菜子湖冬季水鸟空间分布与环境因子的关系研究
2022-03-04姚简周立志魏振华李凌晨
姚简, 周立志,*, 魏振华, 李凌晨
菜子湖冬季水鸟空间分布与环境因子的关系研究
姚简1,2, 周立志1,2,*, 魏振华1,2, 李凌晨1,2
1. 安徽大学资源与环境工程学院, 合肥 230601 2. 湿地生态保护与修复安徽省重点实验室(安徽大学), 合肥 230601
水鸟空间分布与环境因子的关系是生态研究的重要内容, 环境因子的变化可以通过改变栖息地的生境特征、食物资源从而直接或间接影响水鸟的种类、数量和分布。利用网格单元对2019—2020年菜子湖冬季水鸟及其环境因子进行调查, 共记录到水鸟6目12科41种, 其优势种为豆雁()和黑腹滨鹬()。运用空间自相关和多元逐步回归分析, 研究冬季水鸟的空间分布特征, 并确定影响不同取食集团水鸟分布的主要环境因子。网格鸟类占比显示, 菜子湖冬季水鸟主要分布在堂屋(30.98%)、梅花大圩(10.53%)、季庄(7.87%)、车富岛(6.49%)以及双兴村(5.67%); 食苔草水鸟与食块茎水鸟呈现显著的空间自相关, 聚集分布(<0.05,>0); 食鱼水鸟、食无脊椎水鸟和食种子水鸟为随机分布(>0.05)。环境因子对不同取食集团水鸟的影响也有所差异, 食鱼水鸟会倾向于选择大面积的开阔水域, 湿地裸露出的泥滩会吸引食无脊椎水鸟在此栖息, 沉水植被的退化使得食块茎水鸟聚集分布在农田和草滩中, 广阔草滩的形成使得食苔草水鸟数量明显增加, 食种子水鸟往往分布在植被量低且水域开阔的生境中; 到居民点距离或到道路距离对食块茎水鸟、食苔草水鸟、食种子水鸟和食鱼水鸟均产生显著性影响(<0.05)。因此, 在菜子湖湿地保护和管理时应考虑环境因子对不同取食集团越冬水鸟影响的差异。
冬季水鸟; 菜子湖; 环境因子; 取食集团; 空间分布
0 前言
越冬期是水鸟周年生活史的重要组成部分, 冬季水鸟群落组成和空间分布可以充分地反映栖息地质量状况[1–2]。冬季水鸟栖息地环境因子的改变会直接影响水鸟在栖息地的空间分布以及身体状况, 进而影响春季候鸟的离开时间和随后的成功繁殖[3]。因此, 了解冬季水鸟的空间分布及其与环境因子之间的关系非常重要。以往的生态学研究表明, 环境因子对鸟类的影响是由多因素决定的[4–6]。首先, 环境因子对不同取食集团和物种之间都存在着较大的差异, 不同取食集团的水鸟通过不同的方式有效地利用生物资源[7–8]。因此, 在分析湿地水鸟分布时, 应考虑水鸟物种的生态学特征。其次, 在较小的空间尺度上, 其相关研究通常侧重于栖息地内环境因子的改变, 如景观特征[9]、栖息地变化[10]、水位波动[11]、人为干扰[12]等。在这些因素中, 生境特征和人为干扰被认为是影响越冬水鸟空间分布和数量的最重要因素[13–14]。
菜子湖是长江中下游典型的浅水通江湖泊, 呈现秋季下降、春夏季上涨的季节性水位波动特征, 使得丰富的营养物质和大量的沉积物在湖泊中积累, 进而使得湖泊具有较高的生产力, 在冬季枯水期为全球性的迁徙水鸟提供了重要的食物资源[15–16]。每年成千上万的迁徙水鸟被吸引到此停歇或越冬, 水鸟数量在每年12—1月达到峰值并保持相对稳定[17–18]。然而未来几年, 由于引江济淮工程的全面施工, 必将导致菜子湖冬季水鸟数量的减少以及环境因子的改变, 因此, 有必要进一步深入研究水鸟的空间分布与环境因子之间的关系。
本文在2019—2020年的菜子湖湿地冬季水鸟调查基础上, 以菜子湖不同取食集团水鸟为研究对象, 应用空间统计学和多元逐步回归, 研究冬季水鸟的空间分布特征及其环境因子的关系, 并筛选出关键环境因子。从而为菜子湖湿地水鸟保护和栖息地的管理提供建议。
1 材料与方法
1.1 研究区域
菜子湖(30°42′—30°59′ N, 116°58′—117°12′ E)位于东亚-澳大利西亚候鸟迁徙线路上, 全湖由白兔湖、菜子湖、嬉子湖3个子湖连接而成。菜子湖区位于整个湖泊的中部, 在松山和车富岛处与白兔湖南北向相通, 在小麦嘴与嬉子湖东西向相连。菜子湖湿地位于亚热带湿润气候区, 受季风气候的影响, 在每年的6—8月降雨量较大, 平均降雨量为1344 mm,气温在7月达到最高, 1月达到最低, 年平均气温为16.5 ℃, 年无霜期约250 d。独特的地理位置和多样的气候条件, 使得菜子湖具有丰富的生物资源, 逐渐成为候鸟栖息和停歇的必经之路。
1.2 鸟类调查、分类和多样性分析
越冬水鸟大约在每年的10月飞至菜子湖越冬, 到翌年3月份离开, 在越冬前、后期, 由于菜子湖作为水鸟的中途停歇地, 其数量和空间分布并不稳定。而水鸟数量在12—1月份达到峰值并保持相对稳定, 在这个时期能够更加充分的反映菜子湖越冬水鸟的空间分布及其与环境因子的关系[3]。因此, 本研究分别于2019年12月14日和2020年1月7日对菜子湖冬季水鸟进行调查, 利用0.01°×0.01°的网格将鸟记录在网格中, 总共选取了51个采样点, 并确保在采样点附近能够观测到菜子湖内的所有网格。
为避免在调查过程中对水鸟重复计数, 我们将调查人员分成4组进行同步调查, 每组负责其相对应的网格范围。调查人员使用双筒望远镜(EL 10×32 WB, WAROVSKI)、单筒望远镜(ATS 20—60×80 HD, SWAROVSKI)和手持GPS接收机采用直接计数法对水鸟进行计数。同时记录鸟类的种类、数目、生境以及鸟类在网格中的具体位置。
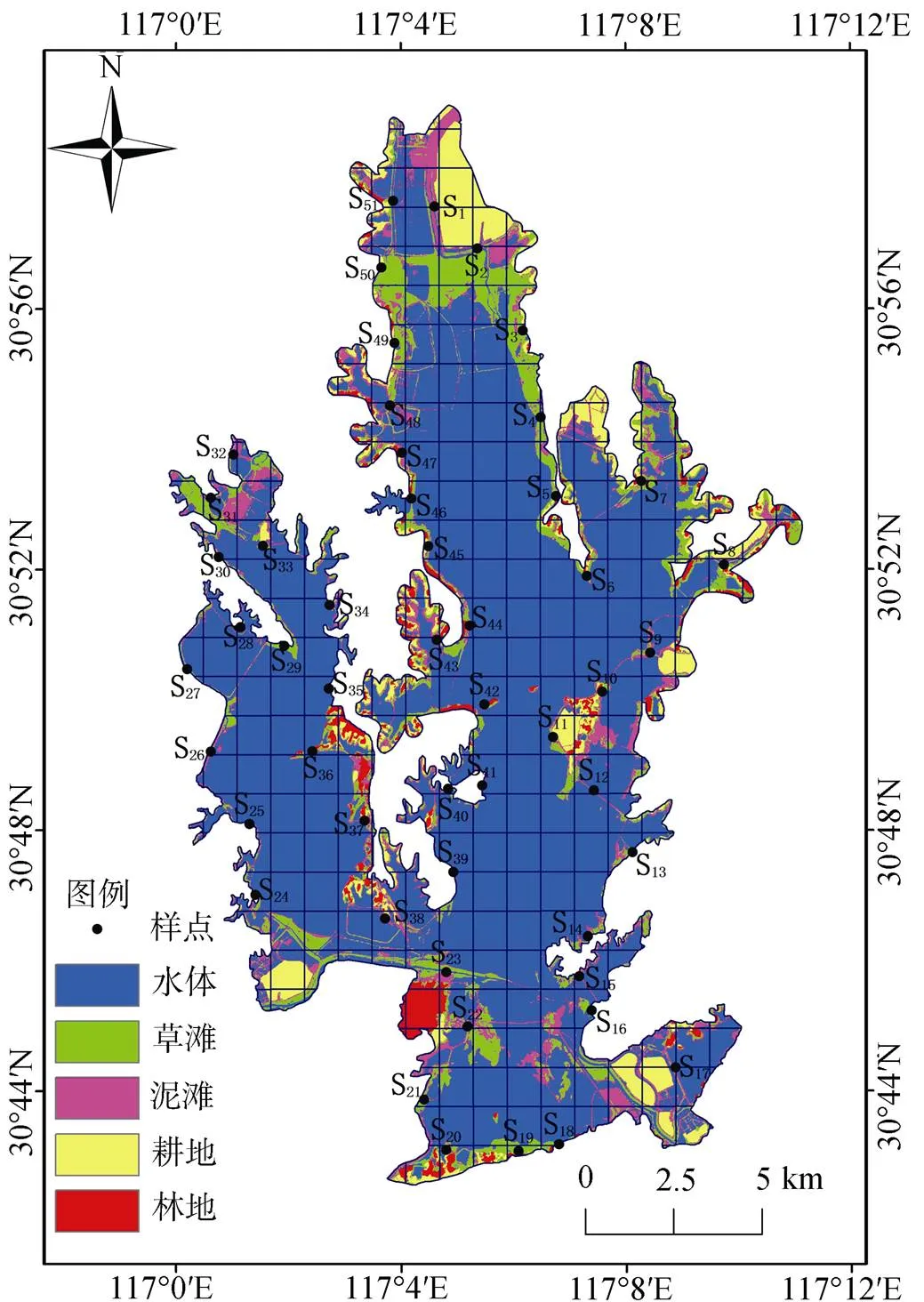
注: S1: 梅花大圩西; S2: 梅花大圩南; S3: 双兴村; S4: 李家湾; S5: 八家; S6: 大胡庄; S7: 马安; S8: 中屋; S9: 荣家埠; S10: 车富岛北; S11: 车富岛西; S12: 车富岛南; S13: 汤家咀; S14: 董庄; S15: 杨家冲; S16: 燕窝山; S17: 鲍洼; S18: 狮岭村; S19: 元家桥; S20: 龙家大屋; S21: 尤庄; S22: 堂屋南; S23: 堂屋北; S24: 古塘胜利; S25: 枫树窑; S26: 西瓜咀; S27: 许咀头; S28: 周家咀; S29: 大坟墓; S30: 赵店村; S31: 季庄; S32: 王家同; S33: 西庄; S34: 王家老屋; S35: 钱家咀; S36: 渔业村; S37: 大屋; S38: 榆咀; S39: 老屋; S40: 六家湾; S41: 苏家咀; S42: 方汪; S43: 胡老屋; S44: 张瓦屋; S45: 陈家湾; S46: 小杨庄; S47: 鸽子山; S48: 松杨咀; S49: 西家坂; S50: 方家咀; S51: 伍家老屋。
Figure 1 Location of study area
为了更好的分析环境因子对冬季水鸟空间分布的影响, 考虑水鸟物种的生态学特征, 根据水鸟在菜子湖的主要觅食习性, 将水鸟分成5组, 即食鱼水鸟、食无脊椎水鸟、食块茎水鸟、食苔草水鸟和食种子水鸟[19–20], 统计网格水鸟数量百分比, 即监测点网格中所有水鸟的个体数量占全湖网格中全部水鸟个体数量的百分比。
湿地水鸟的生物多样性使用Shannon-Wiener多样性指数()进行衡量[10]:
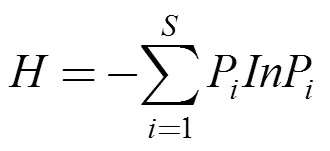
其中,代表网格内物种数,P是第种物种数量占所有物种数量的百分比。
P≥10%为优势种, 1%≤P<10%为常见种,P<1%为稀有种[21]。
1.3 环境因子的调查
通过实地考察和已有研究的基础上, 选取草滩面积、水域面积、泥滩面积、斑块密度、NDVI、到居民点距离和到道路距离作为环境因子。根据调查日期和遥感影像质量, 选择距离调查日期较近的哨兵2号(https:// scihub.copernicus.eu/dhus/#/home)和高分一号(http://www.cresda.com/CN)遥感影像。通过SNAP软件和ENVI 5.3软件对遥感影像进行预处理。道路和居民点的信息在Google Earth上提取。
根据前人研究和实地调查并结合菜子湖景观空间分布特征, 影像采用监督分类、神经网络进行解译, 分为草滩、水体、泥滩、耕地和林地五类, 并对分类精度进行检验, 精确度较高均达90%以上, 可以作为研究使用[22]。在ArcMap 10.2中对解译结果按照0.01°×0.01°的网格进行裁剪并转换为tiff格式, 导入Fragstats 4软件中计算鸟所在网格中的草滩面积、水域面积、泥滩面积和斑块密度[23]。最后运用ArcMap 10.2的空间分析, 得出鸟到道路及居民点的距离。运用ENVI软件的波段运算从遥感影像中提取鸟所在网格的归一化植被指数(), 计算公式如下:
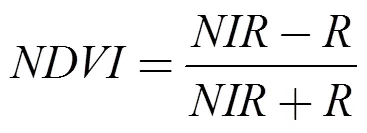
代表近红外波段的反射值;代表红光波段的反射值。
1.4 空间统计分析
通过空间自相关分析可以得到水鸟所在网格单元的数量是否与其相邻网格单元存在相关性, 进而反映出水鸟数量在空间上的聚集程度和空间差异性[24]。本文采用全局莫兰指数对菜子湖冬季水鸟的空间分布进行空间自相关分析, 其计算公式如下:
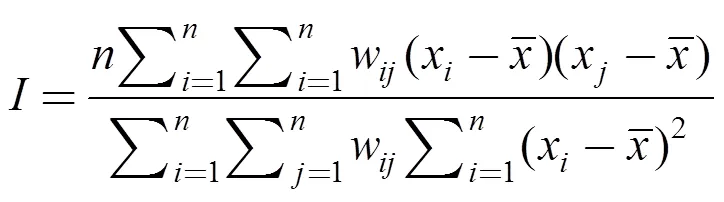
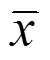
1.5 多元逐步回归分析
对网格中不同食性冬季水鸟多度与环境因子进行Spearman相关分析, 从而得到不同取食集团冬季水鸟多度变化与所选环境因子的密切程度。多元逐步回归是运用线性关系构建多个自变量和因变量之间的关系, 并考虑方差异质性, 剔除影响方程准确性的因子, 从而获得自变量对因变量的影响程度。因此, 当环境因子与水鸟呈现出显著性相关时(<0.05), 建立环境因子与物种数据的多元逐步回归模型, 从而筛选主要的影响因子[26–27]。
为了消除环境因子由于量纲不同而造成的影响, 在进行逐步回归之前, 需要对环境因子进行标准化处理。同时, 建立环境因子与物种数据的逐步回归模型, 需要不同取食集团冬季水鸟数据服从正态分布, 本文通过“个案排秩”转换得到不同取食集团冬季水鸟数量的正态得分值[28], 并通过正态性检验(>0.05), 说明经过“个案排秩”转换后的数据服从标准正态分布。然后, 对转换后的环境因子与物种数据进行逐步回归运算。以上所有统计分析均在SPSS 22中进行。
2 结果
2.1 水鸟种类和数量
2019—2020年冬季两次水鸟调查总共记录水鸟49802只水鸟, 隶属6目12科41种。其中食苔草水鸟数量最多, 食块茎水鸟最少。种群数量最多的7种水鸟分别是豆雁()、黑腹滨鹬()、绿翅鸭()、斑嘴鸭()、白额雁()、普通鸬鹚()、白琵鹭(), 其数量占总数的84.9%。在41种鸟类中, 被纳入IUCN红色名录鸟类7种, 包括极危物种白鹤(); 濒危种东方白鹳(); 易危种白头鹤()和鸿雁(); 近危种凤头麦鸡()、罗纹鸭()和白腰杓鹬()(表1)。其中普通鸬鹚、白琵鹭、白额雁、豆雁、白头鹤和东方白鹳的数量超过东亚-澳大利西亚候鸟迁徙路线种群大小的1%。
2.2 水鸟的空间分布
菜子湖冬季水鸟调查表明, 菜子湖冬季水鸟主要分布在堂屋、梅花大圩、季庄、车富岛以及双兴村, 其数量分别占总数的30.98%、10.53%、7.87%、6.49%和5.67%。菜子湖各子湖区冬季水鸟的组成差异较大, 水鸟主要分布在菜子湖和白兔湖, 其数量占总数的82.87%, 而在嬉子湖分布的数量较少, 仅仅占总数的17.13%。而对于濒危物种而言, 其空间分布与水鸟种群数量分布相似, 濒危物种在白兔湖和菜子湖中分布的比例较大, 占总数的78.46%, 而嬉子湖仅仅占有21.54%, 其中梅花大圩为易危物种白头鹤()在菜子湖的主要越冬地。在此次调查中, 白兔湖的水鸟种类最多为35种, 其次是菜子湖为29种, 嬉子湖水鸟种类最少为24种。白兔湖、菜子湖、嬉子湖中冬季水鸟的Shannon-Wiener多样性指数分别为2.12、1.61、1.86。但在利用网格单元的Shannon-Wiener多样性指数得到的空间分布图中, 菜子湖的多样性指数要高于嬉子湖(图2)。
对菜子湖冬季水鸟进行空间自相关分析, 发现不同取食集团冬季水鸟在菜子湖的分布特征存在差异。其中, 食苔草水鸟和食块茎水鸟在菜子湖的空间分布呈现显著的正相关(<0.05,>0), 表现为聚集分布。而食鱼水鸟、食无脊椎水鸟和食种子水鸟不具有显著性相关(>0.05), 表现为随机分布。对于整个菜子湖冬季水鸟而言, 由于食苔草水鸟占水鸟总数的比例较大, 进而使得冬季水鸟在菜子湖的空间分布呈现出正相关(<0.05,>0), 表现为聚集分布(表2)。
2.3 水鸟与环境因子的关系
对不同食性冬季水鸟与环境因子进行Spearman相关分析, 结果表明(表3),食鱼水鸟与水域面积、到居民点距离和到道路距离达到极显著正相关(< 0.01); 与草滩面积、斑块密度和NDVI之间的相关性达到极显著负相关(<0.01)。食无脊椎水鸟与泥滩面积和NDVI达到极显著正相关(<0.01); 与水体面积达到极显著负相关(<0.01)。食块茎水鸟与NDVI、草滩面积和到居民点距离达到显著正相关(<0.05)。食苔草水鸟与草滩面积、NDVI、到居民点距离和到道路距离达到极显著正相关(<0.01)。食种子水鸟与水域面积达到极显著正相关(P<0.01); 与到居民点距离达到显著正相关(P<0.05); 与草滩面积、泥滩面积、NDVI达到极显著负相关(<0.01); 与斑块密度达到显著负相关(<0.05)。

表1 菜子湖冬季水鸟种类和数量
注: 国I、国II分别为国家I级、II级重点保护物种, 省I、省II分别为安徽省地方I级、II级重点保护物种; “+++”: 优势种, “++”: 普通种, “+”: 稀有种。

图2 菜子湖冬季水鸟空间分布图(a. 个体数量; b. 物种数; c. 多样性指数)
Figure 2 Spatial distribution of wintering waterbirds at Caizi Lake (a. abundance, b. number of species, c. diversity index)

表2 菜子湖冬季水鸟空间自相关检验
为了分析菜子湖冬季水鸟空间分布及其与环境因子的关系, 采用多元逐步回归来剔除未通过显著性检验的环境因子, 从而建立起环境因子和不同取食集团水鸟间的最优回归方程。由表4可知, 逐步回归方程的决定系数均为0.5<R<1, 表明主要环境影响因子已基本考虑进去, 剩余因子包括其他影响较小的因子、未研究的环境因子和试验误差[29]。逐步回归方程的回归关系达到极显著水平(<0.01), 方程的相关系数均大于0.7, 可知该方程的拟合度较好, 说明模型可以很好反映菜子湖冬季水鸟与环境因子之间的关系[30]。统计量均接近2, 表明残差之间相互独立, 水鸟与环境因子间构建的回归方程具有一定的可信度。

表3 菜子湖不同食性冬季水鸟与环境因子的相关性分析
注: * 表示该自变量与因变量之间的相关性<0.05; ** 表示该自变量与因变量之间的相关性<0.01。
由表5可知, 通过回归分析得出各环境因子的值均小于2, 说明因子之间不存在严重的共线性问题。并且, 通过逐步回归所选出的环境因子其检验的显著性水平均小于0.05。结果表明, 环境因子对不同取食集团水鸟的影响不同。对于食鱼水鸟而言, 其关键环境因子是水域面积和到居民点距离。食无脊椎水鸟的关键环境因子是泥滩面积和水域面积, 食块茎水鸟的关键环境因子是NDVI和到居民点距离。食苔草水鸟的关键环境因子是草滩面积和到道路距离。食种子水鸟的关键环境因子是草滩面积、水域面积和到居民点距离。
3 讨论
3.1 水鸟的空间分布格局
以往研究表明, 水鸟在湿地中的空间分布与栖息地特征息息相关, 如水位、水面面积、植被面积、景观结构等[31–32]。在湖泊水位下降的过程中, 由于菜子湖和白兔湖区内的地势较高, 导致湖区底部暴露出来而形成大面积的草滩和泥滩, 为菜子湖越冬水鸟提供更多的食物资源和栖息环境。而嬉子湖由于地势较低, 草滩分布面积及范围较少, 深水区域分布较广。从而导致菜子湖和白兔湖内冬季水鸟的种类、数量都要远高于嬉子湖。菜子湖区由于在一些网格中有大面积连续草洲而聚集上千只豆雁, 使得豆雁数量占绝对优势(60%以上), 而其他水鸟占水鸟总数的比例较低, 导致整个菜子湖区的Shannon-Wiener多样性指数低于嬉子湖。但是在其他网格单元中, 菜子湖区的局部小生境与嬉子湖相比更加复杂, 空间异质性较高, 进而形成Shannon- Wiener多样性指数的空间分布图中, 菜子湖区要高于嬉子湖区[33]。

表4 菜子湖不同食性冬季水鸟与环境因子的逐步回归分析(模型汇总)
在菜子湖中, 大多数水鸟都具有流动性, 它们一般随着自身生存需要的食物资源而聚集或分散[34]。菜子湖是一个通江湖泊, 其鱼类可以在水域连通的条件下, 在不同生境斑块间迁移和洄游完成其生活史周期和节律, 使得鱼类不会固定在某一个区域, 而是处于一种流动的状态[35–36], 进而影响食鱼水鸟在菜子湖中的分布不具有空间自相关, 呈现为随机分布。食无脊椎水鸟随机分布的原因可能是冬季菜子湖中的大坝和养殖塘的随机排水而出现了大面积的泥滩和浅水水面。菜子湖沉水植物的稀缺, 使得食块茎水鸟改变觅食生境而聚集分布在草滩和稻田[37]。食苔草水鸟一般会集群出现在苔草洲滩进行觅食, 因此在空间分布上表现为聚集分布。大多数的食种子水鸟一般白天在安全的栖息地(湖泊、池塘等)进行休息, 日落后则飞到附近的农田、滩涂和草地进行觅食[38]。它们经常在觅食地和栖息地之间飞行, 并且这种飞行不是定向的而是分散的, 使得它们特别是在冬季表现出高度的区域流动性[39], 从而使得食种子水鸟的空间分布类型是随机的。

表5 菜子湖不同食性冬季水鸟与环境因子的逐步回归分析(系数)
3.2 环境因子对冬季水鸟的影响
在本研究中, 观察到的食鱼水鸟中深水食鱼水鸟(银鸥、红嘴鸥及潜水鸟)所占比例较大, 而浅水食鱼水鸟(鹭科和东方白鹳)相对较小。浅水食鱼水鸟多在浅滩被观察到, 而深水食鱼水鸟却分布在开阔的水域[15]。同时, 更大的水域面积为食鱼水鸟提供了更丰富的食物资源和食物种类, 并减少了干扰和竞争[40]。食无脊椎水鸟主要以滩涂湿地的摇蚊属、腹足类等无脊椎底栖生物为食, 一般会选择水深不超过5 cm的区域, 因为该区域的底栖生物较为丰富, 能够为食无脊椎水鸟提供充足的食物[41]。因此, 浅水和泥滩对食无脊椎集团水鸟至关重要。在上个世纪, 菜子湖的食块茎水鸟主要在浅水和湿润的泥滩中挖掘大型沉水植物的块茎为食[5,42]。然而在过去20年内, 菜子湖由于密集的水产养殖导致湖泊内沉水植物的快速消失, 进而引起湖泊湿地退化严重和食块茎水鸟的急剧下降[35,43]。滩涂和浅水中块茎的稀缺, 使得食块茎水鸟觅食地由泥滩生境向草滩和稻田生境转移, 从而导致菜子湖食块茎水鸟主要分布在梅花大圩、双兴邻近的草滩和稻田。这与Li[44]等和Zheng[45]等的研究结果一致, 这样就解释了食块茎水鸟与NDVI呈正相关而与泥滩面积无关。雁类通常会选择低纤维高蛋白的新生植物为食, 冬季湖泊水位下降而形成的洲滩中苔草迅速萌发生长, 使得食苔草水鸟被吸引在新出露的草地上进行觅食, 其环境容纳量与草滩中的苔草生物量呈指数式增长[46]。食种子水鸟主要以涉水鸭为主, 它们一般会在较高水位和开阔水面的生境进行栖息, 避开植被生物量较大的区域[14,47]。因此, 在草滩面积较大的地区, 它们的数量较少。
人类的干扰往往会促使水鸟投入更多的时间保持警戒, 从而使水鸟的觅食时间减少, 进而导致水鸟能量摄入的减少。甚至在人为干扰较为严重的时候, 人类活动迫使水鸟放弃生境较好的觅食地、提高其在飞行的能量消耗和在不太理想的栖息地中觅食[48–49]。在人为干扰方面, 本文选取了到居民点距离和到道路距离两个环境因子。结果表明, 到居民点距离或到道路距离对食鱼水鸟、食块茎水鸟、食苔草水鸟和食种子水鸟均产生显著性影响。在此次冬季水鸟调查中, 菜子湖的水位与往年相比较高, 导致暴露的泥滩较少, 与此同时, 在一些离居民点或者道路不远的养殖塘中发现大量的鸻鹬类水鸟, 这是由于渔业生产过程中水位降低进而暴露出大量的泥滩, 吸引鸻鹬类水鸟在此觅食, 导致在本次研究中这两个环境因子没有对食无脊椎水鸟产生显著性影响。
4 结论
菜子湖冬季水鸟主要分布在堂屋、梅花大圩、季庄、车富岛以及双兴村, 并且在各子湖区水鸟的组成差异较大, 菜子湖和白兔湖内冬季水鸟的种类、数量都要远高于嬉子湖。食苔草水鸟和食块茎水鸟的空间格局为聚集分布, 而食鱼水鸟、食无脊椎水鸟和食种子水鸟为随机分布。环境因子对不同取食集团水鸟的影响也有所不同, 生境因子和人为干扰是影响水鸟空间分布的主要因子。
[1] GUNARATNE A M, JAYAKODY S, BAMBARADENIYA C N B. Spatial distribution of aquatic birds in Anavilundawa Ramsar wetland sanctuary in Sri Lanka[J]. Biological Invasions, 2009, 11(4): 951–958.
[2] 蒋科毅, 吴明, 邵学新. 杭州湾及钱塘江河口冬季水鸟群落多样性及其空间分布[J]. 长江流域资源与环境, 2013, 22(5): 602–609.
[3] ZHANG Pingyang, ZOU Yeai, XIE Yonghong, et al. Shifts in distribution of herbivorous geese relative to hydrological variation in East Dongting Lake wetland, China[J]. Science of the Total Environment, 2018, 636: 30–38.
[4] DRONOVA I, BEISSINGER S R, BURNHAM J W, et al. Landscape–level associations of wintering waterbird diversity and abundance from remotely sensed wetland characteristics of Poyang Lake[J]. Remote Sensing, 2016, 8(6): 462.
[5] WANG Wenjuan, FRASER J D, CHEN Jiakuan. Wintering waterbirds in the middle and lower Yangtze River floodplain: changes in abundance and distribution[J]. Bird Conservation International, 2016, 27(2): 167–186.
[6] 黄越. 北京城市绿地鸟类生境规划与营造方法研究[D]. 北京: 清华大学, 2015.
[7] ZHANG Yong, JIA Qiang, PRINS H H T, et al. Effect of conservation efforts and ecological variables on waterbird population sizes in wetlands of the Yangtze River[J]. Scientific Reports, 2015, 5(1): 17136.
[8] 王亚芳. 鄱阳湖越冬水鸟群落对冬汛的响应及其对生态系统的指示[D]. 南昌: 南昌大学, 2018.
[9] 李相逸, 曹磊, 马超, 等. 天津滨海滩涂湿地鸟类丰富度与环境因子的关系研究[J]. 风景园林, 2018, 25(6): 107–112.
[10] 颜凤, 刘本法, 余仁栋, 等. 围填海对盐城珍禽自然保护区越冬水鸟群落及空间分布的影响[J]. 生态科学, 2018, 37(6): 20–29.
[11] RAJASHEKARA S, VENKATESHA M G. Eco–spatial and temporal variation in waterbirds composition and their relationship with habitat characteristics of Urban Lakes of Bengaluru city, India[J]. International Journal of Advanced Research, 2014, 2(7): 60–80.
[12] ROSIN Z M, SKÓRKA P, WYLEGAŁA P, et al. Landscape structure, human disturbance and crop management affect foraging ground selection by migrating geese[J]. Journal of Ornithology, 2012, 153(3): 747–759.
[13] YUAN Yujie, ZENG Guangming, LIANG Jie, et al. Effects of landscape structure, habitat and human disturbance on birds: A case study in East Dongting Lake wetland[J]. Ecological Engineering, 2014, 67: 67–75.
[14] ZHANG Yong, FOX A D, CAO Lei, et al. Effects of ecological and anthropogenic factors on waterbird abundance at a Ramsar Site in the Yangtze River Floodplain[J]. Ambio, 2019, 48(3): 293–303.
[15] LI Chunlin, LI Haifeng, ZHANG Yong, et al. Predicting hydrological impacts of the Yangtze–to–Huaihe Water Diversion Project on habitat availability for wintering waterbirds at Caizi Lake[J]. Journal of Environmental Management, 2019, 249: 109251.
[16] 周盛, 杨森, 鲍明霞, 等. 基于遥感反演预测“引江济淮”工程对菜子湖越冬水鸟栖息地的影响[J]. 生态科学, 2019, 38(1): 71–78.
[17] 陈锦云. 安徽沿江湖泊越冬水鸟群落结构研究[D]. 合肥: 安徽大学, 2011.
[18] 王晓媛, 江波, 田志福, 等. 冬季安徽菜子湖水位变化对主要湿地类型及冬候鸟生境的影响[J]. 湖泊科学, 2018, 30(6): 1636–1645.
[19] XIA Shaoxia, LIU Yu, WANG Yuyu, et al. Wintering waterbirds in a large river floodplain: Hydrological connectivity is the key for reconciling development and conservation[J]. Science of The Total Environment, 2016, 573: 645–660.
[20] WANG Yuyu, JIA Yifei, GUAN Lei, et al. Optimising hydrological conditions to sustain wintering waterbird populations in Poyang Lake National Natural Reserve: implications for dam operations[J]. Freshwater Biology, 2013, 58(11): 2366–2379.
[21] 宋昀微, 周立志. 生境变化对升金湖越冬水鸟群落时空动态的影响[J]. 安徽农业大学学报, 2019, 46(4): 610–617.
[22] ZHOU Yunkai, NING Lixin, BAI Xiuling. Spatial and temporal changes of human disturbances and their effects on landscape patterns in the Jiangsu coastal zone, China[J]. Ecological Indicators, 2018, 93: 111–122.
[23] DALLIMER M, MARINI L, SKINNER A M J, et al. Agricultural land–use in the surrounding landscape affects moorland bird diversity[J]. Agriculture, Ecosystems and Environment, 2010, 139(4): 578–583.
[24] 王芳, 袁兴中, 熊森, 等. 重庆澎溪河湿地自然保护区生物多样性空间格局及热点区[J]. 应用生态学报, 2020, 31(5): 1682–1690.
[25] DU Haiwen, WANG Yong, ZHUANG Dafang, et al. Temporal and spatial distribution characteristics in the natural plague foci of Chinese Mongolian gerbils based on spatial autocorrelation[J]. Infectious Diseases of Poverty, 2017, 6(1): 124.
[26] ZHU Zhengrong, ZHOU Lizhi, YU Chao, et al. Do geese facilitate or compete with wintering Hooded Cranes () for forage resources?[J]. Diversity, 2020, 12(3): 105.
[27] 罗宜富, 李磊, 李秋华, 等. 阿哈水库叶绿素a时空分布特征及其与藻类、环境因子的关系[J]. 环境科学, 2017, 38(10): 4151–4159.
[28] 蔡元刚, 王明田, 蔡怡亨, 等. 利用地面气象资料建立四川省日总辐射计算模型[J]. 中国农业气象, 2019, 40(9): 543–556.
[29] 唐朝臣, 罗峰, 李欣禹, 等. 甜高粱产量及品质相关性状对环境因子反应度分析[J]. 作物学报, 2015, 41(10): 1612–1618.
[30] 江敏, 余根鼎, 戴习林, 等. 凡纳滨对虾养殖塘叶绿素a与水质因子的多元回归分析[J]. 水产学报, 2010, 34(11): 1712–1718.
[31] REID J R W, COLLOFF M J, ARTHUR A D, et al. Influence of catchment condition and water resource development on waterbird assemblages in the Murray–Darling Basin, Australia[J]. Biological Conservation, 2013, 165: 25–34.
[32] MCKINNEY R A, RAPOSA K B, COURNOYER R M. Wetlands as habitat in urbanizing landscapes: Patterns of bird abundance and occupancy[J]. Landscape and Urban Planning, 2011, 100(1/2): 144–152.
[33] 颜凤, 李宁, 杨文, 等. 围填海对湿地水鸟种群、行为和栖息地的影响[J]. 生态学杂志, 2017, 36(7): 2045–2051.
[34] ZHANG Chang, YUAN Yujie, ZENG Guangming, et al. Influence of hydrological regime and climatic factor on waterbird abundance in Dongting Lake Wetland, China: Implications for biological conservation[J]. Ecological Engineering, 2016, 90: 473–481.
[35] WINEMILLER K O, MCINTYRE P B, CASTELLO L, et al. Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong[J]. Science, 2016, 351(6269): 128–129.
[36] 古辰, 姜美彤, 蒋忠冠. 生境过滤作用对长江下游菜子湖鱼类物种和功能组成的影响[J]. 湖泊科学, 2020, 32(1): 124–133.
[37] 宫蕾. 安徽沿江湖泊越冬白头鹤()觅食生态的研究[D]. 合肥: 安徽大学, 2013.
[38] TAJIRI H, OHKAWARA K. The effects of flooding and plowing on foraging site selection by sintering dabbling ducks in rice fields[J]. Ornithological Science, 2013, 12(2): 127–136.
[39] SOONS M B, VAN DER VLUGT C, VAN LITH B, et al. Small seed size increases the potential for dispersal of wetland plants by ducks[J]. Journal of Ecology, 2008, 96(4): 619–627.
[40] VAN DIJK J G B, DUIJNS S, GYIMESI A, et al. Mallards feed longer to maintain intake rate under competition on a natural food distribution[J]. Ethology, 2012, 118(2): 169–177.
[41] 胡越凯, 庞毓雯, 焦盛武, 等. 基于遥感的杭州湾湿地鸻鹬类水鸟适宜生境时空变化特征研究[J]. 杭州师范大学学报(自然科学版), 2019, 18(3): 319–328.
[42] ZHANG Yong, CAO Lei, BARTER M, et al. Changing distribution and abundance of Swan Goosein the Yangtze River floodplain: the likely loss of a very important wintering site[J]. Bird Conservation International, 2010, 21(1): 36–48.
[43] FOX A D, CAO Lei, ZHANG Yong, et al. Declines in the tuber–feeding waterbird guild at Shengjin Lake National Nature Reserve, China – a barometer of submerged macrophyte collapse[J]. Aquatic Conservation Marine and Freshwater Ecosystems, 2011, 21(1): 82–91.
[44] LI Chunlin, ZHOU Lizhi, XU Li, et al. Vigilance and activity time–budget adjustments of wintering Hooded Cranes,, in human–dominated foraging habitats[J]. PLOS ONE, 2015, 10(3): e0118928.
[45] ZHENG Meng, ZHOU Lizhi, ZHAO Niannian, et al. Effects of variation in food resources on foraging habitat use by wintering Hooded Cranes()[J]. Avian Research, 2015, 6(3): 186–195.
[46] 孟竹剑. 鄱阳湖越冬雁类栖息地模型及环境容纳量研究[D]. 南昌: 南昌大学, 2018.
[47] 牛俊英, 衡楠楠, 张斌, 等.上海市南汇东滩围垦后海岸带湿地冬春季水鸟生境选择[J].动物学研究, 2011, 32(6): 624–630.
[48] MARTÍN B, DELGADO S, DE LA CRUZ A, et al. Effects of human presence on the long–term trends of migrant and resident shorebirds: evidence of local population declines[J]. Animal Conservation, 2014, 18(1): 73–81.
[49] CHUDZIŃSKA M E, VAN BEEST F M, MADSEN J, et al. Using habitat selection theories to predict the spatiotemporal distribution of migratory birds during stopover – a case study of pink–footed geese[J]. Oikos, 2015, 124(7): 851–860.
Relationship between environmental factors and spatial distribution of wintering waterbirds at Caizi Lake, China
YAO Jian1,2, ZHOU Lizhi1,2,*, WEI Zhenhua1,2, LI Lingchen1,2
1. School of Resources and Environmental Engineering, Anhui University, Hefei 230601, China 2. Anhui Province Key Laboratory of Wetland Ecosystem Protection and Restoration(Anhui University), Hefei 230601, China
It is an important content of ecological research to study the relationship between the spatial distribution of waterbirds and environmental factors. Variations in environmental factors can directly or indirectly affect the species, abundance and distribution of waterbirds by changing habitat characteristics and food resources. In this study, we investigated the wintering waterbirds and environmental factors at Caizi Lake in China using grid cells, in the winter of 2019-2020. A total of 41 waterbirds species belonging to 6 orders of 12 families were counted during the surveys, and the dominant species were bean geese () and dunlin (). Spatial autocorrelation and multiple stepwise regression analyses were used to study the spatial distribution patterns of wintering waterbirds and determine the main environmental factors affecting their distribution in different foraging groups. The percentage of individuals of grid birds results showed that the wintering waterbirds at Caizi Lake were mainly distributed in Tangwu (30.98%), Meihuadawei (10.53%), Jizhuang (7.87%), Chefu Island (6.49%) and Shuangxing Village(5.67%). The distribution patterns of tuber-eating waterbirds and sedge-eating waterbirds revealed significantly positive autocorrelations and clustered distributions (<0.05,>0), while those of seed-eating waterbirds, invertebrate-eating waterbirds and fish-eating waterbirds showed non-autocorrelational and random distributions (>0.05). Environmental factors had differing effects on different waterbird foraging groups. Fish-eating waterbirds tended to choose the open water which has large areas, while the mudflats exposed by the wetland attracted invertebrate-eating waterbirds. The degradation of submerged vegetation led to the aggregation and distribution of tuber-eating waterbirds in farmland and grassland habitats.The quantity of sedge-eating waterbirds increased significantly because of the formation of vast grasslands. Seed-eating waterbirds were often distributed in low-vegetation and open water habitats. The distance to the nearest settlement or to the road had significant effects on tuber eaters, sedge eaters, seed eaters, and fish eaters (<0.05). Therefore, the influence of environmental factors on different foraging groups of wintering waterbirds should be considered in the protection and management of the Caizi Lake wetland.
wintering waterbirds; Caizi Lake; environmental factors; foraging groups; spatial distribution
10.14108/j.cnki.1008-8873.2022.01.001
姚简, 周立志, 魏振华, 等. 菜子湖冬季水鸟空间分布与环境因子的关系研究[J]. 生态科学, 2022, 41(1): 1–10.
YAO Jian, ZHOU Lizhi, WEI Zhenhua, et al. Relationship between environmental factors and spatial distribution of wintering waterbirds at Caizi Lake, China[J]. Ecological Science, 2022, 41(1): 1–10.
Q89
A
1008-8873(2022)01-001-10
2020-10-27;
2020-11-15
国家自然科学基金项目(32171530); 引江济淮工程菜子湖候鸟越冬期湿地生境保护适应性调度试验研究及生态环境监测项目(YJJH-ZT-ZX-20180404062)
姚简(1994—), 男, 江西萍乡人, 硕士, 主要从事环境生态与资源管理研究, E-mail: yaojian7219@163.com
周立志, 男, 博士, 教授, 主要从事水鸟与湿地生态学和环境生态与资源管理研究, E-mail: zhoulz@ahu.edu.cn
