萱草属植物F1 代花部性状遗传分析及QTL 定位
2022-03-03张玲玲李森刘娟公菲菲高阳侯非凡亢秀萍王艳芳邢国明
张玲玲,李森,刘娟,公菲菲,高阳,侯非凡,亢秀萍,王艳芳,邢国明*
(1.山西农业大学 园艺学院,山西 太谷 030801;2.山西省设施蔬菜提质增效协同创新中心,山西 太谷 030801;3.大同黄花产业发展研究院,山西 大同 037004)
萱草属植物种质资源丰富,在我国有着2000多年的栽培历史,是国际公认的三大宿根花卉之一。APG IV 分类系统将其划分为单子叶植物纲(Monocotyledoneae)阿福花科(Asphodelaceae)萱草亚科(Hemerocallidoideae)[1]。因其花型花色丰富多彩,花期绿期长,已成为重要的园林绿化材料[2]。萱草属植物包括夜间开放的黄花菜类群和白天开放的萱草类群[3],在长期栽培过程中,自然杂交和人工杂交普遍存在,经过100 多年的育种研究,品种已达8 万以上[4]。不同品种在花被片长宽、花朵开放和闭合时间、花色和花型等表型性状方面存在着极大的差异[5]。目前杂交育种是萱草的主要育种方式之一[6],种间杂交是创造新种质和选育新品种的重要育种技术和方法,可创造丰富的种质资源,杂交后代的杂种优势一般比较强[7]。
当今,随着测序技术的发展和新型标记的开发,国内外研究学者已在菊花[8-9]、百合[10]、月季[11]、康乃馨[12]等多种观赏植物中陆续展开数量性状基因定位研究。花器官是萱草属植物观赏性状中最直观的外在表现,也是其育种的主要目标之一。花朵直径、内外花瓣长度和宽度是萱草属植物的主要观赏性状。
本研究以黄花菜地方品种‘大同黄花’为母本(P1),萱草品种‘摇篮曲’为父本(P2)构建杂交F1群体(71 株)为材料,对其5 个花部性状:花朵直径、外瓣长度、外瓣宽度、内瓣长度和内瓣宽度进行统计分析,探讨表型性状遗传规律及变异特点,并利用SNP 标记对5 个花部性状进行QTL 定位研究,旨在为萱草属植物花部性状的分子标记辅助育种提供理论依据。
1 材料与方法
1.1 试验材料
以黄花菜地方品种‘大同黄花’为母本(P1),萱草品种‘摇篮曲’为父本(P2)构建杂交后代,采用常规田间管理。试验材料种植、杂交、取样均在萱草属植物种质资源圃(山西农业大学)中进行。

图1 杂交亲本(A)和杂交F1 代花器官(B)Fig.1 Floral organs of Hybrid parents(A)and hybrid F1 generation(B)
1.2 性状测定方法
于2019 年7-8 月观测亲本与杂交F1 群体的花朵直径(Flower diameter,FD)、外瓣长度(Sepal length,SL)、外瓣宽度(Sepal width,SW)、内瓣长度(Petal length,PL)和内瓣宽度(Petal width,PW)5 个花部性状,每个性状重复观测3 次,统计3 次重复的平均值。各性状测量标准如表1。

表1 杂交F1 花部性状测定标准Table1 Determination standard of flower characters of Hy⁃brid F1
1.3 主要花部性状杂种优势分析
‘大同黄花’和‘摇篮曲’杂交构建的F1 代杂种优势通过中亲优势和中亲优势率、超亲优势和超亲优势率的大小来表示。
杂交后代各性状的中亲优势(Hm)以杂交后代5 个花部性状各自的平均值(Fm)与中亲值(MPV)之差来表示。中亲优势率通过公式RHm(%)=[Hm/MPV]×100 计算。超亲优势(H)则通过杂交后代各性状的平均值(Fm)与高亲值(HP)之差表示。超亲优势率RH(%)=[H/HP]×100[13]。
使用Excel 2010 对5 个花部性状表型数据进行初步统计分析,用IBMSPSS Statistics 23 统计软件进行Pearson 相关分析。
1.4 QTL 作图
基于课题组前期构建的种间高密度遗传图谱[14-15],运用Map QTL 6.0 软件对杂交F1 代的主要花部性状数据进行QTL 定位分析。首先利用置换检验做1000 次重复,估算F1 群体中LOD 阈值大小。然后利用区间作图法(Interval Maping)对5 个花部性状进行QTL 分析,扫描步长为1 cM。区间作图法分析检测得到5 个花部性状相关QTL位点后,将这些位点中最高LOD 值所在位置的标记或与其紧密连锁的标记作为协同因子,再对检测到的QTL 进行MQM(多座位QTL 模型)检测,以LOD 值最高的位点作为QTL 所在的位置。
各性状QTL 命名的方式是:q,性状英文缩写名称(首字母大写),所在连锁群,QTL 在该连锁群上的顺序。例如qSL1.1 是指第1 连锁群上外瓣长度的第1 个QTL[16]。
2 结果与分析
2.1 萱草属植物杂交F1 群体主要花部性状遗传变异分析
2.1.1 杂交双亲及其杂交后代群体主要花部性状表型分析
本试验中父母本5 个花部性状的描述性数据如表2 所示。其中,母本‘大同黄花’花朵直径达11.50 cm,外瓣长宽分别为9.65 cm,1.25 cm,内瓣长宽为9.63 cm,1.93 cm;父本‘摇篮曲’花朵直径为10.17 cm,外瓣长宽和内瓣长宽为:8.20 cm,3.74 cm,7.12 cm 和4.87 cm。

表2 杂交双亲主要花部性状统计Table2 Statistics of main flower traits of the parents cm
对杂交F1 代花朵直径、外瓣长度、外瓣宽度、内瓣长度和内瓣宽度进行描述性统计分析,结果见表3。杂交F1 群体中5 个花部性状的变异系数为7.01%~12.50%,其中外瓣长度的变异系数最小,分离不明显。外瓣宽度和内瓣宽度的变异系数为12.39%和12.50%,表明这2 个性状在F1 个体间的变异较大。供试的71 份材料花朵直径最大值为11.46 cm,最小值为8.31 cm,外瓣长宽变异范围为6.90~9.63 cm,1.50~2.76 cm,内瓣长宽变异范围为6.85~9.84 cm,2.08~3.96 cm。
从表3 列出的偏度和峰度及杂交F1 群体各花部性状的频率分布图表明(图2),花部性状大部分都介于父母本之间,均表现出较好的连续性正态分布趋势,由此推测5 个花部性状均属于多基因控制的数量性状。
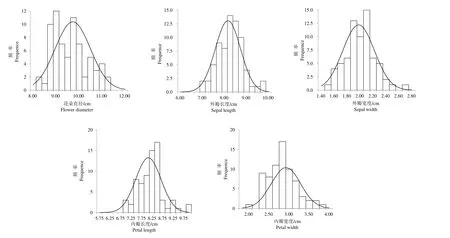
图2 杂交F1 代主要花部性状的频率分布Fig.2 Frequency distribution of main flower traits in Hybrid F1

表3 ‘大同黄花’与‘摇篮曲’杂交F1 群体主要花部性状特征值Table3 Main flower traits values in the Hybrid F1 of′Datong Huanghua′and ′Yaolanqu′
2.1.2 杂交F1 群体表型性状的遗传变异分析
对杂交F1 群体的5 个花部性状进行统计分析。结果如表4 所示,F1 中花朵直径的平均值相当于中亲值的89.67%,绝大多数F1 代花朵直径小于双亲中的低亲个体,表明花朵直径有变小的趋势,介于双亲之间的个体数占23.94%,没有超过高亲的个体。外瓣长度的平均值占中亲值的91.27%,有53.52% 个体的外瓣长度小于低亲,46.48%介于双亲之间,没有出现高于高亲的个体。表明F1 代的花朵直径和外瓣长度有一定的退化趋势。外瓣宽度和内瓣宽度平均值占中亲值的百分比分别为79.20%和85.88%,均没有低于低亲和高于高亲的个体。5 个花部性状中的内瓣长度平均值相当于中亲值的96.42%,其中杂交F1中有4.23%小于低亲个体,介于双亲之间的个体占92.96%,在内瓣长度这一性状中有2.81%的F1 代为超亲个体。

表4 杂交F1 各性状遗传变异分析Table4 Genetic variation analysis of traits in Hybrid F1
2.1.3 杂交F1 群体杂种优势分析
杂交F1 群体中5 个花部性状的杂种优势如表5 所示,均表现为负向中亲优势值,中亲优势率为−20.80%~−3.58%,其中以花朵直径的负向中亲优势最强,为−20.80%;以内瓣长度的负向中亲优势最弱,为−3.58%。说明5 个花部性状的杂种优势在F1 代表现为下降趋势性。T 检验表明,F1 群体中5 个表型性状中亲优势值均达到极显著水平(P<0.01)。其次根据图2 和表3 可发现,杂交F1 群体中除内外瓣宽度外,均存在正向或负向超出父母本的个体,表明超亲分离现象在杂交F1 代中普遍存在。而F1 群体各性状的平均值均介于父本和母本之间,且通过计算其超亲优势均为负值,说明在F1 群体中5 个花部性状的超亲分离普遍现象未演化成超亲优势。

表5 ‘大同黄花’与‘摇篮曲’杂交F1 群体主要花部性状的杂种优势表现Table5 Heterosis of main flower characters in F1 Progeny of′Datong Huanghua′and ′Yaolanqu′
2.1.4 杂交F1 群体主要花部性状的相关性分析
对杂交F1 群体的5 个花部性状进行皮尔逊相关分析(表6)后发现,5 个表型性状产生的10 对相关性中6 对相关性达到极显著相关(P<0.01),且6 对性状的相关性呈现极显著正相关。其中,花朵直径与外瓣长度、花朵直径与内瓣长度、外瓣长度与内瓣长度的相关系数均大于0.5,分别为0.623、0.642 和0.936,表明这3 对性状间相关性紧密。而内瓣宽度与其他性状之间不相关。

表6 杂交F1 群体5 个花部性状皮尔逊相关性分析Table6 Pearson correlation of 5 flower traits in F1 progeny
2.2 杂交F1 群体主要花部性状QTL 定位
基于萱草属种间杂交高密度遗传图谱对F1 群体5 个花部性状进行QTL 定位分析,估计每个QTL 对表型的贡献率(表7)。结果表明,共检测到16 个控制5 个花部性状的QTLs,分布在第2、3、4、5、8、9、10、11 号 连 锁 群 上,LOD 的 峰 值 介 于2.06~4.17 之间,单个QTL 的贡献率为12.5%~23.7%。控制花朵直径的QTL 位点共3 个,分别位于2、4、5 号连锁群上,贡献率分别为14.0%、16.2% 和13.6%。 其 中 位 于4 号 连 锁 群 上15 955 563~18 093 408 间的qFD4.1 贡献率最高,区间长度为1.613 cM。与外瓣长度相关的QTL位点共3 个,分别位于4、8、11 号连锁群上,贡献率分别为19.3%、12.9%和23.7%。其中位于11 号连锁群上59 674 131~60 681 150 间的qSL11.1 贡献率最高,区间长度为1.290 cM。在2、5、9 号连锁群上分布有3 个与外瓣宽度相关的QTL 位点,贡献率分别为17.3%、13.8%和16.1%。其中位于2 号 连 锁 群 上291 392 439~292 229 183 间 的qSW2.1 贡献率最高,区间长度为0.645 cM。我们检测到3 个与内瓣长度相关的QTL 位点,分别位于8、10、11 号连锁群上,贡献率分别为17.1%、23.3% 和21.1%。 其中位于10 号连锁群上46 864 791~47 992 915 间 的qPL10.1 贡 献 率 最高,区间长度为0.967 cM。与内瓣宽度相关的QTL 位点共3 个,分别位于3、4、10 号连锁群上,贡献率分别为12.9%、14.3%和12.5%。其中位于4 号 连 锁 群 上283 686 883~285 329 645 间 的qPW4.1 贡献率最高,区间长度为2.581 cM。

表7 杂交后代群体5 个花部性状QTL 分析Table7 QTL mapping of 5 flower traits in F1 progeny
3 讨论
杂交是将优良性状或基因转移到后代的重要技术手段,是选育有价值的新品种的有效方法之一[17]。杂交后代的遗传与变异规律可通过遗传标记来进一步明确。花部性状作为主要观赏性状是对观赏植物品质进行选择的重要参考依据。高超利等[18]将萱草品种‘金娃娃’与‘红宝’杂交发现,杂交后代变异系数最小的是单株花葶数,最大的是花序着花数,而花期变异不大。赵珺等[6]将野生萱草品种‘北黄花菜’与大花萱草品种‘回复’和‘玫瑰回旋’进行杂交,杂交后代中20%花直径显著高于高亲,花被管长的超低亲现象达到了95%,而花序高的超低亲现象达到87.5%,大花萱草品种‘回复’和野生萱草品种‘北黄花菜’杂交后代的花梗长变异系数最大,变异系数最小的是‘北黄花菜’ב回复’的花被管长。任毅等[19]构建的43 个杂交组合中,变异系数最大的有苞片长、苞片宽及每葶蕾数。与花径呈极显著相关的性状有株高、花被花萼长、花被宽、苞片长、花被花萼色及花被管长,除苞片长外均为正相关。本研究采用黄花菜地方品种‘大同黄花’和萱草品种‘摇篮曲’进行杂交,结果表明,外瓣长度的变异系数最小,分离现象不明显,外瓣宽度和内瓣宽度的变异系数分别为12.39%和12.50%,表明这2 个性状在杂交F1 个体间的变异较大。杂交F1 群体花朵直径最大值为11.46 cm。花部性状大部分都介于父母本之间,呈现出较好的连续性正态分布趋势,推测5个花部性状均属于多基因控制的数量性状。杂交后代5 个花部性状的中亲优势值均达到极显著水平(P<0.01)。中亲优势值和中亲优势率均为负值,表明这5 个花部性状存在具有显性遗传效应的杂种优势,也表明萱草属植物具有高度杂合性,这在菊花[20]、柿[21]、苹果[22]和朱顶红[23]等植物中也有发现。除内瓣长度有2.81%为超亲个体外,其余性状皆未出现超亲个体,外瓣宽度和内瓣宽度均介于双亲之间,而花朵直径和外瓣长度相比杂交亲本表现出了一定的退化趋势。在相关性分析中,内瓣宽度与其余。个性状之间不相关,而花朵直径与外瓣长度、花朵直径与内瓣长度相关性紧密,说明主要是内外瓣长度决定了花朵直径的大小[24]。杂交后代的杂种优势源于杂交父母本间基因的异质性[23]。本试验中杂交后代群体存在广泛的分离和杂种优势衰退的现象,这些现象可能会导致杂种优势的利用受到限制[25],但却在杂交后代的单株优良性状选择方面带来了更加广阔的空间。
随着分子标记技术的发展,数量性状基因定位已经成为植物遗传育种研究中的有力手段,并在 观 赏 植 物 中 广 泛 应 用[10,11,26]。本 试 验 基 于 课 题组前期构建的种间杂交高密度遗传图谱,结合杂交后代5 个花部性状在2019 年的表型数据进行QTL 定位分析,共检测到15 个相关QTLs,分布在除1、6 和7 号连锁群的8 个连锁群上。各个QTL的LOD 值介于2.06~4.17 之间,可解释12.5%~23.7%的表型变异。将为今后的基因克隆和分子标记辅助育种技术在萱草属植物中的应用奠定重要基础。
4 结论
本研究对‘大同黄花’和‘摇篮曲’为亲本的杂交F1 代主要花部性状的遗传规律进行了分析。杂交F1 代中,外瓣宽度变异系数最大。5 个花部性状的中亲优势值均达到极显著水平(P<0.01)。花朵直径和外瓣长度表现出了一定的退化趋势。花朵直径与外瓣长度、花朵直径与内瓣长度、外瓣长度与内瓣长度相关性紧密,而内瓣宽度与其他各项指标之间不相关。本研究共检测到与萱草属植物杂交后代5 个花部性状相关的15 个QTLs,分布在8 个连锁群上。各个QTL 的LOD 值介于2.06~4.17 之间,可解释12.5%~23.7%的表型变异。对萱草属植物种质创新利用与品种改良具有重要意义,为今后萱草属植物花部性状相关基因发掘及候选基因筛选鉴定奠定了基础。
