基于根伸长测试终点的小白菜Cd 及Se-Cd 物种敏感性分布
2022-02-27李湘凌张忠恩卢新哲魏迎春
胡 翔,李湘凌,2*,张忠恩,赵 燕,卢新哲,魏迎春
1.合肥工业大学资源与环境工程学院,安徽 合肥 230009
2.安徽省矿产资源与矿山环境工程技术研究中心,安徽 合肥 230009
3.自然资源部平原区农用地生态评价与修复工程技术创新中心,浙江 杭州 311200
4.浙江省地质调查院,浙江 杭州 311200
Cd(镉)是一种广泛存在于土壤环境中的有毒重金属[1-2],可导致植物生物量生产、养分获取、抗氧化酶活性和光合作用等生理过程恶化[3-5],同时易累积在植物可食用部分并进入食物链危害人类健康[6-8].2014 年《全国土壤污染状况调查公报》显示,Cd 为我国农田土壤点位超标率最高的重金属[9],以暴露剂量-效应为核心的Cd 生态风险评价受到了广泛关注[8-11].Se(硒)是人类必需的微量营养元素,摄入适量Se 可提高人体免疫能力,发展富Se 农产品产业成为农业结构调整的重要措施[12],而富Se 环境下的Cd 毒性变化是富Se 农产品产业发展必须考虑的因素.现有研究表明,Se 对Cd 毒性影响具有双重效应,一方面Se能抑制植物体内的过氧化作用,恢复细胞膜结构和功能的完整性[13-14],降低重金属引起的氧化应激[15],减轻重金属Cd 对植物的生理毒性[16-17];另一方面Se 会破坏细胞膜增强Cd 渗透作用,导致Cd 毒性加剧[18-19].但是,关于Se 影响下的Cd 剂量-效应关系及毒性阈值的研究鲜有报道,因此加强Se-Cd 复合作用下的相关研究对农产品的Cd 生态风险控制具有积极意义.
小白菜(Pai-Tsai,Brassica chinensisL.)是我国种植面积大、产量高、经济价值高的叶类蔬菜之一,其品种多样,不同品种间Cd 毒性效应差异较大[20-21].考虑到种子萌发期是小白菜等植物最早受环境胁迫的阶段[10,22-23],且由于根直接与污染物接触、对污染物敏感性较强,其直接影响后期植物生长、生物量和品质,因此众多研究者将根伸长作为污染物毒性效应研究的表征终点[3,24-25].该研究以种子萌发期根伸长为表征终点开展Cd 暴露剂量-效应关系研究,可有效筛选出小白菜Cd 耐性品种,为小白菜的种植提供参考.
物种敏感性分布 SSD (species sensitivity distribution)常用于水体环境[26-27]、整个生态系统[28]或同一物种的不同品种[29]的生态风险阈值研究,该方法通过概率分布模型将不同物种(品种)的效应浓度进行合理外推,确定对环境中95%物种(品种)具有保护作用的最大环境有害浓度HC5(hazardours concentration for 5% of species)[30-31].现有研究主要采用Log-logistic、Log-normal 和Burr-Ⅲ等分布模型进行SSD 曲线拟合,其中Burr-Ⅲ分布模型因具有灵活性、拟合特性较好等优点被广泛应用[30-33].利用SSD不仅能够有效估算小白菜的Cd 毒性阈值,而且可以探究Se 对小白菜Cd 毒性阈值的影响,弥补相关研究的不足,为富Se 环境下的Cd 毒性阈值确定提供参考.
该研究采用国际种子检验协会测试协议中的培养皿滤纸法[34],选取种子萌发期的根伸长作为测试终点,探究Cd 及Se-Cd 对11 种小白菜萌发期根伸长的毒性效应,采用Log-logistic 模型及低剂量Hormesis兴奋效应模型进行剂量-效应曲线拟合,得到不同比例效应浓度数据,并基于Burr-Ⅲ分布模型外推得到小白菜SSD 曲线和毒性阈值.通过研究可确定小白菜的Cd 及Se-Cd 暴露剂量-效应关系,筛选出Cd 耐性品种,以期为小白菜Cd 生态风险评价及控制提供理论依据.
1 材料与方法
1.1 供试材料
供试试剂:分析纯氯化镉(CdCl2·2.5H2O)、亚硒酸钠(Na2SeO3).
供试小白菜:AJH(矮脚黄)、BBQ(八宝青)、HYG(黑油冠)、GGB(高梗白)、SYM(四月慢)、SG259(苏冠259)、YDE(油冬儿)、SHQ(上海青)、SZQ(苏州青)、KR(抗热)和NY(奶油) 11 种小白菜种子,购于合肥市合丰种业有限公司等多个蔬菜种子公司.
1.2 试验方法
该研究采用培养皿滤纸法[34]分别开展Cd 及Se-Cd 作用下11 种小白菜萌发期的根伸长试验,并根据实际情况稍做修改.
Se、Cd 浓度设置:根据预试验结果确定适宜Se浓度为0.6 mg/L,设置11 个Cd 浓度梯度分别为0、0.1、0.5、1、2、5、10、20、30、40、60 mg/L.Cd 处理组分别添加11 个浓度Cd,Se-Cd 处理组分别添加11 个浓度Cd 和适宜浓度Se (0.6 mg/L),其中,对照样品(CK)不添加Cd 和Se.每个处理设置3 个重复.
试验过程:将种子用去离子水洗净,3% H2O2浸泡30 min 消毒和打破种子休眠,再用去离子水进行洗净、浮选处理;然后用镊子将饱满种子均匀播种在培养皿中的滤纸上,播种粒数为30 粒,再加入Se、Cd 溶液,盖好玻璃培养皿,置于人工气候培养箱(RGX-350ES,350 L,河北博汇仪器有限公司)中避光培养(温度26 ℃,相对湿度75%).培养7 d 后[34-35],用游标卡尺测定根长度,计算不同Cd 及Se-Cd 处理水平下的平均根伸长,并以对照样品(CK)的根伸长为基准,计算相对根伸长RRE (relative root elongation).
1.3 剂量-效应曲线拟合
一般情况下,采用Log-logistic 模型拟合小白菜Cd 毒性的剂量-效应关系曲线,拟合方程:

式中:Y为不同处理水平下的小白菜相对根伸长,%;X为Cd 浓度,mg/L;y0和b为模型参数;M为EC20Cd或 EC50Cd的自然对数值,其中EC20Cd和EC50Cd分别为20%和50%比例效应浓度,该研究中,即以对照样品CK 的根伸长为基准,小白菜RRE 抑制水平为20%和50%时对应的Cd 浓度.
当土壤中低浓度Cd 对植物产生较强兴奋效应时,采用低剂量Hormesis 兴奋效应模型进行毒性效应拟合[36],拟合方程:
乡土植物资源是城市植物多样性最重要的组成部分,对于森林群落来说,乡土树种的占比越高,其适应性、稳定性和抗逆性就越好,同时能维持自身的营养平衡,保持自然更新[13]。本研究中乡土树种根据《上海植物志》和《华东五省一市植物名录》确定[11-12]。

式中,a、b、c、d为模型参数,k为效应比例.当k值为20 或50 时,c为相应的效 应比例浓度 EC20Cd或EC50Cd.
1.4 物种敏感性分布曲线拟合
根据1.3 节中式(1)(2)得到的EC20Cd或 EC50Cd,采用Burr-Ⅲ分布模型拟合得到小白菜Cd 物种敏感性分布曲线,分别计算得到保护95%小白菜品种的Cd毒性阈值HC5(基于 EC20Cd和 EC50Cd的Cd 毒性阈值分别为 HC520和HC550),Burr-Ⅲ分布模型[29,37]:

1.5 数据处理与分析
使用Microsoft Excel 2016、Origin 2018 软件进行数据整理及图件制作,使用SPSS 21.0 软件进行单因素方差分析及多重比较.
2 结果与分析
2.1 Cd 及Se-Cd 作用对小白菜根伸长的影响
Cd 处理组中,11 种小白菜的RRE 见图1(a).品种YDE、HYG 和SZQ 的RRE 随着Cd 浓度增加,呈现先上升后下降的趋势,在Cd 浓度为0.1 mg/L 时RRE 最 大〔YDE (102.7%)、HYG (102.7%)和SZQ(103.3%)〕,低浓度Cd(≤1 mg/L)对萌发期根伸长具有刺激抗逆性,可能与小白菜胚乳液化和分解能力增强,暂时产生了积极作用有关[23].品种AJH、SHQ、SG259、SYM、GGB、NY、BBQ、KR 的RRE 随着Cd浓度增加而降低.显著性检验表明,部分品种在特定Cd 浓度范围内(AJH 中Cd 浓度为5~60 mg/L,YDE、HYG、SG259、BBQ 和KR 中Cd 浓度为1~60 mg/L,SZQ、SYM 和GGB 中Cd 浓度为2~60 mg/L)的RRE均显著低于CK,而品种SHQ、NY 在全部Cd 浓度下的RRE 均显著低于CK.
Se-Cd 处理组中,11 种小白菜的RRE 见图1(b).全部品种小白菜的RRE 随着Cd 浓度增加均呈现先上升后下降的趋势,其中,品种YDE、SHQ、SG259、SZQ、SYM 和KR 在Cd 浓度为5 mg/L 时的RRE 最大,品种AJH、HYG、GGB、NY 和BBQ 在Cd 浓度为10 mg/L 时的RRE 最大.与Cd 处理组比较,Se-Cd 处理组RRE 峰值的增幅明显,其中YDE、HYG 和SZQ 的RRE 峰值差为32.6%~40.9%,SG259、SYM 和BBQ 的RRE 峰值差为6.5%~9.2%,其余品种的RRE 峰值差为10%~15%.

图1 不同浓度水平Cd 及Se-Cd 处理下小白菜相对根伸长Fig.1 Relative root elongation response of Pai-Tsai to different concentrations of Cd and Se-Cd
Cd 处理组和Se-Cd 处理组的RRE 随着Cd 浓度变化具有明显的差异,在较低Cd 浓度(AJH、YDE、GGB、NY、BBQ、KR、SYM 中Cd 浓度≤20 mg/L;HYG、SZQ 中 C d 浓度≤30 mg/L;SHQ、SG259 中Cd 浓度≤10 mg/L)作用下,Cd 处理组的RRE 均显著小于Se-Cd 处理组;在较高Cd 浓度作用下,Cd 处理组中大部分品种的RRE 大于Se-Cd 处理组.
2.2 Cd 和Se-Cd 对小白菜根伸长毒性的剂量-效应关系
基于Log-logistic 模型和低剂量Hormesis 兴奋效应模型,分别拟合得到11 种小白菜RRE 在Cd 及Se-Cd作用下的Cd 剂量-效应曲线(见图2).在Cd 处理组中,品种YDE、HYG 和SZQ 的Cd 兴奋效应较低(RRE<105%),基于低剂量Hormesis 兴奋效应模型和基于Log-logistic 模型得到的拟合曲线基本相同,该现象与孙聪等[29,36]等研究结果相似.

图2 不同品种小白菜的Cd 及Se-Cd 剂量-效应关系曲线Fig.2 Cd and Se-Cd dose-effect relationship curves of Pai-Tsai cultivars
与Cd 处理组比较,Se-Cd 处理组所有品种小白菜都具有更明显的Cd 兴奋效应.Cd 处理组(YDE、HYG 和SZQ)中最大兴奋效应Cd 浓度为0.1 mg/L,Se-Cd 处理组中最大兴奋效应Cd 浓度提高为5~10 mg/L.但当Cd 浓度超过兴奋效应浓度后,Se-Cd 处理组中多数品种的RRE 迅速降低,其毒性效应超过Cd处理组,该规律在品种NY 和KR 中尤其突出,但品种SG259 (Cd 浓度为30~60 mg/L)和SYM (Cd 浓度为60 mg/L)受高浓度Cd 作用时,Se-Cd 处理组的毒性效应却低于Cd 处理组.
基于剂量-效应关系拟合曲线,得到不同品种小白 菜的Cd 效 应比例浓度 EC20Cd和 EC50Cd(见表1).Cd处理组中,品种间 EC20Cd和 EC50Cd差异较大,品种SZQ的EC20Cd(25.10 mg/L)和 EC50Cd(40.25 mg/L)最大,分别为品种NY的EC20Cd(2.03 mg/L)和EC50Cd(8.03 mg/L)的12.36 和5.01 倍.Se-Cd 处理组的品种间EC20Cd和 EC50Cd差距小于Cd 处理组,品种SZQ 的EC20Cd(30.12 mg/L)和EC50Cd(48.45 mg/L)最大,分别为品种SG259 的EC20Cd(8.77 mg/L)和 EC50Cd(14.74 mg/L)的3.43 和3.29 倍.
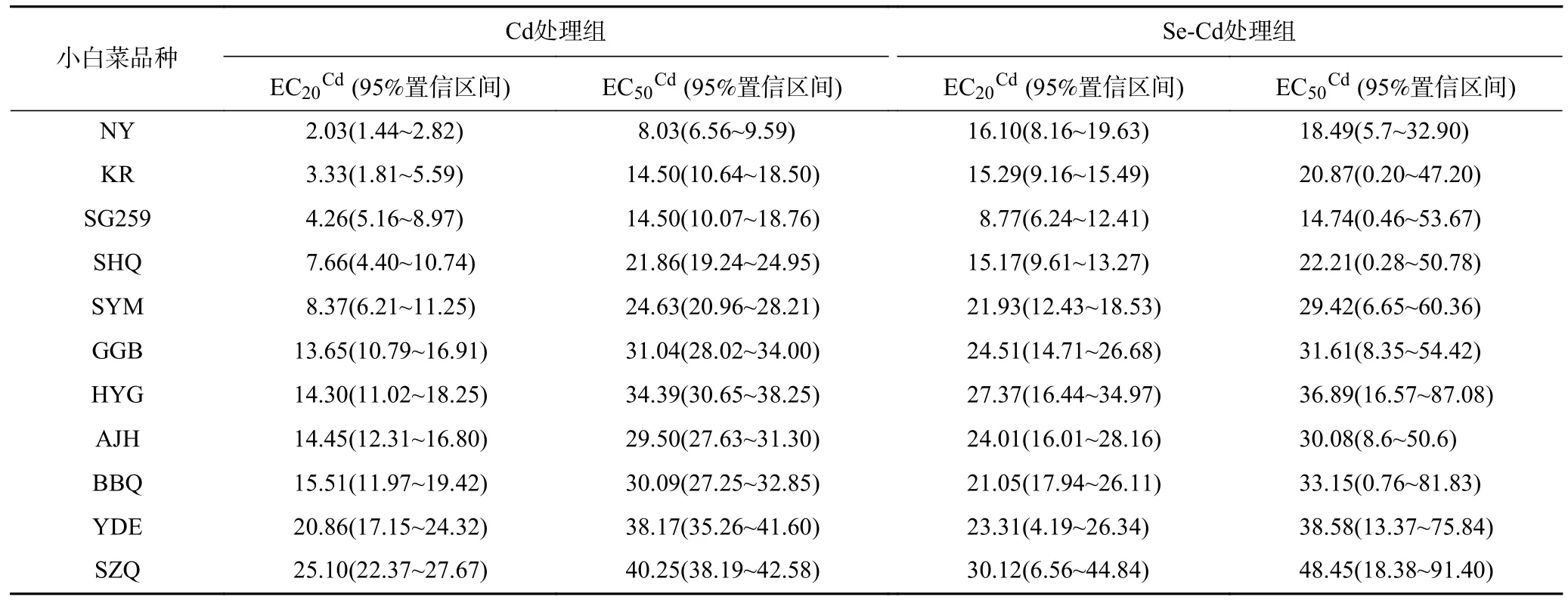
表1 Cd 及Se-Cd 处理组小白菜相对根伸长的不同比例效应浓度EC20Cd和 EC50Cd Table 1 EC20Cd and EC50Cd of Pai-Tsai cultivars based on relative root elongation response to Cd and Se-Cd mg/L
对比Cd 处理组与Se-Cd 处理组的结果表明,适宜浓度Se 可不同程度地提高小白菜的EC20Cd和EC50Cd,其中品种NY的EC20Cd提高了7.9 倍,EC50Cd提高了2.3 倍,而SZQ 和YDE 的EC20Cd和EC50Cd仅提高了1.2 倍左右.
综上,适宜浓度Se 对小白菜Cd 毒性的影响具有双重性,一方面Se 可缓解低浓度Cd 对小白菜的毒性效应,提高Cd 兴奋效应;另一方面Se 加剧了高浓度Cd 的毒性效应,Se 可有效提高小白菜的不同比例效应浓度 EC20Cd和 EC50Cd.
2.3 小白菜物种敏感性分布及HC5 值预测
基于小白菜EC20Cd、EC50Cd得到Cd 及Se-Cd 处理组的拟合SSD 曲线(见图3).结果表明,品种SZQ、YDE、HYG 表现出较强的Cd 耐性,品种NY、SG259、KR 对Cd 较为敏感.图3 显示,效应比例(即抑制水平)和添加Se 均能明显影响小白菜Cd 敏感性分布.
基于不同比例效应浓度(EC20Cd或EC50Cd)得到的小白菜Cd 敏感性排序不同.在不同效应比例下,Cd处理组中的Cd 敏感性品种NY 和耐性品种SZQ、YDE 的敏感性排序基本相同,其余品种的敏感性排序均产生了变化.Se-Cd 处理组中,除品种SG259 和耐性品种SZQ 外,其他品种在不同效应比例下的敏感性排序不同.
Se 不影响Cd 耐性品种SZQ 的敏感性排序,导致Cd 敏感性品种NY 的敏感性排序后移,对Cd 较强耐性品种HYG 和YDE 的敏感性影响较复杂.添加Se 对基于 EC20Cd的小白菜品种敏感性排序〔见图3(a)(b)〕的影响大于基于 EC50Cd的敏感性排序影响〔见图3(c)(d)〕.
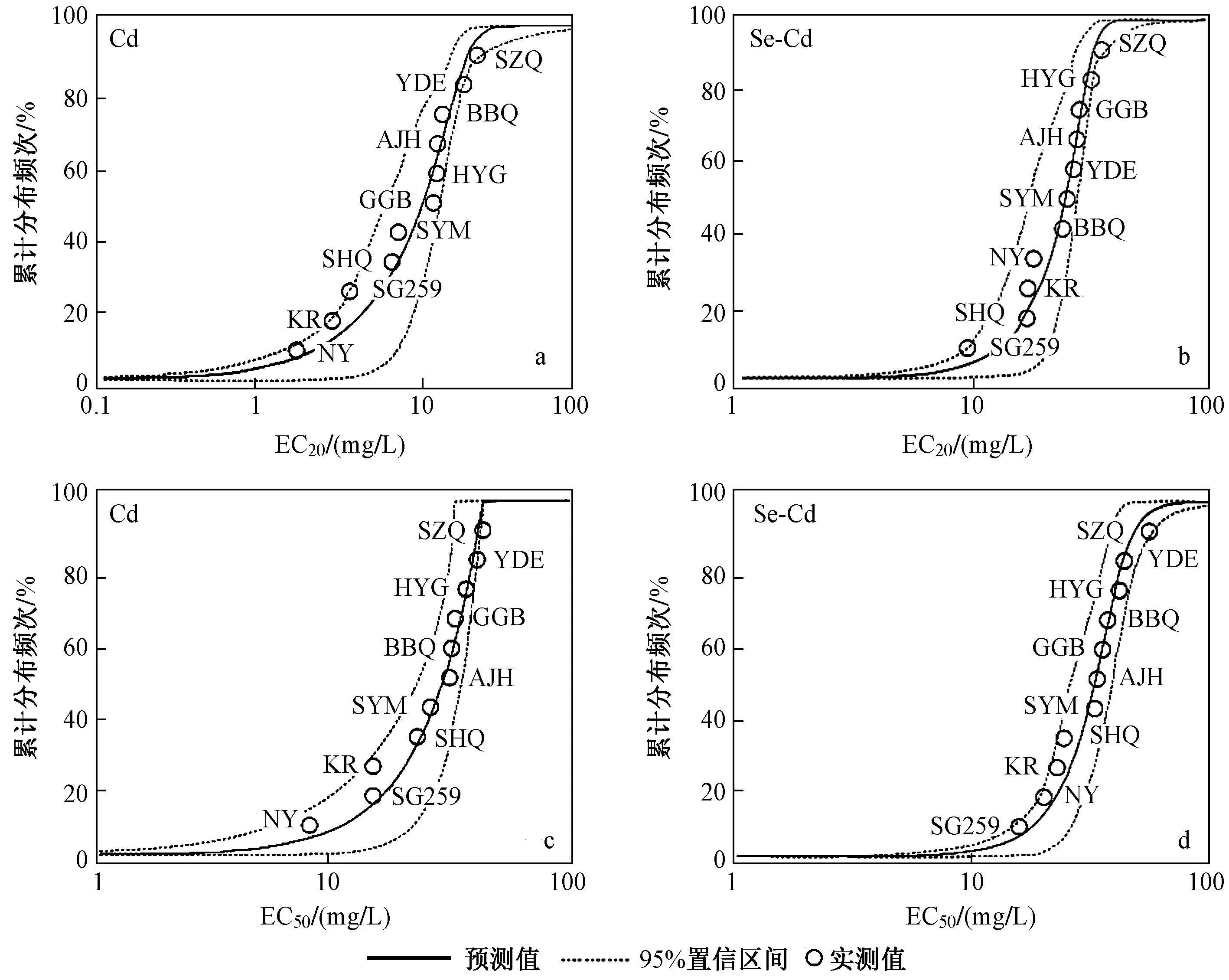
图3 不同小白菜品种基于EC20Cd或EC50Cd 的物种敏感性累计分布频次曲线Fig.3 Cumulative frequency of species sensitivity distributions of Pai-Tsai cultivars based on the EC20Cd and EC50Cd
基于EC20Cd和EC50Cd,采用Burr-III 分布模型分别拟合得到毒性阈值HC520和HC550(见表2).Cd 处理组的HC520和HC550分别为2.94 和11.71 mg/L,Se-Cd 处理组的HC520和HC550分别12.00 和16.28 mg/L.添加适 宜浓度Se,HC520提高了约4.0 倍,HC550提高了约1.4 倍,明显提高了对95%小白菜品种具有保护作用的最大Cd 浓度.
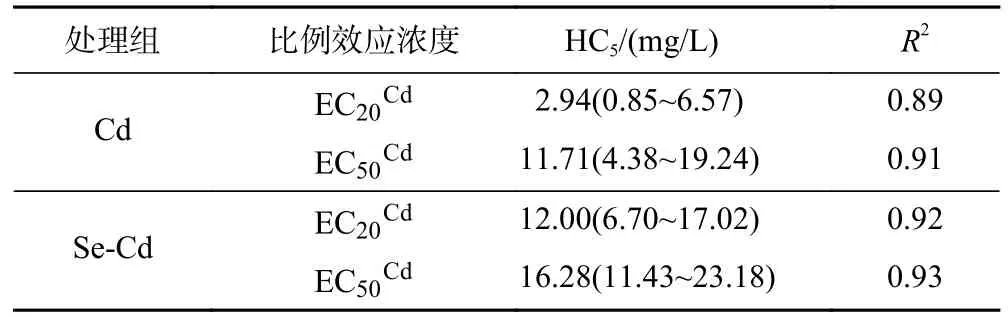
表2 基于Burr-Ⅲ分布模型拟合的保护95%小白菜品种Cd 的毒性阈值(HC5)Table 2 Hazardours concentration for 5% of species (HC5) of Cd to Pai-Tsai determined with Burr-Ⅲ model
综上,基于不同效应比例会影响小白菜Cd 敏感性顺序,适宜Se 浓度会不同程度影响Cd 对小白菜RRE 的敏感性顺序,且有效提高了保护95%小白菜品种不受到Cd 胁迫的毒性阈值.
3 讨论
3.1 Cd 毒性效应差异分析
不同品种小白菜的Cd 毒性效应存在较大差异,其中,SZQ、YDE、HYG 为Cd 耐性品种,NY、KR、SG259 为Cd 敏感性品种.耐性品种YDE、HYG 和SZQ 在低浓度Cd (0.1 mg/L)时出现RRE 峰值,存在一定的Cd 兴奋效应,其余品种均无Cd 兴奋效应,且随着Cd 浓度增加,全部小白菜均表现出不同程度的毒性效应.Liu 等[37]发现,在Cd 浓度为0.658 mg/kg时对部分小白菜品种生物量有兴奋效应,随着浓度增加,不同小白菜品种的毒性效应具有显著差异.王文光等[38]研究发现,在低Cd 浓度(≤1 mg/L)下可促进番茄种子萌发,而随着浓度增加,发芽率、发芽势和根伸长减少,且不同品种番茄种子对Cd 溶液的敏感性存在一定差异,表达出不同的耐受能力.可见,由于受体或毒性表征终点不同可导致Cd 兴奋效应浓度和毒性浓度范围存在差异.
不同品种小白菜Cd 敏感性的差异较大,该研究发现小白菜品种间EC20Cd最大值是最小值的12 倍以上,最大、最小 EC50Cd相差超过5 倍,效应比例(抑制水平)越低,品种间的比例效应浓度差距越大.孙聪等[29]选取17 个水稻品种研究发现,不同品种Cd 的EC50Cd最大和最小值分别为61.61 和4.3 mg/kg,二者相差约14 倍.张乙涵等[39]探究四环素对8 种不同植物的敏感性差异,发现不同植物之间的EC10Cd相差约16 倍,EC50Cd相差12 倍.因此,选取不同受体或毒性效应比例得到的毒性数据差异性较大,在实际应用中可根据污染物不同毒害程度获取特定受体的毒性阈值,以保护多数的小白菜品种不受Cd 毒害.
Arshad 等[40]发现,在Cd 胁迫下小麦的SOD、POD活性升高,CAT、AsA 和酚类物质活性降低,导致产量和品质降低.钱雷晓等[41]认为,Cd 可导致小白菜对氮素的吸收代谢紊乱,光合速率降低,且其胁迫响应的程度与小白菜的品种有关.Ronzan 等[6]研究表明,Cd 胁迫水稻时,其根系发育受到抑制,改变了侧根原基的组织和发育,对根系结构产生了负面影响,造成生物合成和运输的紊乱.
3.2 Se 对小白菜Cd 毒性效应的影响
Se 对不同品种小白菜的Cd 毒性效应影响较大.Se 对小白菜的Cd 剂量-效应影响具有双重作用,在较低Cd 浓度时,Se 有效减缓了Cd 对小白菜的毒性效应,主要表现为低Cd 浓度条件下,Se-Cd 处理组Cd 兴奋效应及峰值RRE 对应的Cd 浓度均显著大于Cd 处理组;较高Cd 浓度时,Se 增强了Cd 对小白菜的毒性效应,显著抑制小白菜的生长发育.Se 对作物Cd 毒性影响的双重性已有类似报道,余垚等[42]发现,添加合适浓度Se 后,部分受Cd 胁迫的青菜地上和地下生物量有上升的趋势;Malik 等[43]研究发现,Se 可以促进植物螯合肽(PC)的形成与提高PC 合成酶的活性,因为PC 含有大量巯基,巯基可络合Cd 离子,将Cd 转化为毒性较小的络合形态,减少对植物的毒害作用.Ding 等[18-19]发现高Cd 浓度条件下,添加Se 可显著降低细根的比例和破坏细胞膜,导致根系细胞对Cd 渗透增强并加剧Cd 毒性作用.
目前关于Se-Cd 复合作用下系统性的Cd 毒性研究较少.该研究在人工气候培养箱中采用水培方式开展相关研究,但与土壤环境中的Cd 及Se-Cd 对小白菜的生态效应仍存在一定的差异,有必要进一步采用土培方式开展研究,以提高农田土壤Cd 生态风险控制的实践意义.
3.3 SSD 曲线的应用
SSD 是一种置信度较高的统计学外推方法,当给出特定效应时,可预测其发生的概率[44].Wheeler等[31]建议样本量至少需要10~15 种,以使得到的HC5更可靠[44].该试验选取了11 种小白菜进行研究,样本量满足SSD 法的样本量要求.
EPA 风险评价推荐使用Log-normal 分布模型,新西兰和澳大利亚则采用Burr-Ⅲ分布模型[31]进行SSD 曲线拟合并确定HC5,王小庆等[32]利用Gamma、Log-normal、Log-logistic、Burr-Ⅲ和Weibull 5 种分布模型来拟合不同类型土壤铜SSD 曲线,发现Burr-Ⅲ分布模型最优.该研究选取Burr-Ⅲ分布模型,基于SSD 探究了Cd 及Se-Cd 对小白菜的RRE 毒性效应,适宜浓度Se 有效提高了小白菜Cd 毒性阈值HC520和HC550,这与Semenzin 等[26]的研究结果一致.基于EC20Cd和EC50Cd得到的HC520和HC550有较大差异,且保护的品种可能发生变化,张乙涵等[39]也发现基于EC10和EC50得到的四环素耐性植物分别为莴苣和玉米,而敏感性品种相同.因此,研究物种敏感性分布和毒性阈值时必须考虑到效应比例及其他污染物共同作用的影响.
4 结论
a) 小白菜品种YDE (油冬儿)、HYG (黑油冠)、SZQ (苏州青)为Cd 耐性品种,NY (奶油)、SG259 (苏冠259)、KR (抗热)为Cd 敏感性品种.低Cd (镉)浓度(≤1 mg/L)对耐性品种具有兴奋效应,且在0.1 mg/L浓度时其相对根伸长RRE 最大,其余品种RRE 均随Cd 浓度的增加而降低.
b) 适宜浓度Se (硒)对小白菜Cd 毒性影响具有双重效应,Se 可缓解低浓度Cd 对小白菜尤其是敏感性品种的毒性,增强了高浓度Cd 对多数小白菜品种的毒性,Se 对不同品种小白菜产生双重效应的Cd 浓度范围存在较大差异.
c) 适宜浓度的Se 可影响小白菜的Cd 物种敏感性分布,并显著提高了保护95%小白菜品种不受到Cd 胁迫的毒性阈值,Se-Cd 作用下的H C520(12.00 mg/L)和 HC550(16.28 mg/L)分别为Cd 作用下的4.0 和1.4倍,研究成果为小白菜种植及富Se 环境下的Cd 生态风险控制提供参考.
