应增温的研究结果仍存在较大差异,甚至得到截然相反的矛盾结论[8-9,13]。同时,农作物叶片不同轴面气孔形态特征及其分布状况对增温的响应也呈现不一致的变化趋势[3-4]。除温度外,大气CO2浓度也可以调控植物气孔的生长发育过程[14-15]。基于早期保存植物标本获取的长时间序列气孔数据以及利用气候室CO2富集试验结果均表明,大气CO2浓度升高也可以改变许多植物物种的气孔形态特征[4,12,14-16]。此外,大气CO2浓度和温度的升高还影响植物叶片的气体交换过程[3]。通常而言,温度可以调控植物代谢过程关键酶类的活性,从而对作物的光合与呼吸过程产生影响[3];同时,CO2分子作为植物光合碳同化过程的基本原料之一,也直接决定着作物的光合同化效率[12-13]。尽管未来气候变化情形下大气CO2浓度倍增势必导致全球气温升高,但当前大部分研究仍主要集中在作物气孔特征和气体交换过程对单一环境因素的响应[17-20],而这并不能全面真实地反映未来气候变化对农作物生长发育过程的影响[5]。
众所周知,玉米(L.)是一种在全世界广泛分布的C4物种,也是中国最重要的粮食作物之一[5]。未来大气CO2浓度升高和全球变暖势必改变玉米的光合性能、蒸腾速率以及水分利用效率,并进一步影响玉米产量[7]。然而,目前有关CO2浓度升高和增温协同影响玉米气孔特征和气体交换过程的潜在机理尚不清楚[5,7],尤其对于大气CO2浓度和气温升高双重影响下玉米气孔特征与气体交换效率之间关系的理解还存在较大的分歧[13,16]。因此,将叶片气孔特征变化与气体交换过程有机结合有助于准确了解气候变化背景下CO2浓度升高和增温影响玉米产量的潜在机理。本文利用可精准控制CO2浓度和温度的大型气候室,研究玉米气孔特征及气体交换参数对CO2浓度和温度升高的响应机制,进一步分析玉米气孔特征与净光合速率和蒸腾速率之间的关系,以期从气孔特征变化的角度深入了解大气CO2浓度和温度升高协同影响玉米气体交换过程的潜在机理,为未来气候变化背景下实现农作物绿色高效提质增产提供重要的理论依据。
1 材料与方法
1.1 试验材料与设计
供试玉米品种郑单958(L. cv. Zhengdan 958),采用6个大型人工气候室(高1.8 m×宽1.8 m× 深0.67 m)进行盆栽试验,基质采用壤土混合营养土配制(体积比3∶1)。本研究将6个气候室均分为2组,其中CO2浓度分别设置为400mol/mol(C400)和800mol/mol(C800)。以往研究表明玉米生长的最适温度范围处于25~34 ℃[6,13],为了进一步明确玉米生长的最适温度以及高温胁迫对玉米生长的影响机理,将每组中3个气候室的温度(昼/夜)分别设置为对照(25/19 ℃)、中温(31/25 ℃)、高温(37/31 ℃)。盆栽玉米经播种、发芽、间苗后每盆保留一株,移入气候室进行培养。每个气候室中放置4盆,气候室内的光强1 000mol/(m2s)、光照周期12 h(07:00-19:00)、相对空气湿度50%~60%。60 d处理期内,所有盆栽玉米每周浇1次,每次浇灌2.0 L自来水,每次均有水分从盆底小孔溢出。同时,每周使用Hoagland溶液浇灌1次,每次400 mL。为避免气候室自身差异性对研究结果的影响,在整个处理期间,盆栽玉米每隔10 d轮换1次气候室,以尽量消除气候室本身给试验结果带来的系统误差。
1.2 测定指标及方法
1.2.1 气体交换参数和叶绿素荧光参数的测量
从玉米植株顶端向下选取第1片完全展开的成熟叶片,利用便携式光合测定系统(LI-6400XT,LI-COR Inc.,Lincoln,NE,USA)测量玉米叶片气体交换参数,包括净光合速率(n)、蒸腾速率(r)、气孔导度(g)等。测量气体交换参数时,将叶室内光强设置为1 000mol/(m2×s)、CO2浓度设定为气候室内相对应的C400和C800、叶室测量温度设置为玉米生长过程中气候室内相对应的环境温度。此外,利用叶绿素荧光测定仪(Handy PEA,Hansatech Instrument Ltd.,UK)测量玉米叶片光合系统II(PSII)的最大光化学效率(v/m)。
1.2.2 气孔特征参数测量
利用透明指甲油采集玉米叶片表面的气孔印迹,在配备有照相机的光学显微镜下对气孔印迹进行拍照,并测量和计算玉米气孔长度、气孔宽度、气孔面积、气孔周长、气孔指数及气孔面积指数[13]。
1.2.3 气孔特征的观察及拍照
首先,将采集的玉米叶片样品经磷酸缓冲液冲洗后,用1% (v/v) 锇酸固定。然后,将叶片组织经一系列梯度酒精脱水和干燥后进行镀膜处理。最后,利用扫描电子显微镜(XL30-FEG,FEI,Eindhoven,Netherlands)观察气孔特征并拍照。
1.2.4 叶片形态参数的测量
选取用于测定气体交换参数的叶片,测量该叶片的长度和宽度,同时利用叶面积系数法计算叶片面积[21]。
1.3 数据分析
本研究采用单因素和双因素方差分析法比较不同CO2浓度和温度水平对玉米气孔特征、气体交换参数及叶片参数的影响及其交互效应,而后利用S-N-K(Student-Newman-Keuls)法进行多重比较,分析不同温度和CO2浓度处理间的差异。
2 结果与分析
2.1 大气CO2浓度倍增和高温对玉米气孔形态参数的影响
环境温度升高导致玉米叶片表面气孔个数(气孔密度)显著增多,但高CO2浓度并没有显著改变玉米气孔密度(表1和表2)。将玉米生长温度由25/19 ℃升高至31/25 ℃时,当前大气CO2浓度下(C400)玉米气孔密度分别提高约15%(近轴面)和12%(远轴面);同时升高CO2浓度(C800)处理下玉米叶片气孔密度增加约15%(近轴面)和8%(远轴面)。相似地,继续将环境温度由31/25 ℃提高到37/31 ℃后,C400环境下玉米气孔密度分别增加约28%(近轴面)和25%(远轴面),且C800处理下的气孔密度提高约28%(近轴面)和26%(远轴面)。高温导致2个CO2浓度下玉米不同轴面气孔密度均有所增加,且近轴面气孔密度提高的幅度却明显高于远轴面。此外,玉米叶片不同轴面气孔密度增加幅度随环境温度的不断升高而逐渐增大。利用扫描电子显微镜对玉米叶片表面气孔进行直接观察的结果也证实了上述结论(图1)。相对于25/19 ℃而言,环境温度升至31/25 ℃,C400环境下玉米不同轴面的气孔宽度分别降低4.2%(近轴面)和6.4%(远轴面);同时C800环境下,增温也导致玉米叶片气孔宽度显著降低4.4%(近轴面)和4.3%(远轴面)。将温度由31/25 ℃提高到37/31 ℃后,C400环境下玉米的不同轴面的气孔宽度降低约8.9%(近轴面)和6.8%(远轴面),且C800处理下气孔宽度也下降13.9%(近轴面)和13.6%(远轴面)。该研究结果表明玉米叶片气孔宽度随温度升高而不断减小。此外,双因素方差分析结果还表明CO2浓度和温度对玉米近轴面气孔宽度产生显著的交互作用(< 0.05)。
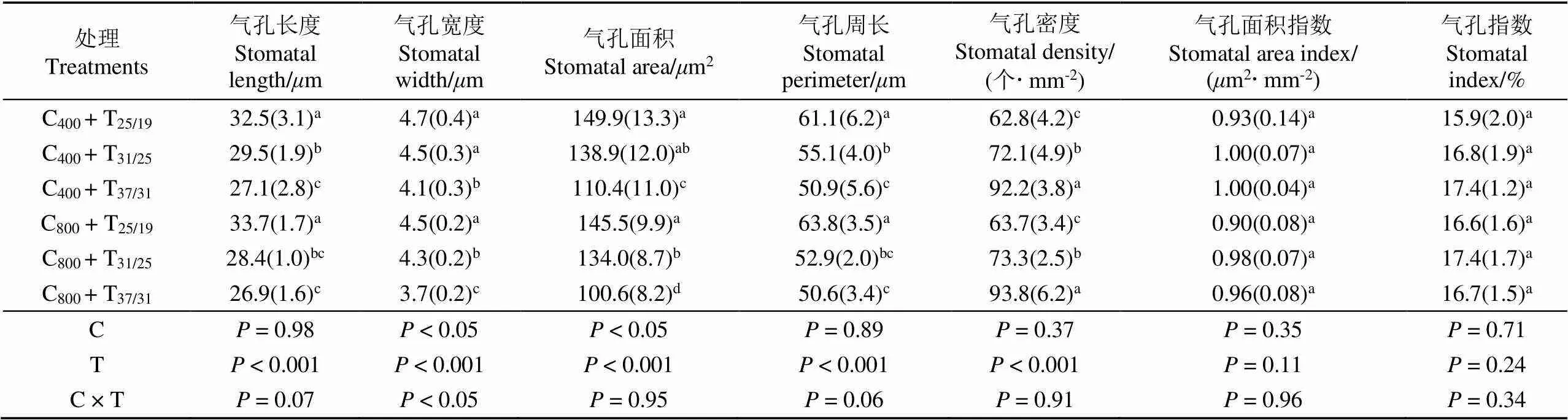
表1 不同处理下玉米叶片近轴面气孔特征参数
注:表中所有数据表示为平均值(标准偏差)。表中字母C代表CO2,其下标为CO2浓度值;T代表温度,下标表示昼/夜温度。不同小写字母表示各个处理间差异显著(< 0.05),下同。
Note: Values given are means (standard deviation). Letter C represents CO2, and its subscript represents the value of CO2concentration; letter T represents temperature, and its subscript represents day/night temperature. Different lowercase letters indicate significant difference among different treatments at the 0.05 level. The same below.

表2 不同处理下玉米叶片远轴面气孔特征参数
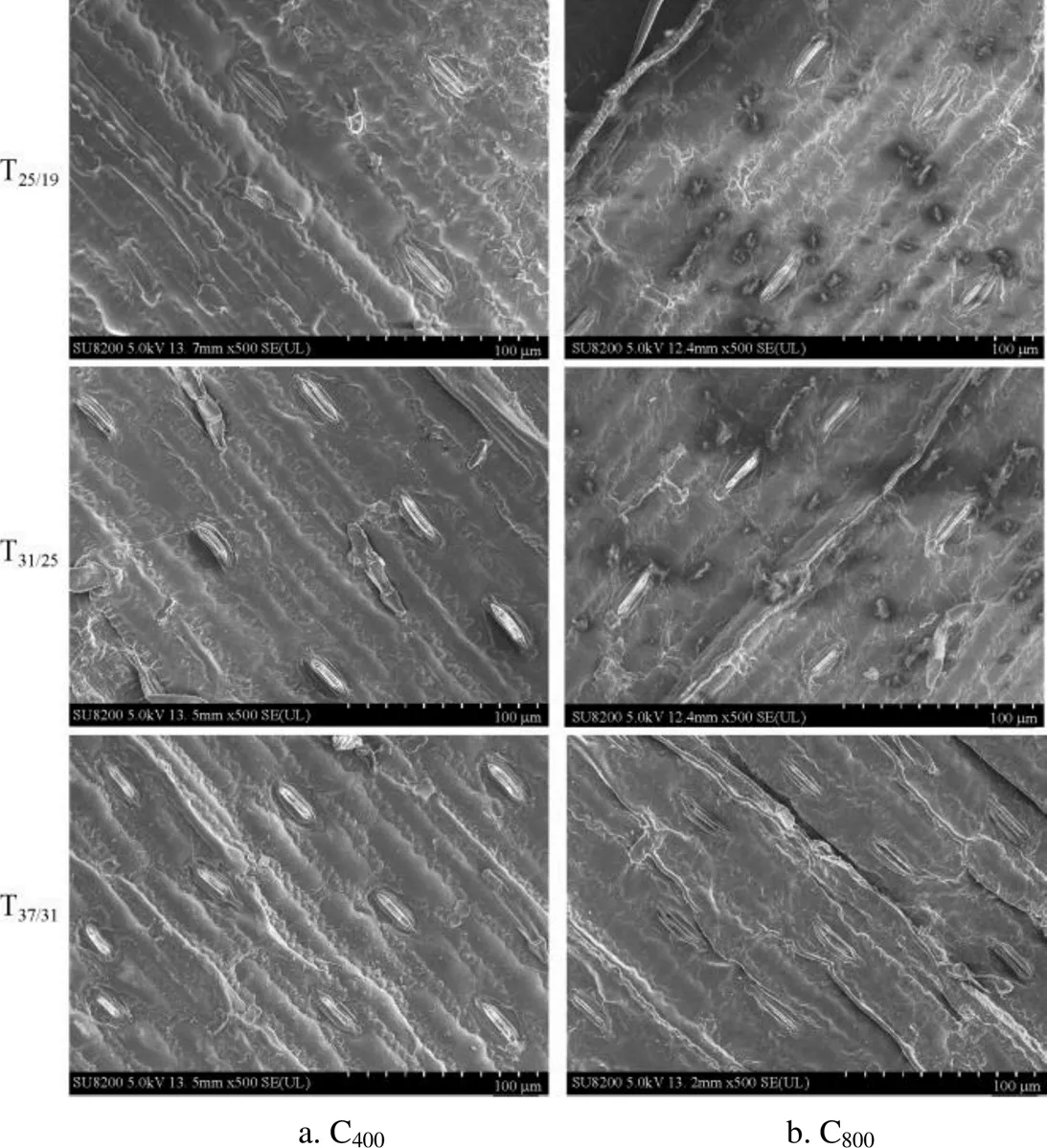
图1 不同大气CO2浓度和温度下玉米叶片气孔电镜扫描图
2.2 大气CO2浓度倍增和高温对玉米气体交换和叶绿素荧光参数的影响
玉米净光合反应速率(n)随环境温度升高呈先增加后降低的变化趋势。C400与C800二氧化碳浓度处理下,当温度从25/19 ℃升高到31/25 ℃,玉米n分别提高约23%和21%,但继续提高温度到37/31 ℃却导致叶片n分别降低约24%和13%(表3)。此外,研究结果还表明,CO2浓度升高对n的影响在不同温度处理下产生显著差异(表3)。在25/19 ℃和31/25 ℃处理下CO2浓度升高对n造成的影响并不显著,但高温(37/31 ℃)处理下CO2浓度升高却导致n显著提高约16%(表3);同时,在高温(37/31 ℃)处理下玉米叶片PSII的最大光化学效率(v/m)较中温处理(31/25 ℃)也显著降低。此外,在高温处理下(37/31 ℃),CO2浓度富集提高v/m约10%(表3),表明当玉米处于极端高温环境时,大气CO2浓度升高可以在一定程度上缓解高温对玉米光合反应位点造成的生理损伤。另外,不同CO2浓度下玉米蒸腾速率(r)均随着环境温度的升高而逐渐增加,当温度由25/19 ℃增加到37/31 ℃分别导致C400和C800处理下r显著提高约57%和84%(表3)。然而,大气CO2浓度升高导致不同温度处理下(25/19 ℃、31/25 ℃和37/31 ℃)玉米的r降低约22%、13%和10%(表3);同时,高CO2浓度还导致3个温度处理下玉米叶片的s分别下降约19%、7%和8%。此外,CO2浓度和温度对玉米的n、s和v/m均产生显著的交互效应(表3)。

表3 不同处理下玉米叶片气体交换参数
2.3 大气CO2浓度倍增和高温对玉米叶片形态参数的影响
图2为大气CO2浓度倍增和高温条件下玉米叶片形态参数的变化,由图2可知不同温度处理下玉米叶片长度、宽度和面积均存在显著差异,高温(37/31 ℃)处理导致C400和C800条件下玉米叶片长度、宽度及叶面积相比25/19 ℃均降低约50%,表明在极端高温条件下,不同CO2浓度环境下的玉米叶片均变得更小,说明高温可能限制了玉米叶片的纵向生长和横向伸展。不同环境温度下玉米叶片长度、宽度和叶面积在2个CO2浓度之间并不存在显著差异。此外,尽管高温改变了玉米叶片形态特征,但却没有对单株玉米的叶片数量产生显著影响,表明玉米植株主要通过调整叶片的形态特征适应高温环境。
2.4 大气CO2浓度倍增和高温条件下玉米气孔特征与Pn及Tr的相关性分析
玉米叶片近轴面和远轴面的气孔密度与n在25/19 ℃和31/25 ℃两个温度处理下均具有较好的相关关系(图3),表明25/19 ℃和31/25 ℃温度下玉米的n主要受气孔因素的限制。然而,在高温(37/31 ℃)处理下玉米叶片两个轴面气孔密度与n的相关性却明显降低,表明在37/31 ℃处理下玉米的光合反应位点可能受到了高温伤害,故高温环境n的降低可能主要受非气孔因素的限制。进一步深入分析发现,相较于25/19 ℃和31/25 ℃2个温度处理,37/31 ℃处理下玉米叶片的最大光化学效率(v/m)也显著降低(< 0.05;表3),证实了极端高温下玉米的光系统遭受到高温伤害。此外,本研究还发现玉米的气孔密度随着温度升高而同向增加(表1和表2),但n却随着温度升高呈现先增加后降低的变化趋势(表3),表明气孔密度与n随温度升高的变化并不一致,也证实在高温(37/31 ℃)下玉米n的下降主要受到非气孔因素限制。此外,玉米叶片气孔密度、气孔面积指数与蒸腾速率之间存在较好的正相关关系(图4),表明气孔密度和气孔面积指数的增加均有助于提升叶片蒸腾。玉米叶片气孔数量增加可以缩短叶片内水分输送到气孔附近蒸腾位点的距离,减小水分在叶片中的传输阻力,气孔面积指数的增加提高了叶片与大气进行水蒸气交换的面积,均有助于提高玉米蒸腾速率。
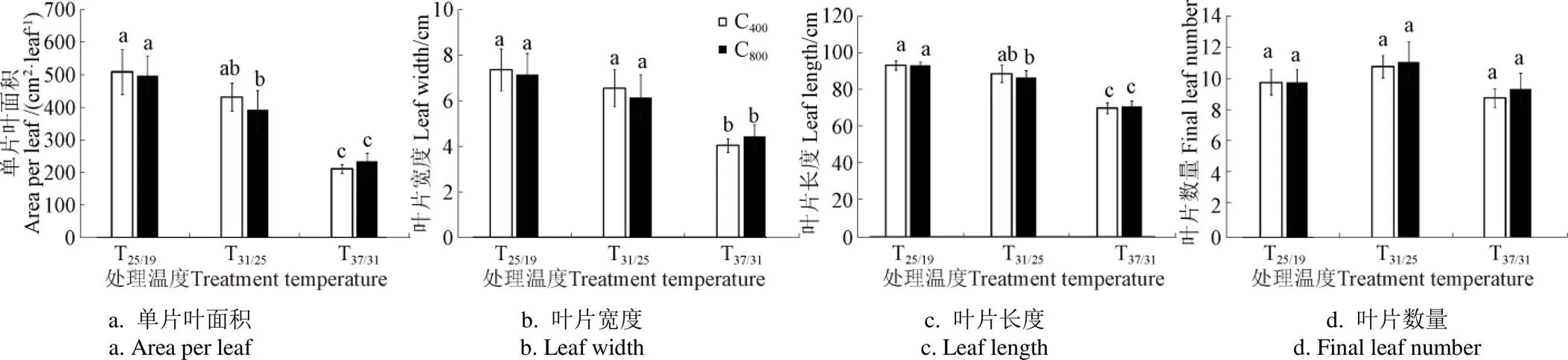
图2 大气CO2浓度和温度升高对玉米叶片形态特征的影响
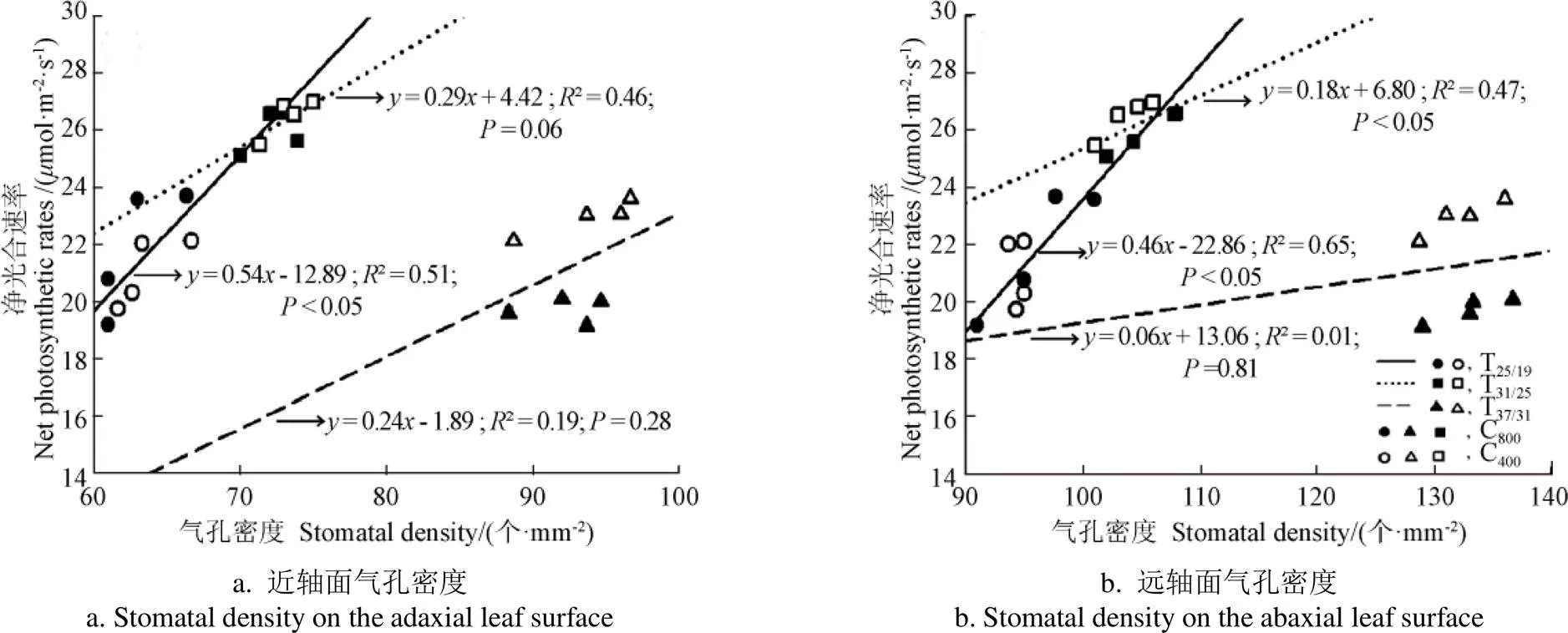
图3 玉米净光合速率与气孔密度的关系
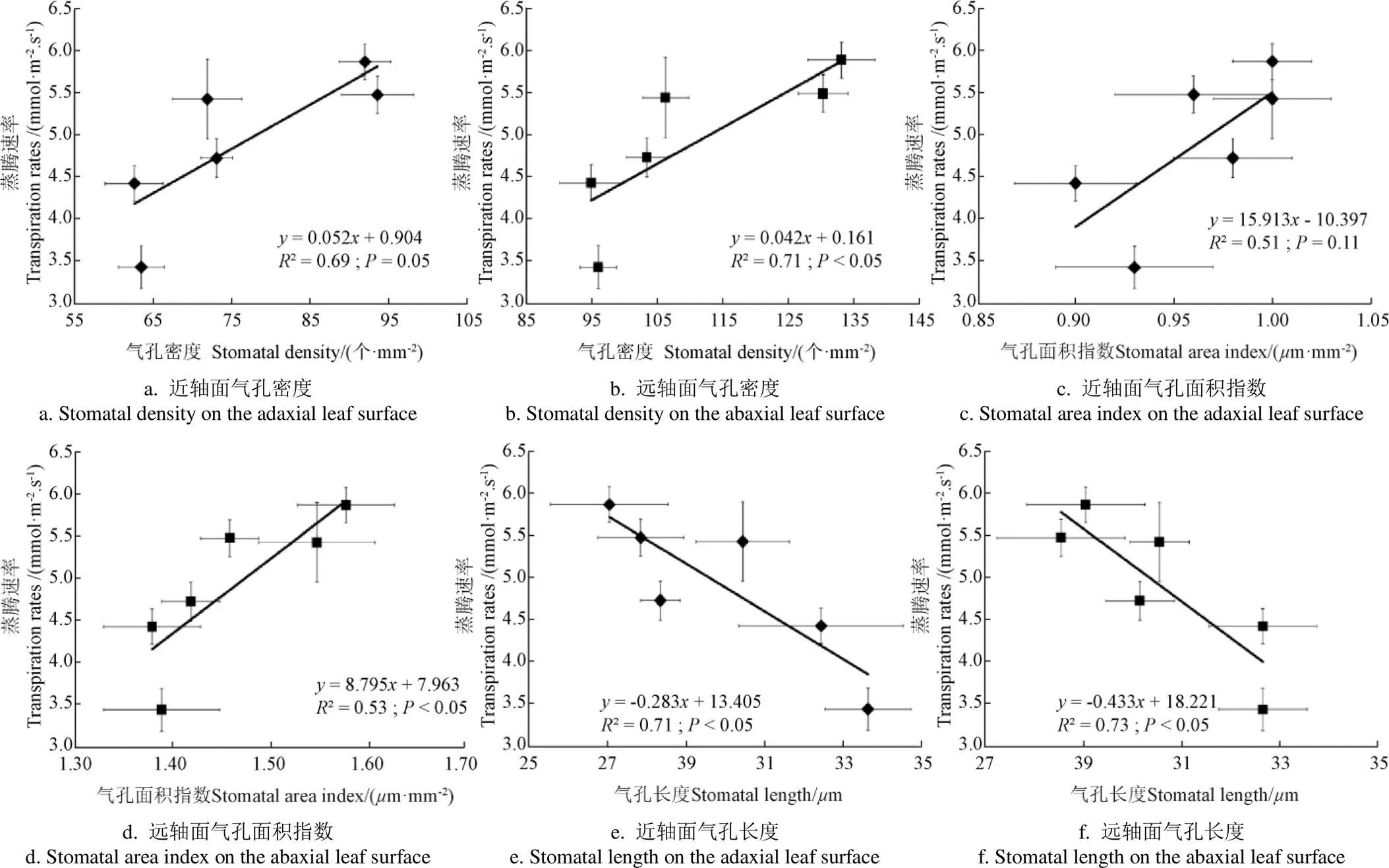
图4 玉米蒸腾速率与气孔特征的关系
3 讨 论
3.1 高温对玉米气孔密度的影响
目前,有关增温对植物气孔密度产生影响的结论还存在分歧。有学者发现增温对叶片气孔密度无显著影响[22],而另有研究表明气孔密度会随温度的升高而减少[9]或增加[4]。本研究发现玉米气孔密度随温度升高而逐渐增加,并且气孔密度与温度之间存在非线性关系,即随着环境温度的不断升高,玉米气孔密度增加幅度也相应变大。因此,小幅度增温对植物气孔密度的影响可能并不明显,而较大的增温幅度才能充分体现植物气孔特征对温度变化的响应规律[4,13],这可能是导致在不同增温试验中研究结果不一致的主要原因[4,9,13,22-23]。值得注意的是,不同植物功能组或物种的气孔对增温敏感性差异也是导致上述矛盾结论的重要原因之一[9]。此外,植物气孔特征还受物种自身遗传因素的控制,乔孟等[24]在拟南芥中发现2个类转录因子和共同调控气孔发育过程中保卫细胞的分化过程;而环境变化能够引起植物细胞在分裂中通过对转录过程的精确调控,进一步改变基因的表达量以适应环境变化[23]。本研究结果表明,玉米叶片气孔密度不仅随着环境温度升高而显著增加,并且叶片近轴面的气孔密度对增温的响应相比远轴面更加敏感,说明玉米叶片两个轴面的气孔数量可能受到了基因的调控,并且增温对气孔控制基因表达量的影响在不同轴面之间存在差异。同时,近期的研究结果还发现,增温会导致作物生育期缩短[5],表皮细胞变小[22],进而增加叶片气孔密度[3]。本研究结果也表明,增温可以减小玉米叶片表皮细胞的伸展长度,进而导致更短和窄的叶片,这也可能是导致玉米叶片气孔密度增加的原因。更进一步讲,本研究中增温导致玉米气孔密度增加可以在较大程度上提高单位叶面积的蒸腾效率,有助于玉米叶片更高效的散失热量,从而适应高温环境[3]。
3.2 大气CO2浓度升高对玉米气孔特征的影响
目前有关大气CO2浓度升高影响农作物的研究主要集中在叶片光合速率[5]、水分利用效率[5]和粮食产量[7]等方面,而未来CO2浓度升高条件下植物通过调整气孔形态特征优化叶片气体交换效率的机理仍不清楚[8,15,25]。本研究将大气CO2浓度由C400提高到C800并没有显著改变玉米的气孔密度和气孔指数,这可能是由于植物叶片表面分布气孔的数量及其比例对CO2浓度升高的响应在较低的CO2浓度环境下才更加显著[26]。另外,植物叶片气孔特征对CO2浓度的响应还同其自身的光合类型有关[16,26],通常C3植物的生理参数及叶片结构特征对CO2浓度的升高表现的更加敏感,而C4植物对CO2浓度的响应并不敏感[26]。本研究中高CO2浓度条件下玉米气孔密度和气孔指数均没有产生明显变化也可能是由于玉米的C4特性而造成的。大气CO2浓度升高调节气孔特征的机理目前还不清楚,但已有的研究结果表明,CO2浓度升高可以调控植物气孔相关编码基因的表达量,从而影响气孔的发育过程[27]。此外,植物叶片的气体交换效率还与气孔的开张程度紧密联系[12]。尽管植物气孔密度和气孔指数主要受长期的环境信号及其遗传进化过程影响,但气孔开度却可以快速响应外界环境的短期变化。以往的相关研究结果表明,光照[28]、CO2浓度[19]、温度[3]及土壤水分[8]等许多环境因子的短期变化均会对植物气孔的开度产生影响。本研究结果也显示,大气CO2浓度升高导致玉米叶片气孔开口面积的减小,可能是由于环境CO2浓度升高造成细胞间CO2浓度的增加,故玉米通过减小气孔开度来限制更多的CO2分子通过气孔从外界大气扩散到叶肉内部并同时降低水分的蒸腾,从而获得更优的碳-水分配策略。
3.3 大气CO2浓度升高条件下高温对玉米气孔特征和气体交换效率的影响
本研究中的高温处理增加了不同CO2浓度条件下玉米的气孔的密度和叶片的蒸腾速率,且气孔密度与蒸腾速率之间存在较好的线性相关关系,表明高温可以导致气孔密度的增加,从而提高叶片的蒸腾效率。然而,玉米的气孔面积指数却没有随温度的升高而增加,表明植物通过气孔散失水分的效率并不与气孔面积呈正相关关系,而是同气孔密度和气孔周长成正比[3,29]。另外,当温度由25/19 ℃升高至31/25 ℃后,玉米的气孔密度显著增加,且同时伴随气孔导度和光合速率的提高,表明增温可以使玉米叶片更高效的从大气获取CO2分子。此外,在25/19 ℃和31/25 ℃温度处理下,玉米气孔密度与n均呈现出良好的线性关系(图3),也证实此时玉光合主要受到气孔的限制。然而,温度由31/25 ℃增至37/31 ℃,玉米气孔密度仍显著增加,但n却急剧降低,表明此时光合可能受到了由高温胁迫引起的非气孔因素限制。本研究中玉米叶片PSII最大光化学效率(v/m)在高温下显著降低的结果也直接支持了上述结论[30]。
已有研究发现在环境胁迫条件下,植物叶片可通过气孔开闭对逆境做出相应的反馈[31],调节叶片细胞水分散失和光合效率间的平衡。故此,气孔开度也是反映植物抗逆性的重要指标之一。本研究结果表明,极端高温(37/31 ℃)条件下,大气CO2浓度升高可以显著增加玉米的n,表明高浓度CO2可以在一定程度上缓解极端高温对玉米产生的生理胁迫。笔者认为,大气CO2浓度升高缓解玉米高温伤害的机理可能与CO2诱导玉米气孔开度减小有关,这主要是由于植物气孔开度、叶片温度以及叶片内水分状况之间均存在着复杂的关系[32],故究其原因可能是高温环境下植物的叶片温度升高,从而导致参与光合过程的关键酶类在高温条件下活性降低,甚至彻底失活[3];植物为了在极大程度上避免高温伤害,通过增大气孔的开张程度提高叶片的蒸腾速率,从而降低叶片温度来适应高温环境[10,32-33],但这同时会带来叶片组织过度失水的风险。然而,大气CO2浓度升高不仅诱导气孔开度减小以降低叶片蒸腾,还可以增加叶片的n,从而提高玉米叶片水平的水分利用效率[5]。本研究也发现,极端高温环境(37/31 ℃)导致玉米叶片的蒸腾速率提高,而大气CO2浓度升高可以诱导玉米气孔的关闭,从而减少气孔开度和叶片蒸腾,进而平衡玉米叶片热量散失和细胞水分之间的关系[10],最终提高玉米抵抗高温胁迫的能力[33]。类似地,Zhang等[10]对番茄幼苗的研究也发现,高温胁迫环境下大气CO2浓度升高可以减小气孔的开张程度,甚至关闭气孔,进一步降低叶片的蒸腾速率以防止番茄叶片中水分过度流失,从而提高番茄的抵抗热胁迫的能力。
4 结 论
本研究深入探讨大气CO2浓度倍增和高温对玉米叶片气孔特征及其气体交换效率的影响机理,得到以下主要结论:
1)环境温度升高导致400和800mol/mol的二氧化碳浓度下玉米不同轴面的气孔密度均明显增加,且两个轴面气孔密度对温度升高的响应均呈现非线性的变化趋势,但玉米叶片近轴面气孔密度对增温的响应相比远轴面更加敏感。
2)当昼/夜温度由25/19 ℃提高到31/25 ℃时,不同CO2浓度下玉米叶片n主要受气孔因素限制,而玉米可以在一定程度上通过调整气孔密度优化叶片的气体交换效率,从而提高叶片净光合速率和蒸腾速率。
3)当温度继续升高到37/31 ℃时,玉米的n主要受非气孔因素限制,而大气CO2浓度升高能够减小气孔开度和叶片蒸腾,从而维持叶片热量散失与叶肉细胞水分含量之间的平衡关系,提高叶片抵抗高温伤害的能力,缓解高温对玉米造成的生理胁迫。
[1] Inergovernmental Panel on Climate Change. “Summary for Policymakers, ” in Climate Change and Land: An IPCC Special Report on Climate Change, Desertifcation, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fuxes in Terrestrial Ecosystems[R]. Geneva: IPCC, 2019.
[2] Wang B, Cai W, Li J, et al. Leaf photosynthesis and stomatal conductance acclimate to elevated [CO2] and temperature thus increasing dry matter productivity in a double rice cropping system[J]. Field Crops Research, 2020, 248: 107735.
[3] Hao L, Guo L, Li R, et al. Responses of photosynthesis to high temperature stress associated with changes in leaf structure and biochemistry of blueberry (L.)[J]. Scientia Horticulturae, 2019, 246: 251-264.
[4] 张茜茜,王海峰. 大气 CO2浓度倍增和增温对冬小麦气孔特征的协同效应[J]. 东北农业科学,2021,46(1):25-30.
Zhang Xixi, Wang Haifeng. Interactive effects of doubling CO2concentration and experimental warming on the stomatal traits of winter wheat[J]. Journal of Northeast Agricultural Sciences, 2021, 46(1): 25-30. (in Chinese with English abstract)
[5] 刘亮,郝立华,李菲,等. CO2浓度和温度对玉米光合性能及水分利用效率的影响[J]. 农业工程学报,2020,36(5):122-129.
Liu Liang, Hao Lihua, Li Fei, et al. Effects of CO2concentration and temperature on leaf photosynthesis and water efficiency in maize[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2020, 36(5): 122-129. (in Chinese with English abstract)
[6] Kim S H, Gitz D C, Sicher R C, et al. Temperature dependence of growth, development, and photosynthesis in maize under elevated CO2[J]. Environmental and Experimental Botany, 2007, 61(3): 224-236.
[7] Abebe A, Pathak H, Singh S D, et al. Growth, yield and quality of maize with elevated atmospheric carbon dioxide and temperature in north-west India[J]. Agriculture Ecosystems & Environment, 2016, 218: 66-72.
[8] Fan X, Cao X, Zhou H, et al. Carbon dioxide fertilization effect on plant growth under soil water stress associates with changes in stomatal traits, leaf photosynthesis, and foliar nitrogen of bell pepper (L. )[J]. Environmental and Experimental Botany, 2020, 179: 104203.
[9] 左闻韵,贺金生,韩梅,等. 植物气孔对大气CO2浓度和温度升高的反应—基于在CO2浓度和温度梯度中生长的10种植物的观测[J]. 生态学报,2005,25(3):565-574.
Zuo Wenyun, He Jinsheng, Han Mei, et al. Responses of plant stomata to elevated CO2and temperature: observations from 10 plant species grown in temperature and CO2gradients[J]. Acta Ecologica Sinica, 2005, 25(3): 565-574. (in Chinese with English abstract)
[10] Zhang H, Pan C, Gu S, et al. Stomatal movements are involved in elevated CO2-mitigated high temperature stress in tomato[J]. Physiologia Plantarum, 2018, 165(3): 569-583.
[11] Yang J, Li C, Kong D, et al. Light-mediated signaling and metabolic changes coordinate stomatal opening and closure[J]. Frontiers in Plant Science, 2020, 11: 601478.
[12] Xu M. The optimal atmospheric CO2concentration for the growth of winter wheat[J]. Journal of Plant Physiology, 2015, 184: 89-97.
[13] 郑云普,徐明,王建书,等. 玉米叶片气孔特征及气体交换过程对气候变暖的响应[J]. 作物学报,2015,41(4):601-612.
Zheng Yunpu, Xu Ming, Wang Jianshu, et al. Responses of the stomatal traits and gas exchange of maize leaves to climate warming[J]. Acta Agronomica Sinica, 2015, 41(4): 601-612. (in Chinese with English abstract)
[14] Heath O V S. Control of stomatal movement by reduction in the normal carbon dioxide content of the air[J]. Nature, 1948, 161: 179-181.
[15] Beerling D J, Royer D L. Reading a CO2signal from fossil stomata[J]. New Phytologist, 2002, 153(3): 387-397.
[16] Driscoll S P, Prins A, Olmos E, et al. Specification of adaxial and abaxial stomata, epidermal structure and photosynthesis to CO2enrichment in maize leaves[J]. Journal of Experimental Botany, 2005, 57: 381-390.
[17] 苏营,张逸飞,牟文雅,等. 大豆主要株型和产量指标对大气CO2和温度升高的响应[J]. 生态学报,2016,36(9):2597-2606.
Su Ying, Zhang Yifei, Mou Wenya, et al. Morphological traits and yield of soybean under elevated atmospheric CO2concentration and temperature[J]. Acta Ecologica Sinica, 2016, 36(9): 2597-2606. (in Chinese with English abstract)
[18] Wang D R, Bunce J A, Tomecek M B, et al. Evidence for divergence of response in Indica, Japonica, and wild rice to high CO2× temperature interaction[J]. Global Change Biology, 2016, 22(7): 2620-2632.
[19] 李菲,刘亮,张浩,等. CO2浓度对大豆叶片气孔特征和气体交换参数的影响[J]. 作物学报,2018,44(8):1212-1220.
Li Fei, Liu Liang, Zhang Hao, et al. Effects of CO2concentrations on stomatal traits and gas exchange in leaves of soybean[J]. Acta Agronomica Sinica, 2018, 44(8): 1212-1220. (in Chinese with English abstract)
[20] Sage R F. Variation in thecatof Rubisco in C3and C4plants and some implications for photosynthetic performance at high and low temperature[J]. Journal of Experimental Botany, 2002, 53: 609-620.
[21] 杨国虎,李建生,罗湘宁,等. 干旱条件下玉米叶面积变化及地上干物质积累与分配的研究[J]. 西北农林科技大学学报(自然科学版),2005,33(5):27-32.
Yang Guohu, Li Jiansheng, Luo Xiangning, et al. Studies on leaf area change and above-ground dry material accumulation and distribution of maize in different droughts[J]. Journal of Northwest A&F University(Natural Science Edition), 2005, 33(5): 27-32. (in Chinese with English abstract)
[22] Zheng Y P, Xu M, Hou R X, et al. Effects of experimental warming on stomatal traits in leaves of maize (L. )[J]. Ecology and Evolution, 2013, 3(9): 3095–3111.
[23] Apple M E, Olszyk D M, Ormrod D P, et al. Morphology and stomatal function of douglas fir needles exposed to climate change: elevated CO2and temperature[J]. International Journal of Plant Sciences, 2000, 161(1): 127-132.
[24] 乔孟,于延冲,向凤宁. 拟南芥类转录因子在环境胁迫中的作用[J]. 生命科学,2009,21(1):145-150.
Qiao Meng, Yu Yanchong, Xiang Fengning. The roles of thetranscription factors in the stress responses[J]. Chinese Bulletin of Life Sciences, 2009, 21(1): 145-150. (in Chinese with English abstract)
[25] Royer D L. Stomatal density and stomatal index as indicators of paleoatmospheric CO2concentration[J]. Review of Palaeobotany and Palynology, 2001, 114(1-2): 1-28.
[26] Bettarini I, Vaccari F P, Miglietta F. Elevated CO2concentrations and stomatal density: Observations from 17 plant species growing in a CO2spring in central Italy[J]. Global Change Biology, 1998, 4(1): 17-22.
[27] Gray J E, Holroyd G H, Van Der Lee F M, et al. The HIC signalling pathway links CO2perception to stomatal development[J]. Nature, 2000, 408: 713-716.
[28] 陈斌,刘筱玮,贾琳,等. 光强对4种鸭跖草科植物生长和光合特性的影响[J]. 生态学报,2022,42(4):1450-1461.
Chen Bin, Liu Xiaowei, Jia Lin, et al. Effects of light intensity on the growth and photosynthetic characteristics of fourplants[J]. Acta Ecologica Sinica, 2022, 42(4): 1450-1461. (in Chinese with English abstract)
[29] 武维华. 植物生理学[M]. 北京:科学出版社,2018.
[30] Janka E, Körner O, Rosenqvist E, et al. Using the quantum yields of photosystem II and the rate of net photosynthesis to monitor high irradiance and temperature stress in chrysanthemum ()[J]. Plant Physiology and Biochemistry, 2015, 90: 14-22.
[31] 杨爱峥,李志磊,付强,等. CO2浓度倍增和土壤盐胁迫对藜麦生理特征及产量的影响[J]. 农业工程学报,2021,37(4):181-187.
Yang Aizheng, Li Zhilei, Fu Qiang, et al. Effects of elevated atmospheric CO2on physiological characteristics and yield of quinoa to salinity stress[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2021, 37(4): 181-187. (in Chinese with English abstract)
[32] Kumar A, Omae H, Egawa Y, et al. Influence of water and high temperature stress on leaf water status of high temperature tolerant and sensitive cultivars of snap bean (L. )[J]. Japanese Journal of Tropical Agriculture, 2005, 49: 109-118.
[33] 郑云普,常志杰,范晓懂,等. CO2浓度升高和磷素亏缺对黑麦草气孔特征及气体交换参数的影响[J]. 农业工程学报,2021,37(18):82-89.
Zheng Yunpu, Chang Zhijie, Fan Xiaodong, et al. Effects of CO2concentration increase and phosphorus deficiency on the stomatal traits and leaf gas exchange parameters of ryegrasse[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2021, 37(18): 82-89. (in Chinese with English abstract)
Effects of double atmospheric CO2concentration and high temperature on the stomatal traits and leaf gas exchange of maize plants
Liu Liang1,2, Zhang Yunxin1, Hao Lihua1, Ma Baoguo1, Chang Zhijie1, Yin Jiawei3, Liu Yuanyuan1, Zheng Yunpu1,2※
(1.,,056038,;2.,056038,;3.,,056038,)
To further understand the response mechanism of stomatal traits and leaf gas exchange of maize (L.) to elevated CO2concentration and high temperature stress, we examined the combined effects of double atmospheric CO2concentration and high temperature on plant growth, stomatal traits, and leaf gas exchange parameters of maize grown at six environmental growth chambers with three temperature regimes ((day/night) 25/19 ℃, 31/25 ℃, and 37/31 ℃) and two CO2concentrations (400mol/mol (C400) and 800mol/mol (C800)), respectively. These environmental growth chambers were controlled with the same environmental factors, where the Photosynthetic Photon Flux Density (PPFD) was 1 000mol/(m2·s) and the relative humidity was 50% to 60%. In each chamber, we measured the net photosynthetic rate (n), transpiration rate (r)and stomatal conductance (s) using a portable photosynthesis system (LI-6400XT, LI-COR Inc., Lincoln, NE, USA). The maximum photochemical efficiency of PSII (v/m) was estimated by measuring chlorophyll fluorescence with a photosynthesis efficiency analyzer (Handy PEA, Hansatech Instrument Ltd., Norfolk, UK). In addition, we also measured the leaf area, leaf length, and leaf width as well as the final leaf number of maize plants. The results showed that: 1) The stomatal density of maize was significantly increased by temperature (< 0.001), but barely affected by CO2concentration (> 0.05). Meanwhile, the increase of stomatal density on the adaxial leaf surface was significantly higher than that on the abaxial surface of maize leaves, which indicated that the response of stomatal density on the adaxial leaf surface to elevated temperature might be more sensitive than that on the abaxial leaf surface of maize. Furthermore, the results also showed that the increase of stomatal density was accelerated with the elevated temperature on both the adaxial and abaxial leaf surfaces of maize. 2)The leaf transpiration rate were significantly enhanced by 57% and 84% with increasing growth temperature(day/night) from 25/19 ℃ to 37/31 ℃at both the ambient (C400) and double atmospheric CO2concentrations (C800). And there were linear positive correlation between the stomatal density on the adaxial and abaxial leaf surfaces and the transpiration rate of maize plants (adaxial surface,2=0.69; abaxial surface,2=0.71), which indicated that the leaf transpiration rate could be improved by adjusting stomatal density to optimize leaf gas exchange efficiency under high temperature environment.3) The net photosynthetic rate (n) of maize was also significantly enhanced by 23% and 21% with increasing temperature(day/night) from 25/19 ℃ to 31/25 ℃, however, thendrastically declined by 24% and 13% when the temperature increased from 31/25 ℃ to 37/31 ℃ at both CO2concentrations, which indicated that the high temperature (37/31 ℃) might result in physiological damages to the sites of photosynthetic reaction center, but this thermal stress from high temperature could be alleviated by elevated CO2concentration. Also, the maximum photochemical efficiency (v/m) of maize drastically decreased at both CO2concentrations when the temperature was elevated from 31/25 ℃ to 37/31 ℃. Overall, The results in this study maybe of significance for further understanding the potential mechanisms and processes of elevated atmospheric CO2concentration mitigating the physiological damage of high temperature to maize plants under future climate change.
maize; transpiration; elevated temperature; CO2enrichment; photosynthesis; stomatal traits
10.11975/j.issn.1002-6819.2022.22.008
S164.5
A
1002-6819(2022)-22-0073-08
刘亮,张运鑫,郝立华,等. 大气CO2浓度倍增和高温对玉米气孔特征及气体交换参数的影响[J]. 农业工程学报,2022,38(22):73-80.doi:10.11975/j.issn.1002-6819.2022.22.008 http://www.tcsae.org
Liu Liang, Zhang Yunxin, Hao Lihua, et al. Effects of double atmospheric CO2concentration and high temperature on the stomatal traits and leaf gas exchange of maize plants[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2022, 38(22): 73-80. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2022.22.008 http://www.tcsae.org
2022-09-23
2022-11-10
国家自然科学基金面上项目(32071608);中央引导地方科技发展资金项目(226Z6401G);河北省自然科学基金项目(E2021402031)
刘亮,博士生,讲师,研究方向为农业水资源高效利用与调控。Email:liuliang_12345@sina.com
郑云普,博士,教授,研究方向为农业水土工程及全球变化生态学。Email:zhengyunpu_000@sina.com
