野化训练中饵料源与放养密度对大黄鱼生长、消化和免疫活性的影响
2022-03-01李绅豪陈少波李鹏全刘伟成
李绅豪,陈少波,李鹏全,秦 松,刘伟成
(1.上海海洋大学,上海 201306;2.浙江省海洋水产养殖研究所,浙江温州 325005)
大黄鱼Pseudosciaena crocea 属硬骨鱼纲Osteichthyes、鲈形目Percifomes、石首鱼科Sciaenidae、黄鱼属Larimichthys,主要分布于东海、黄海南部以及南海的雷州半岛东侧水深80 m 以浅的沿岸近海水域,是暖水性近海鱼类[1]。20 世纪50—70 年代大黄鱼是东海最主要的经济鱼类之一。由于捕捞技术的高速发展,大黄鱼被过度捕捞,资源产量严重减少。1974 年中国大黄鱼产量为19.7×104t,而到20 世纪80 年代后期,总产量不足2.0×104t[2-4]。自1977 年起,大黄鱼产卵场不能形成渔汛,到20 世纪80 年代中期,大黄鱼最主要产卵场渔汛消失,大黄鱼剩余群体已经消亡殆尽。
增殖放流是大黄鱼资源修复的重要手段,但目前放流效果不佳,其原因主要包括以下三个方面:第一,人工配合饲料是鱼苗最主要的饵料,食谱单一,且投饲量过于充足,鱼苗捕食能力呈下降趋势;其次,集约化养殖严重压缩鱼苗的活动空间,鱼苗的运动机能下降;第三,鱼苗在养殖池中环境条件单一,缺乏生物饵料信息、捕食者信息和水环境变化信息,致使其野外适应能力下降[5-6]。除了鱼类自身的摄食需求,躲避天敌也是鱼类在自然界提高生存几率的重要能力之一。与养殖群体不同的是,野生鱼类在自然界中由于经常面对天敌的追捕,具有躲避天敌的经验与能力[7],而人工饲养的鱼类绝大多数都没有接触过天敌,缺乏面对天敌时的应对经验与能力,导致养殖的鱼类在自然界中很容易被天敌捕食[8-9]。因此,对鱼类进行野化训练是目前提高放流成活率的主要方法之一。孔彬等[10]的赤眼鳟Squaliobarbus curriculus 江河人工放流前适应性驯养试验表明,经过驯养后回捕率有较大提升。此外,对暗纹东方鲀Takifugu obscurus、罗非鱼Oreochromis mossambicus 等[11-12]相关研究显示,养殖饲料中某些成分可能会影响鱼类生长、免疫和消化能力,且不同的投喂率会直接影响养殖对象肠胃内饲料的存量。在不考虑饲料利用率的前提下,在一定范围内,投喂率越高,鱼的肠胃内饲料量越多,消化代谢水平越高[8]。如红鳍东方鲀Takifugu rubripes,在不同的投喂率下,胰蛋白酶活力、脂肪酶活力和肝脏丙氨酸氨基转移酶活力随饲料投喂水平增加而增加,而投喂率超过4%后,相关酶的活力没有显著性差异,且糜蛋白酶活力和血清尿素氮随投喂水平增加逐渐降低[13]。目前国内外有关大黄鱼的野化仍未见报道。本研究通过野化训练时改变大黄鱼的饵料源与放养密度,探讨不同生长模式对大黄鱼生长能力的影响,以期为今后大黄鱼资源恢复提供有效借鉴。
1 材料与方法
1.1 试验材料
试验鱼为浙江省海洋水产养殖研究所洞头基地人工培育的苗种(闽-粤东族大黄鱼)。选择30 000 尾鱼苗于海上鱼排的网箱中暂养2 周,以适应自然海域环境。暂养过程中,每天分别在06:00 和16:00 各投喂饲料1 次,投喂至表观饱食状态。实验地点位于浙江省温州市洞头区三排港海域。
本试验所投喂的饲料是福州海马饲料有限公司生产的海马牌鱼苗配合饲料,其主要成分包括进口鱼粉、淀粉、豆粕、鱼油、稳定型维生素与有机螯合矿物质。
1.2 试验方法
暂养2 周后,随机选取健康、大小均匀的初始体重为(1.65±0.34)g/尾的幼鱼15 000 尾,随机分为4 组,即对照组、高密度试验组、中密度试验组与低密度试验组。每组放养数量与密度分别为对照组1 800 尾、80尾·m-3,高密度试验组1 800 尾、80 尾·m-3,中密度试验组900 尾、40 尾·m-3,低密度试验组450 尾、20 尾·m-3。每组设3 个平行,分别饲养于规格为3 m×3 m×2.5 m 的试验网箱中,网箱孔径为5 mm×5 mm。在试验期间,保持试验组与对照组之间网箱距离足够远,防止试验组的灯光效应影响对照组,同时试验组停止人工配合饲料投入,每天18:00 将功率4 W 的充电式LED 灯泡放置于网箱底部,续航8 h,以灯光诱集生物饵料供幼鱼捕食,调查发现生物饵料主要为桡足类(爱氏门足戎Thyropus edwardsi、粗糙东刺糠虾Acanthomysis aspera li、丹氏纺锤水蚤Acartia negligens dana、普通波水蚤Undinula vulgaris、双刺唇角水蚤Labidocera bipinnata tanaka、汤氏长足水蚤C.thompsoni、微刺哲水蚤Canthocalanus pauper、异尾宽水蚤Temora discaudata、羽小角水蚤Pontellina plumata、中华哲水蚤Calanus sinicus、中隆水蚤O.media 等),试验中以浮游动物数量表述饵料丰度,饵料丰度为75 只·L-1。对照组每天分别在06:00 和16:00 各投喂饲料1 次,投喂至表观饱食状态,即养殖密度不影响鱼苗摄食强度。试验为期2 周。试验期间,水温为22.7~23.3 ℃,盐度为29.5~29.6,pH 为8.1~8.3,溶解氧为6 mg·L-1以上。
试验第7 天,每口网箱随机选取30 尾鱼,采用MS-222 麻醉,分别测定体质量、全长、体长、体高、体宽、尾柄宽等指标。试验第14 天,试验结束,停食24 h 后,从每口网箱中随机选取30 尾鱼,麻醉后立即低温运输至实验室,再次测定体质量、全长、体长、体高、体宽、尾柄宽等指标。同时,每份样品随机选取6 尾鱼在冰盘上解剖,分离肝脏和肠道组织,用4 ℃的生理盐水冲洗后,吸干表面水分,分别放置于5 mL 冻存管中,并迅速放置于液氮中速冻,于-80 ℃超低温冰箱保存,备测消化酶和抗氧化酶指标。
1.3 测定指标
1.3.1 测定指标1
生长指标包括增重率(WGR,%)、特定生长率(SGR,%·d-1)和肥满度(CF,%)计算公式如下:
式中:m0为试验开始时实验鱼的初始平均体质量,g;mt为结束时试验鱼的初始平均体质量,g;t 为试验天数,d;w 为体质量,g;L 为体长,cm。
1.3.2 测定指标2
抗氧化酶和消化酶指标主要选取溶菌酶(LYM)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、胃蛋白酶、α淀粉酶与脂肪酶等指标,六项酶活指标均采用南京建成生物工程研究所的试剂盒测定。
1.4 数据处理
数据采用Microsoft Excel 软件进行记录,结果采用SPSS 19.0 进行统计分析,主要采用单因素分析各试验因素之间的显著性,显著性水平设置为P<0.05。
2 结果与分析
2.1 野化训练对大黄鱼生长的影响
经过野化7 d 与14 d 后大黄鱼的生长情况如表1、2 所示,增重率与特定生长率如表3 所示。经过野化后,不同密度试验组的全长差异不显著(P>0.05),但是体长、提高、体质量、尾柄宽与体宽均表现为低密度试验组显著高于中密度试验组与高密度试验组(P<0.05),中、低密度试验组之间的差异不显著(P>0.05)。特定生长率与增重率表现为低密度试验组与对照组显著高于高密度试验组与中密度试验组(P<0.05),但是中、高密度试验组之间不存在显著性差异(P>0.05)。
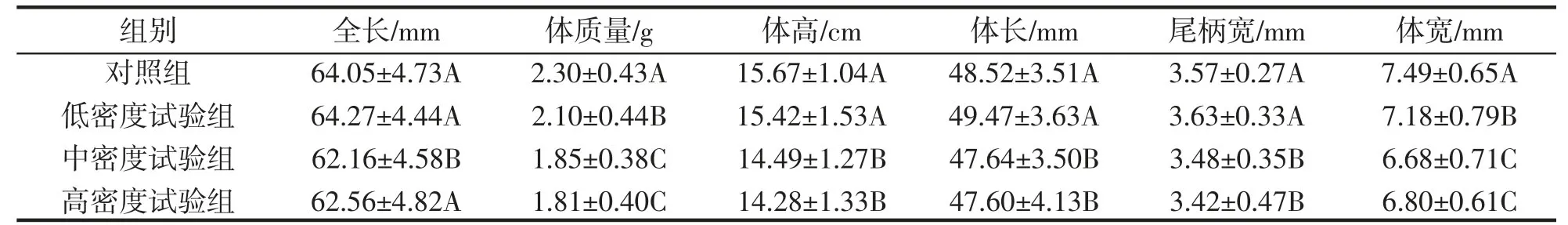
表1 野化训练7 d 后大黄鱼不同组间生长情况对比Tab.1 Comparison of growth among different groups after 7 days of wild training of P.crocea
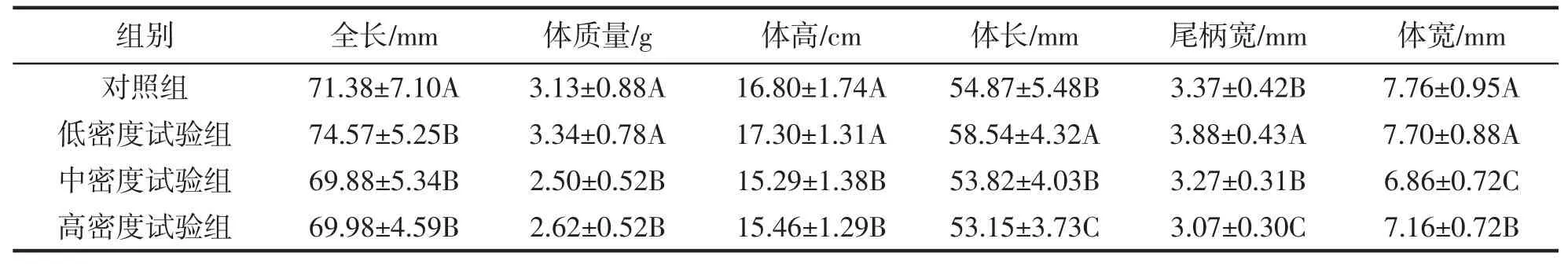
表2 野化训练14 d 后大黄鱼不同组间生长情况对比Tab.2 Comparison of growth among different groups after 14 days of wild training of P.crocea

表3 大黄鱼不同组间生长情况参数始末对比Tab.3 Comparison of growth between different groups in P.crocea
2.2 野化训练对大黄鱼体形指标的影响
不同组别间大黄鱼体形指标对比情况如表4 和表5 所示,实验组大黄鱼的体宽/体长、体高/体长和肥满度均低于对照组,尾柄宽/尾长除高密度实验组外其他2 个试验组均高于对照组。

表4 大黄鱼7 d 生理指标分析表Tab.4 Physiological index analysis table in P.crocea after 7 days
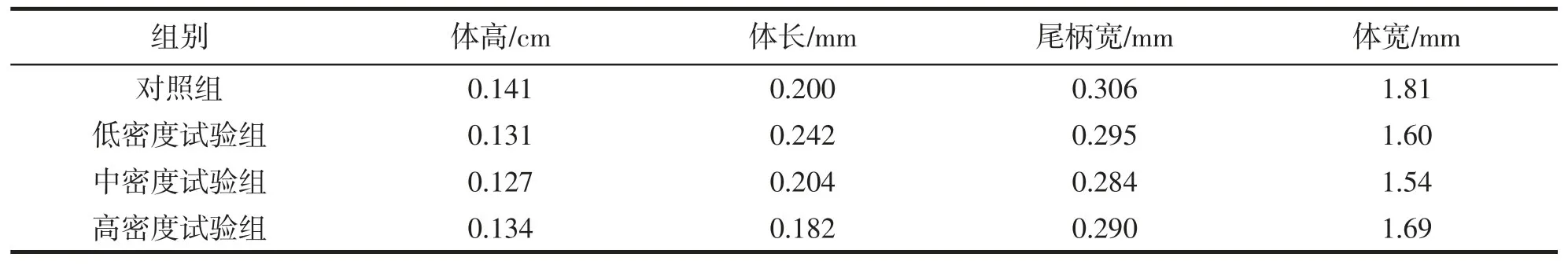
表5 大黄鱼14 d 生理指标分析表Tab.5 Physiological index analysis table in P.crocea after 14 days
2.3 野化训练对大黄鱼抗氧化酶与消化酶活力的影响
不同组别间大黄鱼肝脏中LYM、SOD、CAT 活性如表6 所示,肠胃中胃蛋白酶,脂肪酶与淀粉酶活性如表7 所示。在4 个组别中,LYM、SOD、CAT 的活性均表现为试验组>对照组,且除了SOD 的中密度组、高密度组及对照组外,均差异显著(P<0.05)。消化酶中除了淀粉酶外均表现为试验组>对照组,且除了胃蛋白酶的低密度组、中密度组及对照组以及脂肪酶的对照组、中密度组与高密度组外,其余均存在显著性差异(P<0.05)。
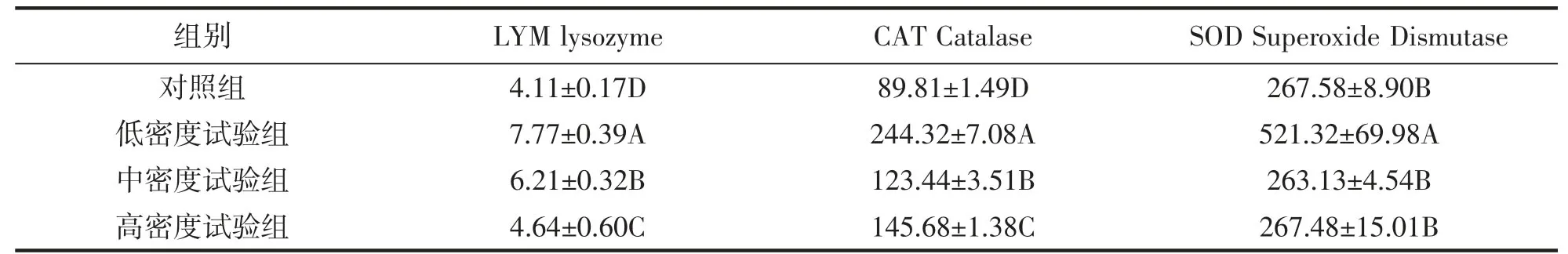
表6 大黄鱼不同组间免疫酶活性对比Tab.6 Comparison of immune enzyme activities among different groups in P.crocea

表7 大黄鱼不同组间消化酶活性对比Tab.7 Comparison of digestive enzyme activities among different groups in P.crocea
2.4 模型预测分析
在饵料丰度足够的前提下,依据野化试验结果,以饵料生物数量对比鱼苗数量及终体质量之间的关系建立模型模拟分析,结果如表8 所示。
对表8 中数据进行分析,可得知在一定条件下,鱼苗的终体质量与养殖密度成反比,与饵料生物密度正比。在低密度情况下,鱼苗终体质量为一稳定值,表明鱼苗摄食不受饵料供应限制,始终处于接近饱食的状态。饵料利用率在低养殖密度时随养殖密度的增加而增加,在高养殖密度时,饵料利用率趋向于稳定值。
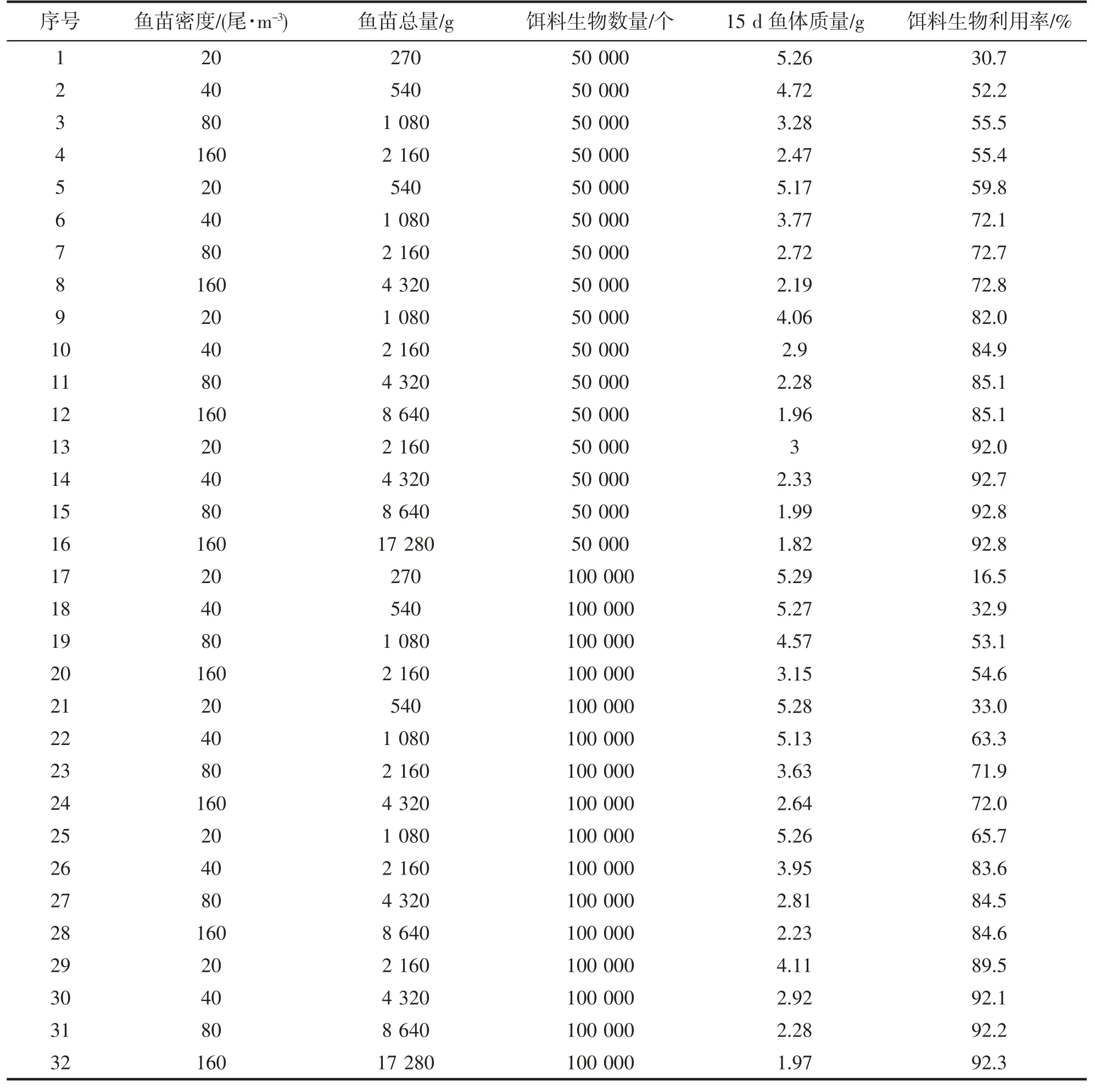
表8 饵料-鱼耦合野化模型在不同初始条件组合下系统运行考察指标的模拟结果Tab.8 Simulation results of system operation inspection indexes of shrimp-fish coupling domestication model under different initial condition combinations
3 讨论
3.1 摄食条件的改变对大黄鱼生长情况的影响
野化训练是提高水生生物放流成活率的主要技术手段之一,该技术是驯化的一个逆向过程,旨在恢复水生生物的自然习性,以适应自然水域[14]。有学者认为,野化训练能对鱼类的增殖放流起到指导作用[15],但是在食性与环境因子的野化选择上,仍需要通过长期的案例研究进行理论验证,同时需要大量实验证实野化训练观察指标的合理性,从而筛选和优化观察指标体系[16]。本试验中主要采用自然过渡和适度干预的原则开展,将室内人工繁育的大黄鱼苗种运输至自然海域网箱中暂养,以适应自然海域生态环境,同时停止人工饲料投喂,通过灯光诱捕饵料生物的技术手段,辅助大黄鱼完成从被动摄食到主动捕食的习性转化,从人工饲料向天然饵料的食性转化。有相关研究表明,个体越大的放流鱼类(如黑龙江鲟鱼)在自然环境中适应能力更强,捕食能力也更强,同时也就保证了其具有更大生存几率,提高了放流幼鱼的成活率[17]。本研究进行野化的大黄鱼鱼苗初始体长约44 mm、体质量1.65 g,野化14 d 后体长达到约55 mm、体重达到3.2 g,此阶段鱼苗属于幼鱼期。有调查显示,16~50 mm 的大黄鱼幼鱼主要摄食中华假磷虾、中华哲水蚤、糠虾类以及箭虫等,饵料尾长度平均为6.8 mm[18]。徐佳奕等[19]对大黄鱼进行的食物组成和摄食习性结果显示,大黄鱼主要摄食十足类(IRI=49.14%)、鱼虾类(IRI=18.31%)和糠虾类(IRI=2.34%)。在本试验中,海区内诱捕到的饵料生物基本满足大黄鱼摄食需要。其它对鱼类进行野化训练的相关研究表明,澜沧江土著鱼类叉尾鲇的野化训练后的生长形状无显著性差异[20],人工饲养倒刺鲃幼鱼经过15 d 的野化训练后的生长无显著差异,但是游泳能力显著提升[21]。在本研究中,经过训练后,试验组大黄鱼体质量优于对照组,体长显著性大于对照组(P<0.05),而硬骨鱼类的游泳速度与体长正相关[22],同时试验组大黄鱼特定生长率从1.30%~3.46%(7 d)
增长至2.98%~4.78%(14 d),而对照组从4.76%(7 d)下降至4.57%(14 d),说明经过训练大黄鱼具有更优的生长性能。对比训练7 d 与14 d 后大黄鱼的生理指标后发现,体高/体长、尾柄宽/体长、体高/体长与肥满度均为试验组低于对照组,此结果与王映等[23]在大黄鱼养殖群体和野生群体形态、鳞片及耳石特征比较中结果相一致,表明经过野化训练后试验组较对照组形体更优。
本研究显示,摄食天然饵料会对大黄鱼的生长有一定的促进作用,同时不同的放养密度也会影响到大黄鱼的生长,在低密度下大黄鱼的生长情况较好。
3.2 野化训练对大黄鱼抗氧化酶与消化酶活性的影响
SOD、CAT 等抗氧化酶是动物机体内消除氧自由基的重要酶,在机体的氧化与抗氧化平衡中发挥重要作用。SOD 是抗氧化系统重要成员之一,它不仅能清除体内多余的自由基,降解溶酶体内的细菌及异物,还在免疫中发挥重要作用[24]。SOD 可催化机体内的超氧化物经歧化反应转化为H2O 和H2O2,清除体内氧自由基,保护机体免受活性物质的损伤。CAT 可以加速物质的摄取和转运,形成水解酶体系,破坏和消除侵入体内的异物,达到机体防御的功能,是评价动物生理代谢水平的一项重要指标[25]。LYM 活性也是一种重要的非特异性免疫的指标,良好的LYM 活性有助于鱼类抵抗水域中各种病原物的侵袭[6]。T-AOC 是动物总抗氧化能力的综合性指标,是机体抗氧化酶系统和抗氧化系统共同作用的结果,能够反映机体对外界刺激的代偿能力以及清除活性氧自由基的综合能力[25]。一些研究表明,在环境中盐度发生变化时,银鲳Pacpus argenteus、钝吻黄盖鲽Pseudopleuronectes yokohamae 等鱼类SOD 活性升高,使得2 种H2O2清除酶CAT 和GPX 的反应活性升高[25]。在本研究中,对大黄鱼肝脏中的几种酶活性进行测定后发现,试验组中大黄鱼肝脏组织LYM、CAT 与SOD 活性活力均高于对照组,说明通过改变摄食的饵料类型,大黄鱼的免疫与应激能力强于投喂人工饵料的大黄鱼。
有相关研究表明鱼类体内的消化酶活性由食性决定,且其变化程度与食物的组成有一定的相关性。如肉食性鱼类含有较高的脂肪酶和蛋白酶活性,而杂食性鱼类比肉食性鱼类表现出较高的淀粉酶活性,在杂食性鱼类中,淀粉酶活性高于蛋白酶活性。一般情况下,肉食性鱼类蛋白酶活性最高,草食性鱼类的蛋白酶活性最低,其蛋白酶活性由强到弱的顺序为:肉食性鱼类>杂食性鱼类>滤食性鱼类>草食性鱼类[26]。本研究结果显示,试验组消化酶中胃蛋白酶与脂肪酶活性均高于对照组,淀粉酶活性低于对照组,说明经过训练后大黄鱼食性更偏向于肉食性鱼类,有利于提高大黄鱼的捕食适应能力。
总体而言,摄食天然饵料生物的试验组大黄鱼具有更优的抗氧化能力与免疫能力,且食性更偏向于肉食性,因此说明摄食天然饵料可以提高大黄鱼的适应能力。
3.3 放养密度对大黄鱼生长与免疫消化能力的影响
放养密度是一项影响鱼类生长的重要环境胁迫因子。随着密度的增加,单位体积内的溶氧、饵料、生存空间等因素都会发生改变。一些研究表明龙纹斑幼鱼、银鲳幼鱼与杂交鳢等生长密度会影响鱼类生长,表现在一定范围内随着野化训练密度的上升,生长与免疫消化水平下降[27-29]。龙斑鱼的密度实验表明,养殖密度最终会影响到龙斑鱼幼鱼的体质量、日增重率与特定生长率等指标,两者成反比关系,且在降低到一定密度之后影响开始变小,直至忽略不计。高放养密度对龙纹斑幼鱼的生长存在着消极影响,而低密度的放养效果也不好[28]。对银鲳幼鱼的研究表明适当提升放养的密度可以改善银鲳幼鱼的生长情况,但在高密度放养时,银鲳幼鱼则会受到拥挤胁迫的影响,肝脏、肾脏中相关酶活力显著升高[29]。一些学者对锦鲤Cyprinus carpio 的研究发现,提高养殖密度,会导致虹鳟Oncorhynchus mykiss 的溶菌酶活性快速下降,并在继续升高密度后维持较低水平[30]。而对杂交鳢的研究结果表明,高密度放养会增加机体能耗,促进氧自由基的产生,最终导致丙二醛含量升高[27]。对俄罗斯鲟Acipenser gueldenstaedti 的结果显示,随着养殖密度的上升,SOD 活性下降[31]。对中华鲟Acipenser sinensis 的结果显示,高密度下,SOD 活性下降[32]。在本研究中,低密度放养时大黄鱼生长情况更优,总体表现为低密度试验组>中密度试验组>高密度试验组,且各组各项指标之间均存在显著性差异(P<0.05)。大黄鱼高密度试验组SOD 活性较中密度试验组与低密度实验组低,说明放养密度过高也会引起机体代谢失衡,造成细胞损伤,降低了机体的抗氧化能力。总体而言,不同放养密度下的大黄鱼生长与消化酶及抗氧化酶活性指标存在差异,表现为随着放养密度的上升,生长与胃蛋白酶及抗氧化酶活性下降。
3.4 最适养殖密度分析
整体而言,鱼苗终体质量与饵料的利用率在一定程度上呈负相关,饵料生物被充分捕食而利用率较高时,鱼苗通常不能饱食而导致终体质量降低;鱼苗饱食而终体质量较高时,饵料生物由于未被充分摄食而导致利用率较低而产生浪费。鱼苗终体质量反映的是产品质量指标,而饵料生物利用率反映的是系统经济性指标,寻找二者之间的平衡点,在确保一定产品质量的情况下达成最高经济性的初始条件组合即为最优组合。根据以上初始条件对饵料生物利用率及鱼苗终体重的影响的分析,可判断出二者同时呈高值时的初始条件应为养殖密度较低而网箱数量较高的情况。将不同初始条件组合下的鱼苗终体重对饵料生物利用率作图,结果如图1 所示。
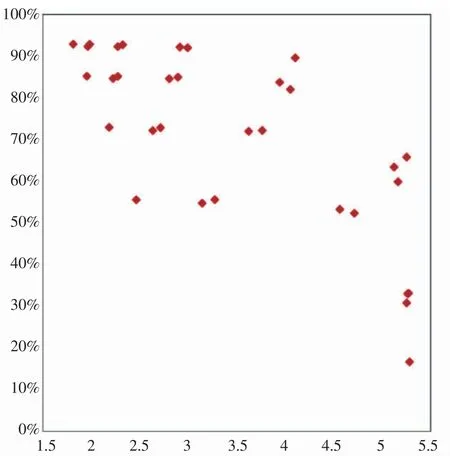
图1 鱼苗终体质量与饵料利用率的相关关系Fig.1 Correlation between fish fry weight and feed utilization rate
若设定鱼苗终体重>4 g 为终鱼苗的质量标准,则图1 中右上方的3 个数据点可满足产品质量与资源利用效率均较优的要求,此3 个数据点的初始条件组合分别为表8 中的#9、#26 和#29 号组合,即鱼苗密度20+饵料生物数量50 000、鱼苗密度40+饵料生物数量100 000、鱼苗密度20+饵料生物数量100 000 这3 个组合。这其中鱼苗密度20+饵料生物数量100 000 的组合为最优,而3 个组合的共同点在于投放饵料生物数量与养殖鱼苗总数量之比相同,均为约50:1。
整体而言,对大黄鱼进行野化的最优密度应为:饵料/鱼数量比约50:1。
4 结论
本研究进行大黄鱼摄食试验,通过对结果的分析与讨论,可以得出如下结论:
(1)通过改变摄食天然饵料,大黄鱼苗种生理指标均有显著性的增长。摄食自然饵料的大黄鱼特定生长率优于人工投喂的大黄鱼,且体长显著大于人工饵料投喂组。
(2)摄食天然饵料后,试验组大黄鱼的抗氧化酶活性指标更优,同时食性更偏向于肉食性,表明摄食训练后大黄鱼适应能力会有提高。
(3)放养密度不同会影响大黄鱼的生长。生长情况与放养密度成负相关,即放养密度越低,生长情况越好。
(4)最适放养密度应选择环境中饵料生物为放养大黄鱼密度的50 倍。
因此认为,通过摄食天然饵料与降低生长密度的方式可以提高大黄鱼的生存适应能力,从而提高大黄鱼的放流成活率。
