百合叶片磷脂酶基因家族转录组学分析
2022-03-03葛金涛王江英汤雪燕腾年军朱朋波孙明伟赵统利邵小斌
葛金涛, 王江英, 汤雪燕, 腾年军, 朱朋波, 孙明伟, 赵统利, 邵小斌
(1.连云港市农业科学院,江苏连云港 222000; 2.南京农业大学园艺学院,江苏南京 210095)
百合是百合科百合属多年生草本球根植物,因其花姿雅致、叶片翠绿、茎秆挺直,常被作为鲜切花生产销售,目前已成为我国四大鲜切花之一,尤其在北方地区通过日光温室生产切花百合用于年宵花销售,具有较高的经济价值。在切花百合的生产过程中,部分品种由于缺钙会导致叶烧病的发生,其症状表现为在百合生长至30~40 cm,花蕾出现前,顶部6~10张新叶离叶尖2 cm处出现灰白色病斑,后期逐渐向叶尖扩展,最后变成焦枯状,有学者推测细胞膜的不稳定是导致叶烧病发生的原因。
细胞膜的主要成分是磷脂,磷脂构成的磷脂双分子层不仅是分隔细胞质与外界环境的屏障,还能够感受外界刺激,产生信号传递物质。磷脂酶(phospholipase)是生物体内负责磷脂代谢和生物合成的一类酶。磷脂酶根据其催化反应的对象分为5类,分别是磷脂酶A1(PLA1)、磷脂酶A2(PLA2)、溶血磷脂酶(PLB)、磷脂酶C(PLC)和磷脂酶D(PLD),在植物中主要为PLA1、PLA2、PLC和PLD这4类。磷脂酶几乎参与植物体所有的生命阶段,尤其在响应外界环境刺激中起着重要的作用。因此,研究其作用机制已成为近年来生物学重要且发展最迅速的领域之一。目前,磷脂酶基因已在许多植物中得到克隆和鉴定,但在百合中尚未开展相关研究。本研究利用Illumina测序对百合正常叶片和叶烧病叶片进行转录组测序分析,并通过数据筛选挖掘磷脂酶基因家族的基因信息,旨在为磷脂酶基因的功能研究奠定基础。
1 材料与方法
1.1 材料
试验采用百合品种Tarrango的18 cm周径种球种植于连云港市农业科学院东辛农场试验基地日光温室中,待百合长至50 cm左右,花苞从叶片中显露时,摘取百合植株同一部位正常叶片(TarCK)和叶烧病叶片(TarULN),放置于干冰保温盒中保存后送样。
1.2 Illumina测序与序列组装
高通量测序委托苏州金唯智生物科技有限公司完成。使用二代测序数据质量统计软件Cutadapt(version 1.9.1)对测序原始数据(Pass Filter Data)去除接头以及低质量序列等,后续信息分析用的Clean Data。采用组装软件Trinity对样品数据从头组装,组装结果通过序列聚类做进一步序列拼接和去冗余处理,得到长的非冗余unigene序列。
1.3 序列分析与功能注释
Unigene 序列通过Blast比对到蛋白质数据库NR、Uniprot、KEGG和COG,选择与给定unigene 序列同源性最高的蛋白,分别获得unigene 序列在4个数据库中的注释信息。
1.4 磷脂酶基因家族的筛选与数据分析
以磷脂酶(phospholipase)作为关键词,在获得的所有 unigene序列注释信息中搜索和筛选,得到磷脂酶基因家族候选基因。对所获候选基因在百合正常叶片(TarCK)和叶烧病叶片(TarULN)中的表达进行差异分析,同时按所获得的注释信息对候选基因进行分类,并预测候选基因参与的代谢途径。对获得磷脂酶基因序列以500 bp为单位做序列长度统计分析。
2 结果与分析
2.1 磷脂酶基因注释分析统计
在本次高通量测序过程中,共有274条Unigene序列被注释为磷脂酶基因(表1)。其中COG蛋白质数据库中得到注释为磷脂酶基因的unigene序列最多,为201条,其次为Uniprot和NR数据库中,分别为170条和126条,KEGG注释的unigene序列最少,为62条,在4个蛋白质数据库中都注释为磷脂酶基因的unigene序列有34条(图1),并且注释的磷脂酶基因类型一致,其中分泌磷脂酶基因4条,序列长度在500~1 000 bp之间,基因9条,基因24条(表2)。
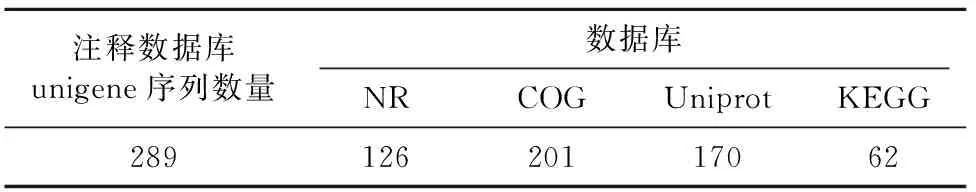
表1 磷脂酶基因注释分析统计 条

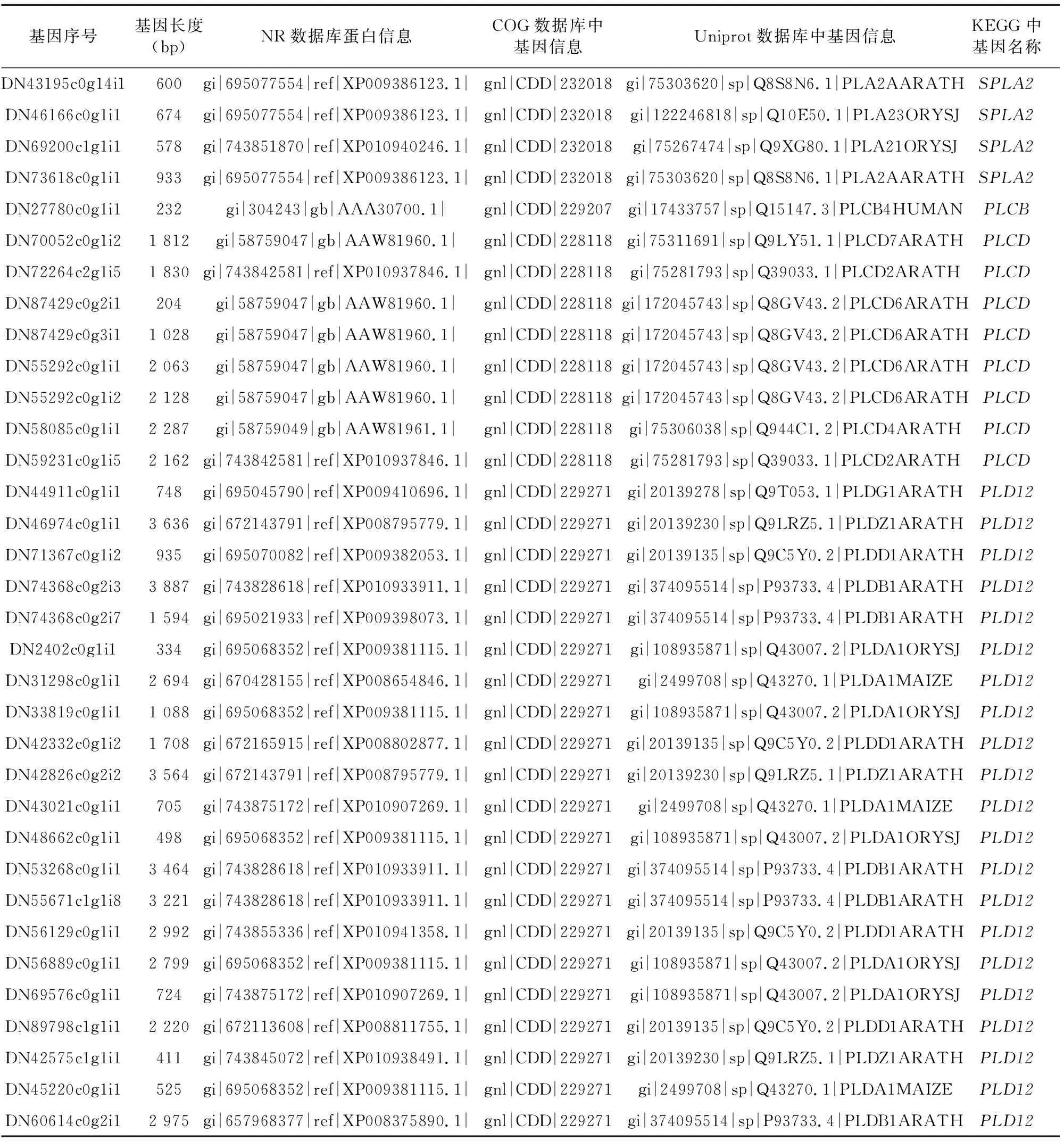
表2 在4个蛋白库都得到注释的磷脂酶基因
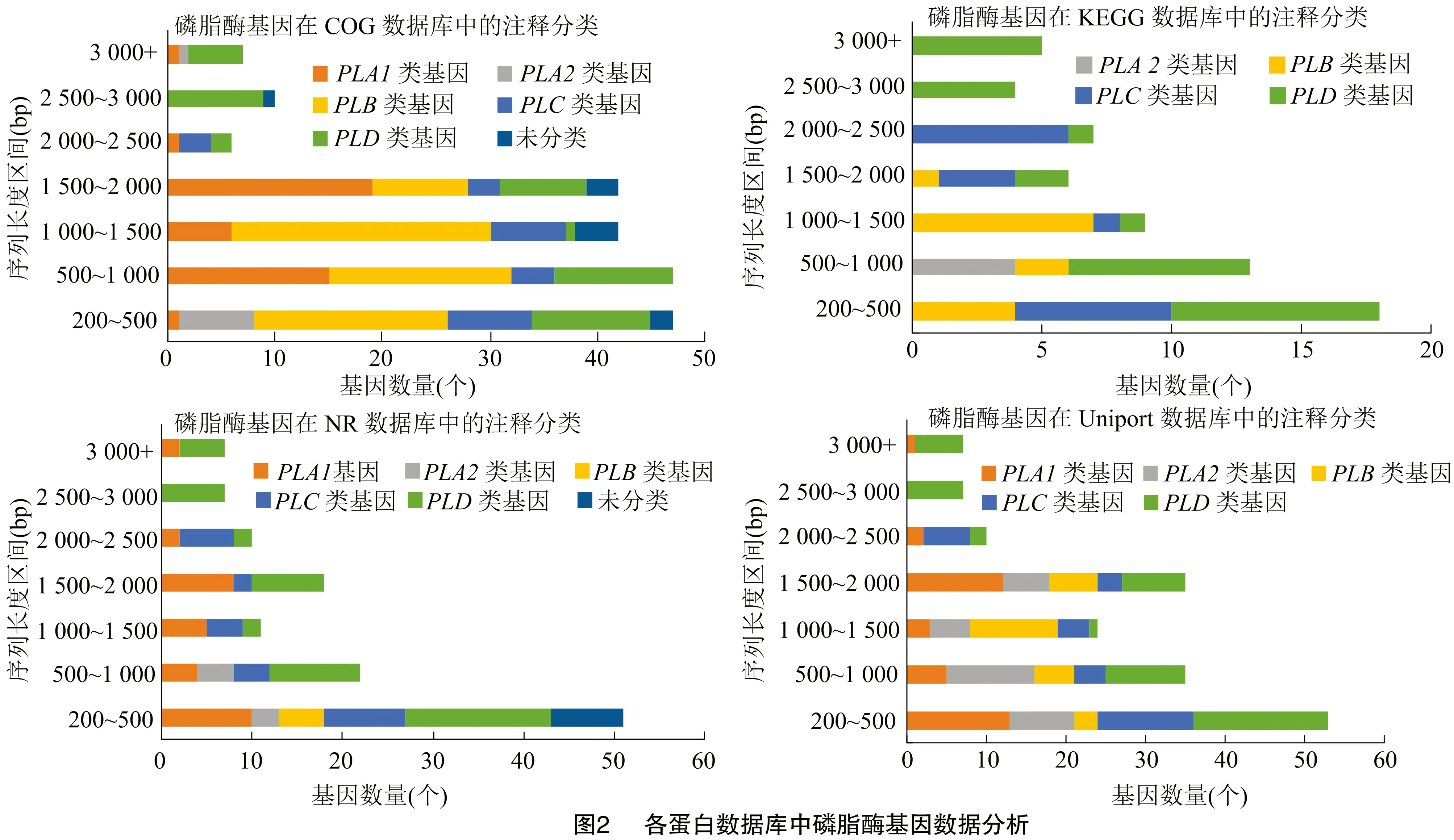
2.2 各蛋白数据库中磷脂酶基因的种类和数量分析
由图2可知,COG数据库中注释的磷脂酶基因最多,尤其是基因较多,经分析是COG数据库中注释为溶血磷脂酶基因(基因)的39个基因,在NR数据库中注释为咖啡酰莽草酯酶基因(caffeoylshikimate esterase)。有3个基因在COG数据库中注释为基因,而在NR数据库中注释为基因。通过比对NCBI数据库中的基因与基因发现,基因与基因具有很高的相似度,通过MEGA 6.06构建同源进化树分析,基因与部分基因的同源性高于基因之间的同源性(图3)。在KEGG数据库中有12个基因注释为lysophospholipaseⅡ(溶血磷脂酶基因LYPLA2),在Uniprot数据库中注释为acyl-protein thioesterase(酰基蛋白硫脂酶APT),利用MEGA 6.06构建百合与NCBI数据库中登录的APT基因的同源进化树,发现百合基因与非洲油棕、枣椰树和野芭蕉的基因同源性较高(图4),通过DNAMAN6.0软件多序列对比分析百合基因与非洲油棕、枣椰树和野芭蕉的基因同源性均高于83%。同时百合基因与NCBI中深圳拟兰、铁皮石斛和豇豆植物登录的基因同源性较高(图5),同源性均在73%以上。


通过比对4个蛋白数据库中注释基因发现,同基因一样与磷脂酶基因具有较高同源性的基因还有LYSOPL2、acylglycerol lipase、acyl-protein thioesterase、peroxiredoxin6,1-Cys peroxiredoxin和acriflavine resistance protein B(表3)。

表3 百合转录组测序中与磷脂酶基因具有较高同源性的基因

上述基因与磷脂酶类基因均具有较高的同源性,其中属于MGAT蛋白类似基因,包含1个水解酶或者乙酰基转移酶结构域和1个溶血磷脂酶结构域。一般属于多功能酶,同时具有乙酰基转移酶活性和水解酶活性。过氧化还原蛋白6 (peroxredoxin 6,Prdx6)是一种具有GSH过氧化物酶和磷脂酶A2活性的双功能蛋白。
2.3 代谢通路分析
对上述在4个蛋白库都得到注释的磷脂酶基因进行代谢通路分析(图6),基因参与了甘油磷脂代谢调控合成1-acyl-sn-glycero-3-phosphochine,参与醚脂类代谢调控合成1-alkyl-sn-gIycero-3-phosphocholine (lyso PAF)和1-(1-alkenyl)-sn-glycero-3-phospho ethanolamine (lysoplasmalogen),在百合叶烧病发生过程中,基因表达量上调。溶血磷脂酶基因基因参与了甘油磷脂代谢调控调控合成 sn-glycero-3-phosphocholine。基因在甘油磷脂代谢调控中调控合成1,2-diacyl-sn-glycerol,在醚脂类代谢调控中调控合成2-acyl-1-(lalkenyl)-sn-glycerol,在百合叶烧病发生过程中,基因表达量上调。基因参与了甘油磷脂代谢调控调控合成1,2-diacyl-sn-glycerol-3P,在醚脂类代谢中调控合成2-acyl-1-(lalkenyl)-sn-glycero-3-phosphate(plasmenic acid)。

3 讨论与结论
磷脂酶是催化磷脂水解释放游离脂肪酸的一类复杂而重要的酶,在植物体内分布广泛。其中PLAs根据序列同源性和进化关系一般分为3类:磷脂酶A1(PLA1)、patatin like磷脂酶(pPLA)和低分子量分泌磷脂酶A2(sPLA2)。也有学者将PLAs分为4类: 水解磷脂酰胆碱(PC)的PLA1、 偏爱磷脂酸(PA)的PLA1(PA-PLA1)、sPLA2和pPLA。在本次试验中百合的基因检测出基因(以为主)、和。在COG中有31条基因注释为的基因,而在NR数据库中31条基因均注释为-,同时在Uniprot数据中31条基因同时注释为patatin-like protein和patatin-related phospholipase A。这表明磷脂酶A(patatin-related phospholipase A,pPLA)与动物(钙离子非依赖型磷脂酶A2)为同源蛋白质,催化磷脂分子中sn-1和sn-2酰基酯键的水解,与Holk等的研究结果一致。
PLB是一种非常重要的代谢酶类,广泛存在于动植物及微生物体内,在植物中尚未见报道,具有水解酶、溶血磷脂酶和转酰基酶活性。水解酶和溶血磷脂酶活性是能够水解磷脂和溶血磷脂的 Sn-1和Sn-2位酯键,生成相应的甘油酰磷脂和脂肪酸,该酶在磷脂的分解代谢中具有非常重要的作用。Kegg数据库中,PLB基因家族分为12条基因注释为lysophospholipaseⅡ(LYPLA2 K06130)和 2条基因注释为pldB(K01048),即溶血磷脂酶基因的特异性较低,在注释时通常不能与其他脂酶基因区别,如咖啡酰莽草酯酶(caffeoylshikimate esterase)和单甘油酯脂肪酶(monoglyceride lipase),其原因是上述3类基因都具有溶血磷脂酶结构域,因此它们的基因序列相似度较高,基因与基因不能区分是因为多个基因兼具溶血磷脂酶活性和酰基蛋白硫脂酶活性。
PLC在植物内起到膜脂重构和胞内信号传导的作用,根据底物亲和力和细胞功能的差异分为磷脂酰肌醇特异性PLC(PI-PLC)和磷脂酰胆碱-PLC(PC-PLC),PI-PLC水解磷酸肌醇,产生肌醇1,4,5-三磷酸(IP3)和二酰甘油(DAG),可能是第2信使。IP3可以快速合成hexakisphosphate (IP6)并触发Ca内流,而DAG可以被DAG kinase (DGK)磷酸化并转化为磷脂酸(PAs)。与PI-PLC不同,PC-PLC也称为非特异性PLC (NPC),优先水解常见的膜磷脂,如PC、磷脂酰乙醇胺(PE)和磷脂酰丝氨酸(PS)。在本次试验中,磷脂酶C分为 PLCβ(K05858)和PLCζ(k05857)和plc(k01114)3类。phosphatidylinositol-specific phospholipase C1(PI-PLC)、non-specific phospholipase C1(NPC)和 non-hemolytic phospholipase C(非溶血性磷脂酶C)PLC-N与phospholipase C,phosphocholine-specific 一样。
磷脂酶D是一类能够水解磷脂质生成磷脂酸和羟基化合物的酶,其催化部位为磷脂质的磷酸二酯键。在百合中磷脂酶D分为磷脂酰肌醇-聚糖特异性磷脂酶D(phosphatidylinositol-glycan-specific phospholipase D)、糖基磷脂酰肌醇-磷脂酶D(glycosylphosphatidylinositol phospholipase D)GPI-PLD(EC:3.1.4.50);PLD3_4(EC:3.1.4.4)和PLD1_2(EC:3.1.4.4),其中PLD1_2又分为5类PLDα1、PLDζ1、PLDγ1、PLDβ1、PLDδ1。磷脂酶D基因结构特异性强,26条基因在4个数据库都得到注释。
植物体内存在的磷脂酶主要为PLA1、PLA2、PLC 和 PLD,在NCBI数据库中已检测到多种植物的基因序列。在本次转录组测序中,虽然有多个基因对到4个蛋白质数据库中,但都没有注释到任何代谢通路上。测序获得的基因与其他植物的溶血磷脂酶基因具有较高同源性,其功能分析需要进一步验证。百合磷脂酶候选unigene 的获得为进一步开展百合磷脂酶基因的克隆及功能鉴定奠定了基础,同时能为后续百合磷脂酶相关代谢途径及分子机制解析提供数据支持。
