玉米LEC1基因家族的鉴定与生物信息学分析
2022-02-25刘畅畅黄敏
刘畅畅 黄敏

摘要:LEC基因家族包含LEC1和LEC2,参与植物的生长发育及储藏物质的积累。LEC1(Leafy Cotyledon 1)是NF-YB9蛋白家族的成员,编码了CCAAT-box结合转录因子HAP3(Heme activated protein 3)亚单位。利用生物信息学方法,获得了16个玉米LEC1基因。分析发现,ZmLEC1蛋白大部分都是酸性的不稳定亲水蛋白,含有1个保守结构域CBFD_NFYB_HMF及10个保守基序。通过与拟南芥LEC1基因和水稻LEC1基因的聚类分析,将玉米LEC1基因家族分为4类:ClassⅠ、ClassⅡ、ClassⅢ、ClassⅣ。经RNA-seq等分析发现,玉米LEC1家族基因具有组织特异性,响应非生物胁迫。启动子分析发现,玉米LEC1基因启动子上含有响应激素和非生物胁迫的顺式作用元件。
关键词:玉米;LEC1基因家族;生物信息学
中图分类号:S513 文献标志码:A 文章编号:1001-1463(2022)01-0031-07
doi:10.3969/j.issn.1001-1463.2022.01.007
Identification and Bioinformatics Analysis of LEC1 Gene Family in Maize
LIU Changchang 1, HUANG Min 1, 2
(College of Life Science, Yangtze University, Jingzhou Hubei 434025, China)
Abstract:The LECfamily genes, consisting of LEC1 and LEC2, are involved in the development and accumulation of storage materials in plants. LEC1(Leafy Cotyledon 1) is a member of the NF-YB9 protein family, encoding the CCAAT-box binding transcription factor HAP3 (Heme activated protein 3) subunit. In this study, we identified 16 LEC1 genes by using bioinformatics. Most of the ZmLEC1 proteins are acidic unstable hydrophilic proteins, containing a conserved domain CBFD_NFYB_HMF and 10 conserved motifs. By aligning the sequence of LEC1 genein Arabidopsis and rice, respectively, these genes were divided into four categories as ClassⅠ、ClassⅡ、ClassⅢ、ClassⅣ. According to the analysis of RNA-seq, ZmLEC1 preferentially expressed in specific tissues, responding to the abiotic stress. Promoter region analysis revealed that the promoter of ZmLEC1 contained cis-acting elements in response to hormonal and abiotic stresses.
Key words:Maize(Zea mays L.);LEC1 gene family;Bioinformatics
植物的胚胎發育一直是人们的研究热点,随着科技水平的迅猛发展,有大量胚胎发育相关基因被克隆出来。LEC1基因最早是在拟南芥中发现的,其在早期发育阶段发挥作用,以维持胚胎细胞的命运[1 ]。LEC1的异位胚后表达能够诱导拟南芥叶片组织中胚状结构的形成、幼苗发育畸形、子叶生长异常,根系生长也受到了抑制[2 ]。继拟南芥LEC1基因被克隆后,玉米、胡萝卜、花生、棉花等作物的LEC1基因也相继被克隆出来并进行了初步的探索与研究[3 - 6 ]。研究发现,基因LEC1还参与植物脂肪酸生物合成的缩合、伸长和去饱和反应。LEC1基因的过度表达导致拟南芥脂肪酸生物合成基因的整体表达增加[7 ],其在玉米中过量表达时提高了种子的油脂含量[8 ]。基因LEC1在油菜中的过量表达可以是其种子油脂含量增加7%~16%[9 ]。
2020年中国玉米产量高达2.61亿t,作为我国的高产粮食作物,玉米是畜牧业、养殖业等重要的饲料来源,也是许多工业用品不可缺少的原料之一,在全国31个省、市、自治区均有种植[10 ]。胚胎的发育影响了种子萌发速率和整齐度,同时也会影响后期的抗虫害能力及产量[11 ]。目前,关于玉米LEC1基因组鉴定及生物信息学分析尚未见报道,本文利用生物信息学手段对玉米LEC1基因家族理化性质、基因结构、蛋白结构、系统发育、启动子元件,基因的差异性表达等进行了分析,以期为进一步探究玉米LEC1基因功能与调节机制奠定基础。
1 材料与方法
1.1 数据来源
通过MaizeGDB(https: //maizegdb.org/gene_center/gene)和NCBI获取玉米LEC1基因家族数据,在Ensembl Plants(http://plants.ensembl.org/index.html)上下载玉米蛋白质的全部序列,在TAIR(https://www.arabidopsis.org)网站上获取拟南芥LEC1基因的蛋白序列。
1.2 方法
1.2.1 玉米LEC1基因家族的鉴定及理化性质分析
通过TBtools工具将获得的玉米LEC1基因家族数据进行进一步筛选。将所得玉米LEC1基因家族蛋白序列提交在线网站ProtParam(https://web.expasy.org/protparam/)上,分析各玉米LEC1蛋白的氨基酸数量,分子量,等电点,蛋白疏水性,不稳定系数等理化性质。
1.2.2 玉米LEC1基因家族的染色体定位 在NCBI数据库中获取玉米LEC1基因家族各成员的在染色体上的位置信息及玉米各染色体长度信息,将上述信息整理并上传至MapGene2Chrom(http://mg2c.iask.in/mg2c_v2.1/),在线网站绘制出玉米LEC1基因染色体定位图。
1.2.3 玉米LEC1基因结构及蛋白结构域分析 通过TBtools工具提取玉米LEC1基因家族各成员的外显子、内含子结构信息,并上传至GSDS2.0(http://gsds.cbi.p-ku.edu.cn/)。通过MEME(http://alternate.meme-suite.org/tools/meme)在線网站分析玉米LEC1-1~16蛋白保守基序。利用NCBI中Batch CD-Search工具分析ZmLEC1-1~16蛋白结构域,并通过TBtools处理得到可视化图形。
1.2.4 玉米LEC1基因启动子顺式元件分析 玉米LEC1-1~16基因上游2 000 bp的启动子序列,通过PlantCARE(http://bioinformatics.psb.ugent.be/webt ools/plantcare/html/)分析顺式作用元件,并利用Tbtools对顺式作用元件的在基因上的位置进行可视化处理。
1.2.5 玉米LEC1蛋白的系统发育分析 从Ensembl Plants数据库下载水稻的全部蛋白质信息,利用从PFAM网站(http://pfam.xfam.org/)得到的蛋白保守结构域CBFD_NFYB_HM的Pfam号(PF 00808)进行信息搜索。将得到全部9个水稻LEC1、14个拟南芥LEC1的蛋白序列和玉米的16个LEC1蛋白序列导入到Tbtools中进行系统发育树构建。
1.2.6 玉米LEC1基因表达模式分析 从MaizeGDB网站下载玉米根、叶、种子、胚乳的转录数据及玉米在冷、热、盐、紫外胁迫和正常生长下的转录组数据,将玉米LEC1-1~16的数据筛选出来并上传至TBtools进行热图绘制。
2 结果与分析
2.1 玉米LEC1基因家族的鉴定及理化性质分析
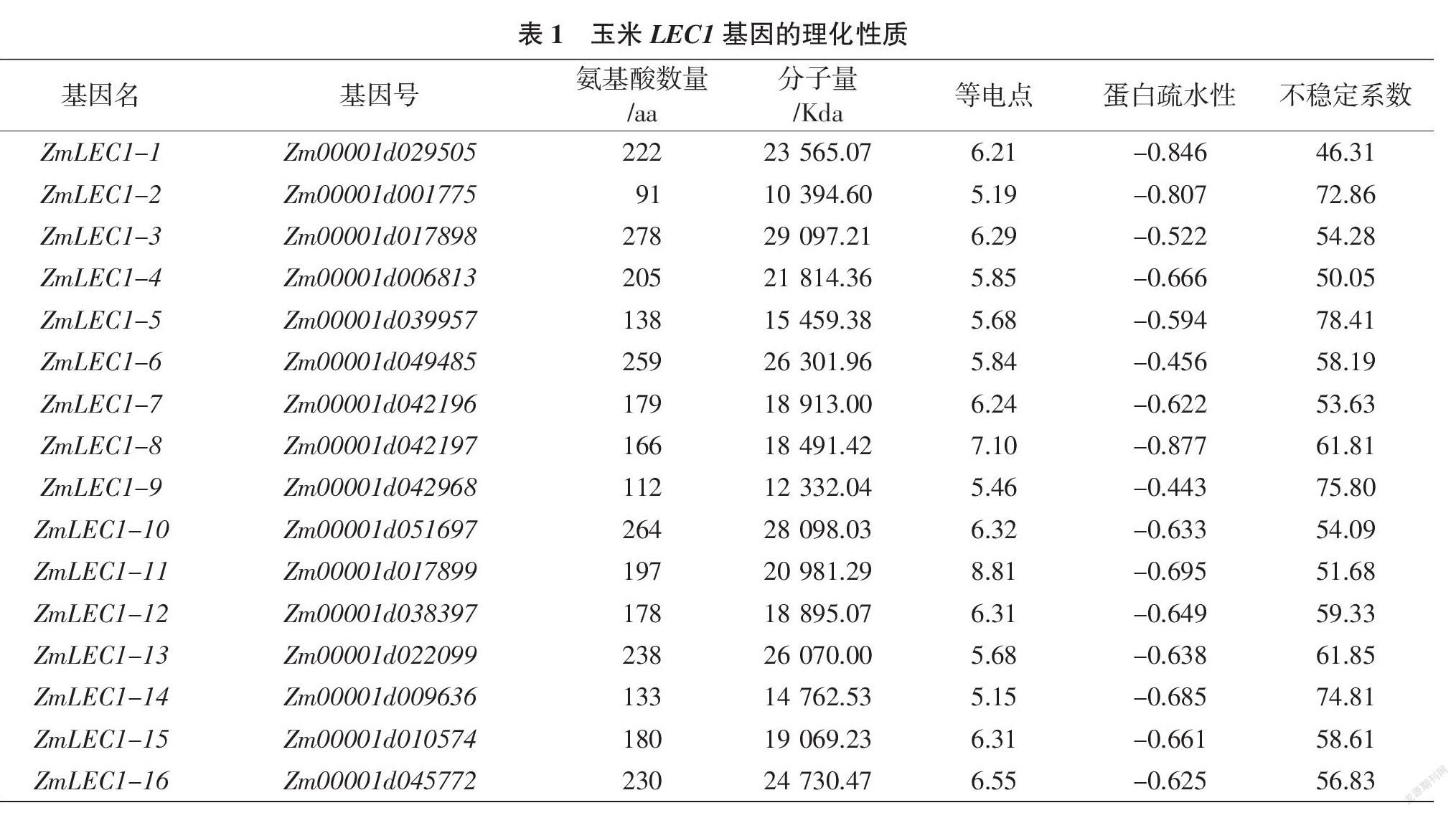
通过从TAIR上得到的拟南芥LEC1蛋白的氨基酸序列,在NCBI和MAIZEGDB上进行BLAST搜索,最终筛选得到16个玉米LEC1基因。根据它们在染色体上的排列顺序,将其命名为ZmLEC1-1~16。玉米LEC1家族蛋白的氨基酸数量为91~278 aa,分子量为10 394.60~29 097.21 Kda。蛋白的等电点除ZmLEC1-9(7.10)和ZmLEC1-11(8.81)外,其余均在7.00以下,说明玉米LEC1蛋白大多是呈酸性的。且蛋白疏水性均小于0,不稳定系数均大于40,说明玉米LEC1蛋白是不稳定的亲水蛋白(表1)。
2.2 玉米LEC1基因家族染色体定位
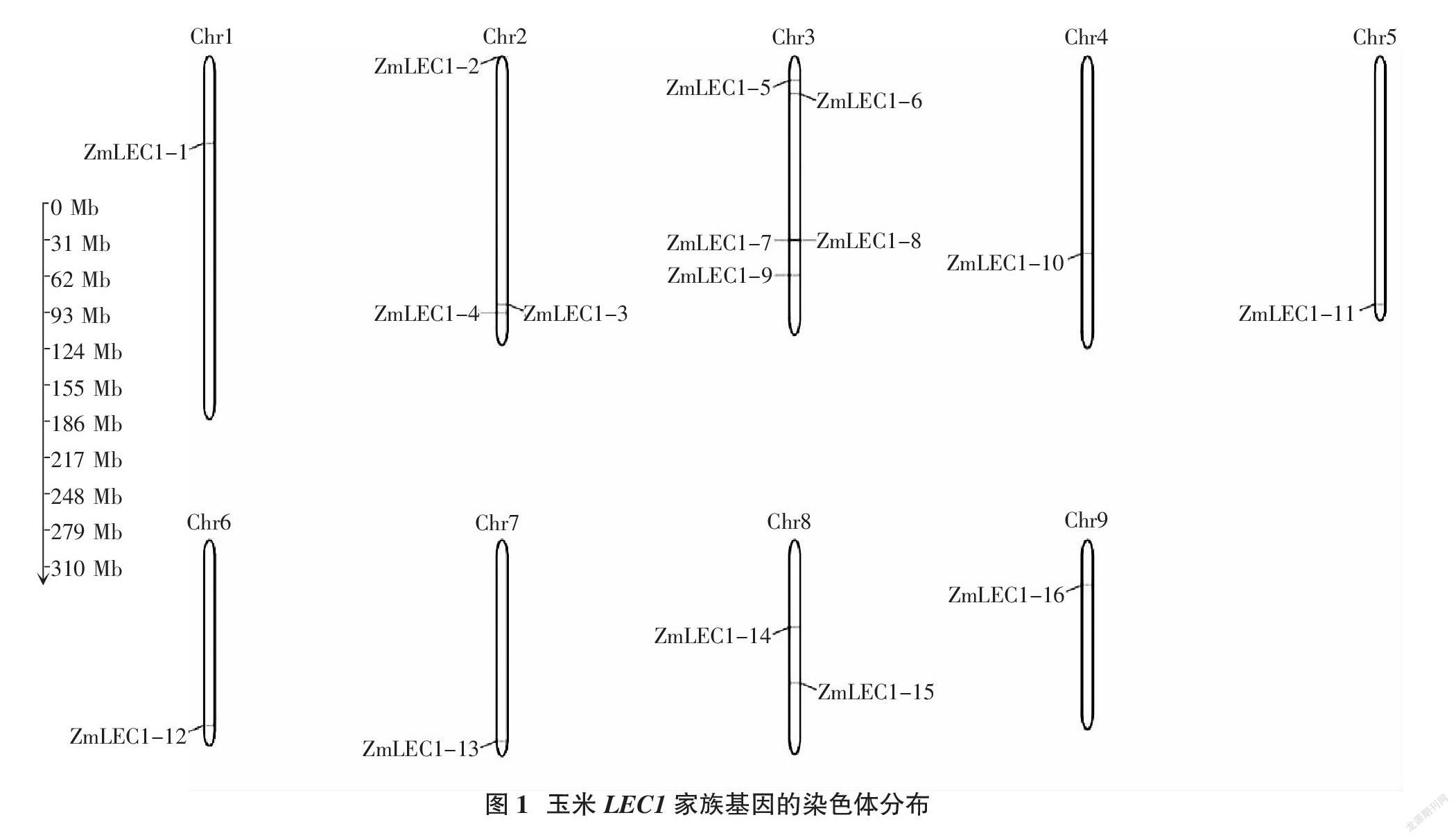
为了进一步了解ZmLEC1基因家族在染色体上的具体分布,通过MG2C在线网站对ZmLEC1-1~16的注释信息进行分析,结果显示,除了第10条染色体无基因分布外,其余染色体上均有分布,其中第2条染色体上有3个基因,第3条染色体上有5个基因,第8条染色体上有2个基因,其余每条染色体上均有1个基因(图1)。
2.3 玉米LEC1基因结构及蛋白结构域
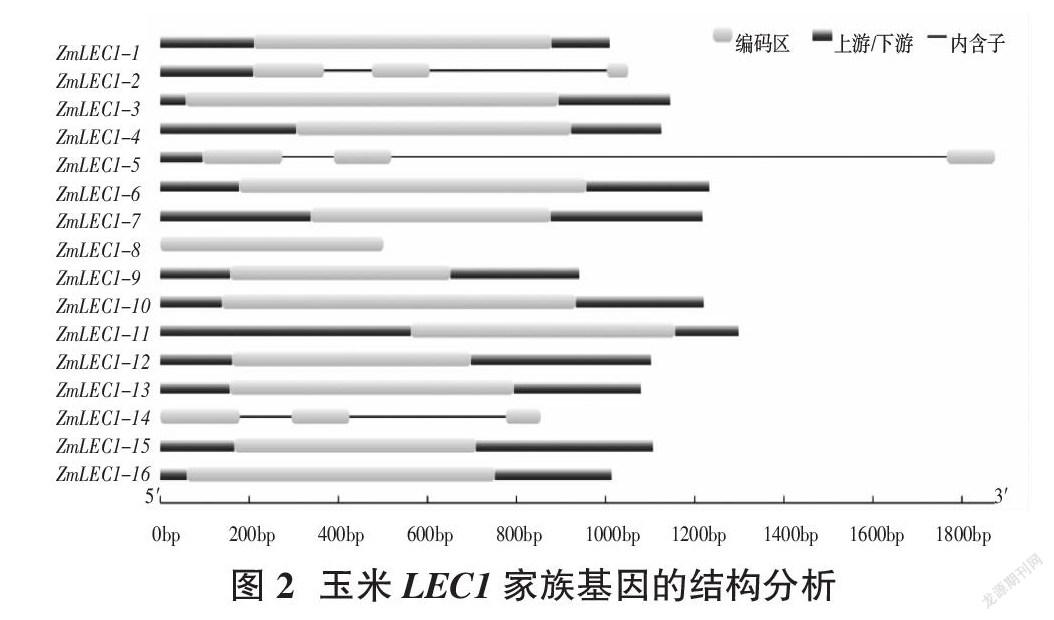
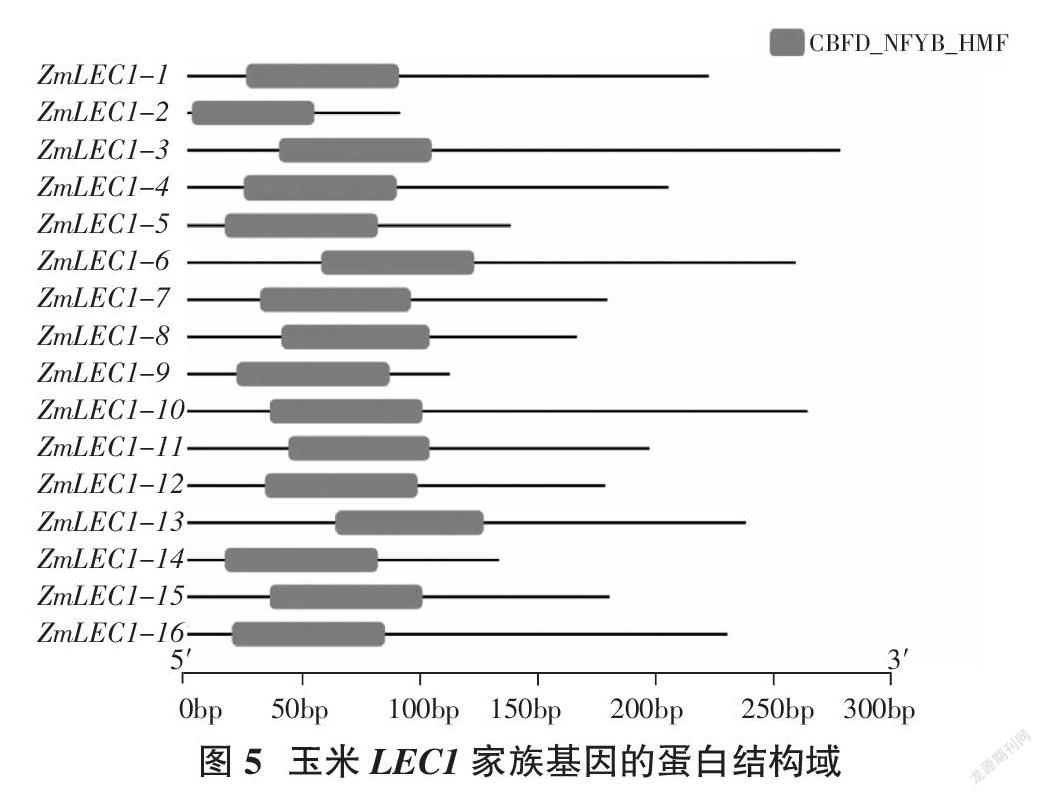
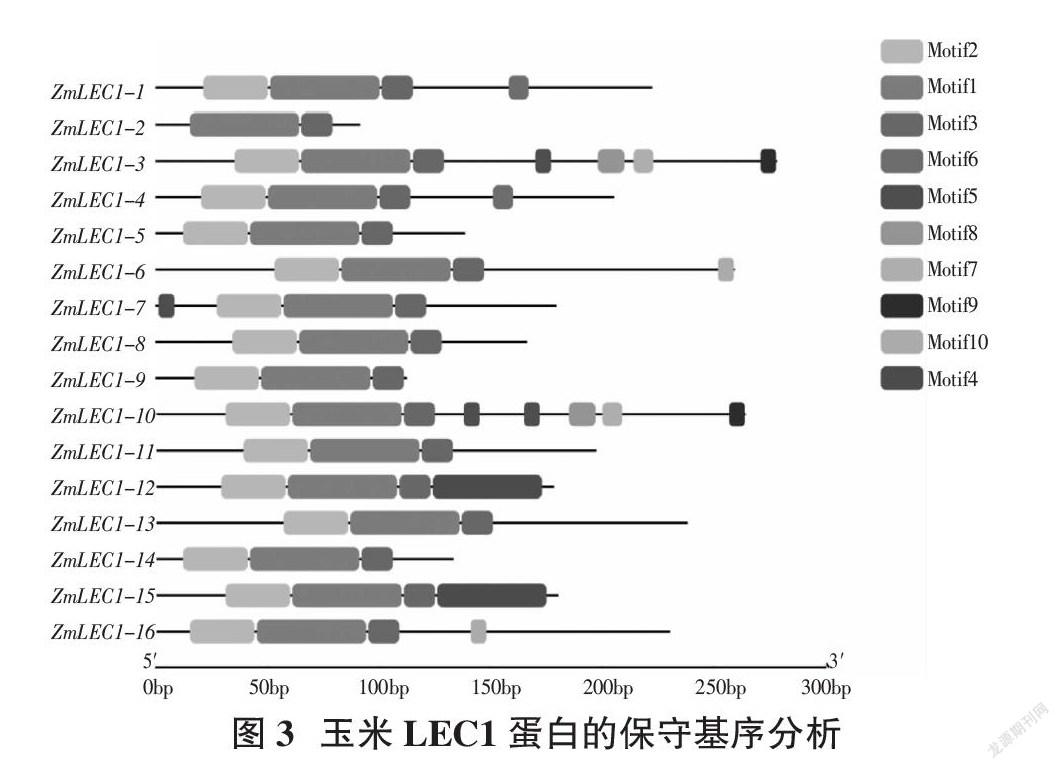
通过GSDS2.0在线网站分析得到玉米LEC1基因结构,发现其编码区为1~3个,其中ZmLEC1-8、ZMLEC1-14没有5′UTR和3′UTR,ZmLEC1-2,ZMLEC1-5没有3′UTR(图2)。利用MEME在线工具对LEC1基因家族的保守结构域进行分析,得到10个保守的motif(图3),命名为motif1-motif10,但每个ZmLEC1-1~16蛋白的motif排列顺序并不完全相同,其中ZmLEC1-3包含7个motif,ZmLEC1-1、ZmLEC1-4、ZmLEC1-6、ZmLEC1-7、ZmLEC1-12、ZmLEC1-15、ZmLEC1-16包含4个motif,ZmLEC1-5、ZmLEC1-8、ZmLEC1-9、ZmLEC1-11、ZmLEC1-13、ZmLEC1-14包含3个motif,ZmLEC1-2包含2个motif(图4)。通过TBtools对LEC蛋白结构域进行分析,发现基因ZmLEC1-1~16蛋白拥有保守的蛋白结构域CBFD_NFYB_HMF(图5)。
2.4 玉米LEC1家族基因啟动子特异顺式作用元件
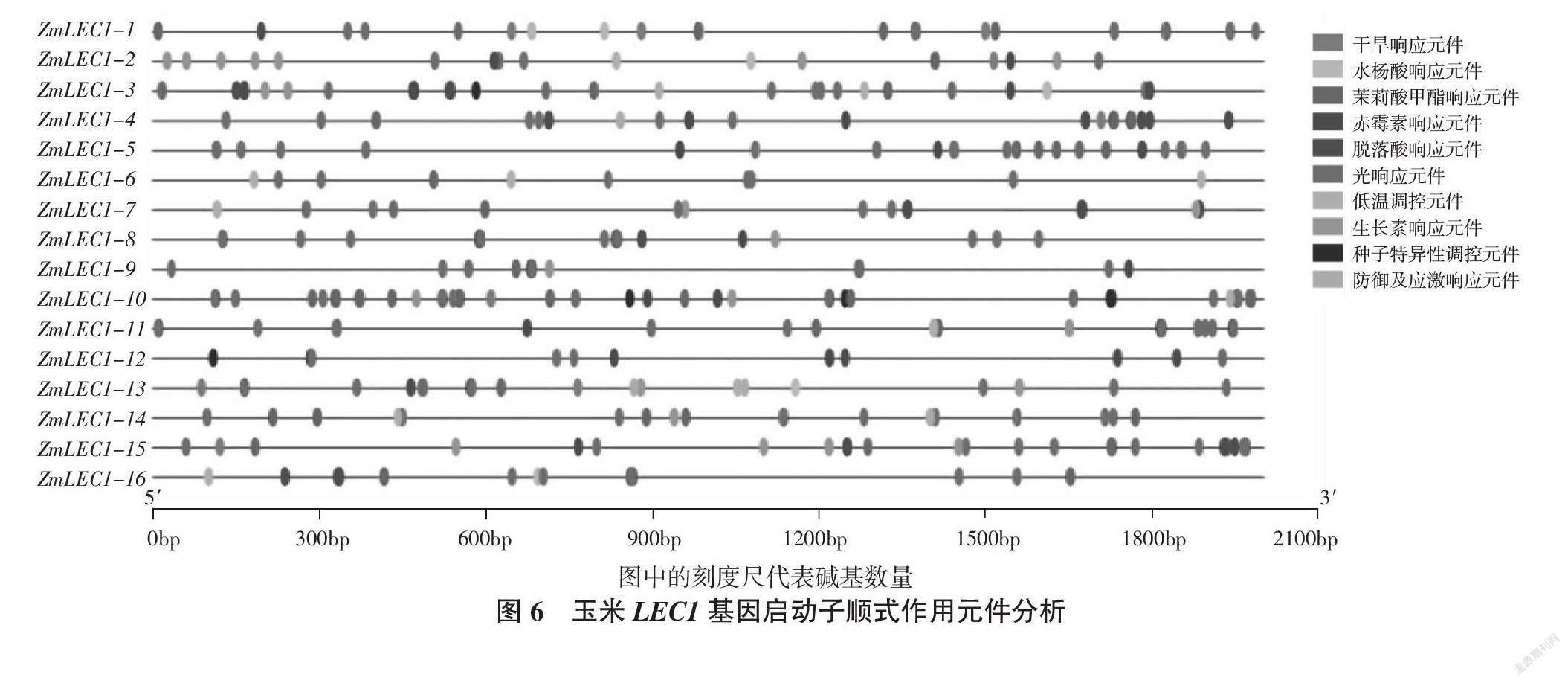
选取玉米ZmLEC1-1~16码子上游2 000 bp的序列进行启动子顺式作用元件的分析。Plant CARE在线分析结果表明(图6),玉米ZmLEC1-1~16区域含有茉莉酸甲酯响应元件(TGACG-motif,CGTCA-motif)、生长素反应元件(TGA-element,Aux RR-core)、水杨酸反应元件(TCA-element)、赤霉素反应元件(GARE-motif,TATC-box,P-box)、脱落酸反应元件(ABRE,SARE)、光响应元件(G- box,Sp1,GATA-motif,Box 4,AE-box,I-box)、干旱响应元件(MBS)、低温调控元件(LTR)、防御及应激响应元件(TC-rich repeats)、种子特异性调控元件(RY-element)。
2.5 玉米LEC1基因的系统发育分析
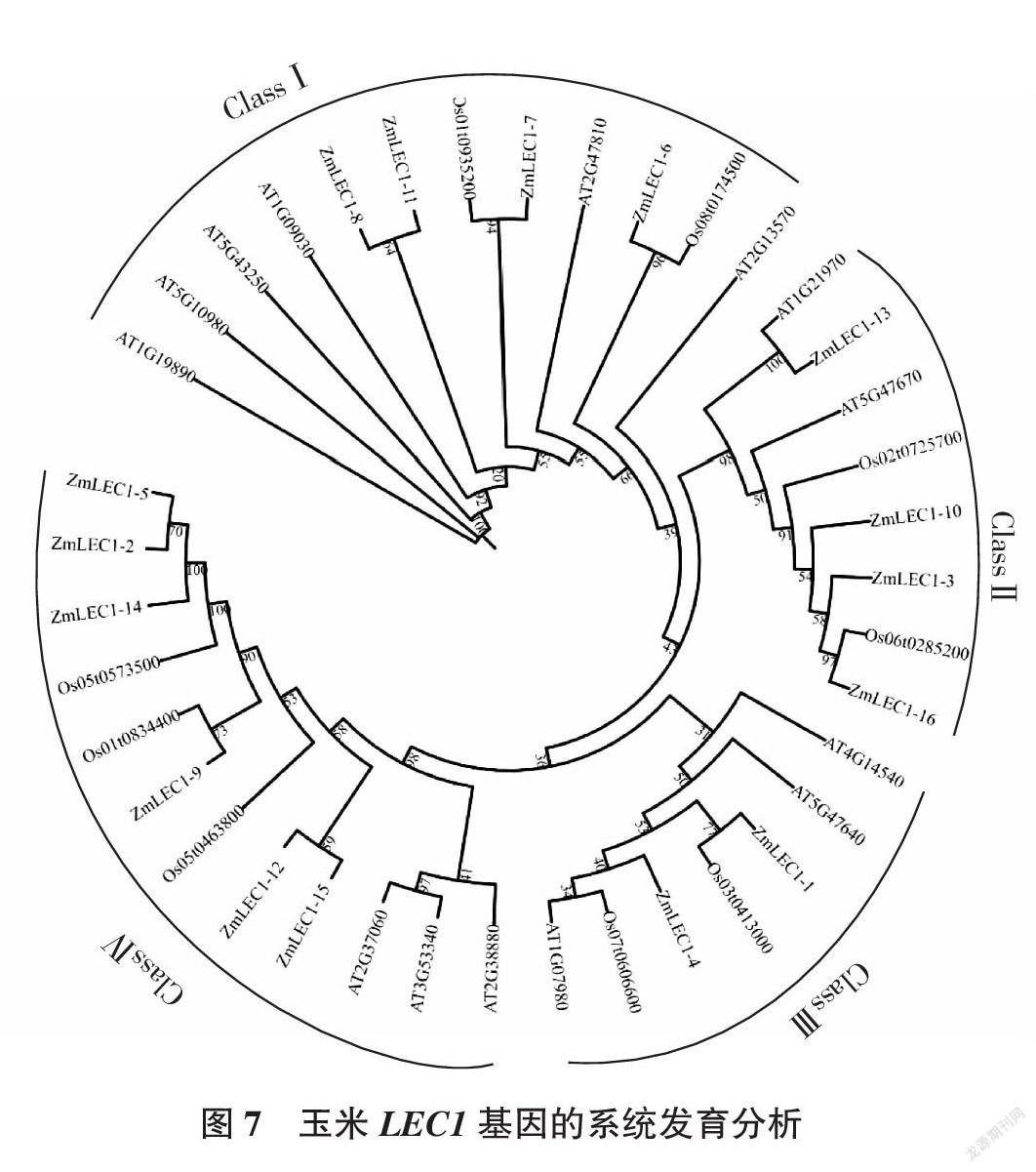
在Ensembl Plants数据库搜索得到9个水稻LEC1和14个拟南芥LEC1的蛋白序列,将获得的拟南芥、水稻和玉米的蛋白序列导入到TBtools中进行系统发育树的构建。结果显示(图7),39个LEC1基因可分为4个亚家族(ClassⅠ、ClassⅡ、ClassⅢ、ClassⅣ),其中ClassⅠ中包含了4个玉米基因(ZmLEC1-6,ZmLEC1-7,ZmLEC1-8,ZmLEC1-11)、2个水稻基因(Os01t0935200,Os08t017450),6个拟南芥基因(AT1G19890,AT5G10980,AT5G 43250,AT1G09030,AT2G47810,AT2G13570);ClassⅡ包含了4个玉米基因(ZmLEC1-3,ZmLEC 1-10,ZmLEC1-13,ZmLEC1-16)、2个水稻基因(Os02t0725700,Os06t0285200)、2个拟南芥基因(AT1G21970,AT5G47670);ClassⅢ中包含了2个玉米基因(ZmLEC1-1,ZmLEC1-4)、2个水稻基因(Os03t0413000,Os07t0606600)、3个拟南芥基因(AT4G14540,AT5G47640,AT1G07980);ClassⅣ包含了6个玉米基因(ZmLEC1-2,ZmLEC1-5,ZmLEC1-9,ZmLEC1-12,ZmLEC1-14,ZmLEC1- 15)、3个水稻基因(Os05t0573500,Os01t 0834400,Os05t0463800)、3个拟南芥基因(AT2G37060,AT3G53340,AT2G38880)。
2.6 玉米LEC1基因表达模式分析
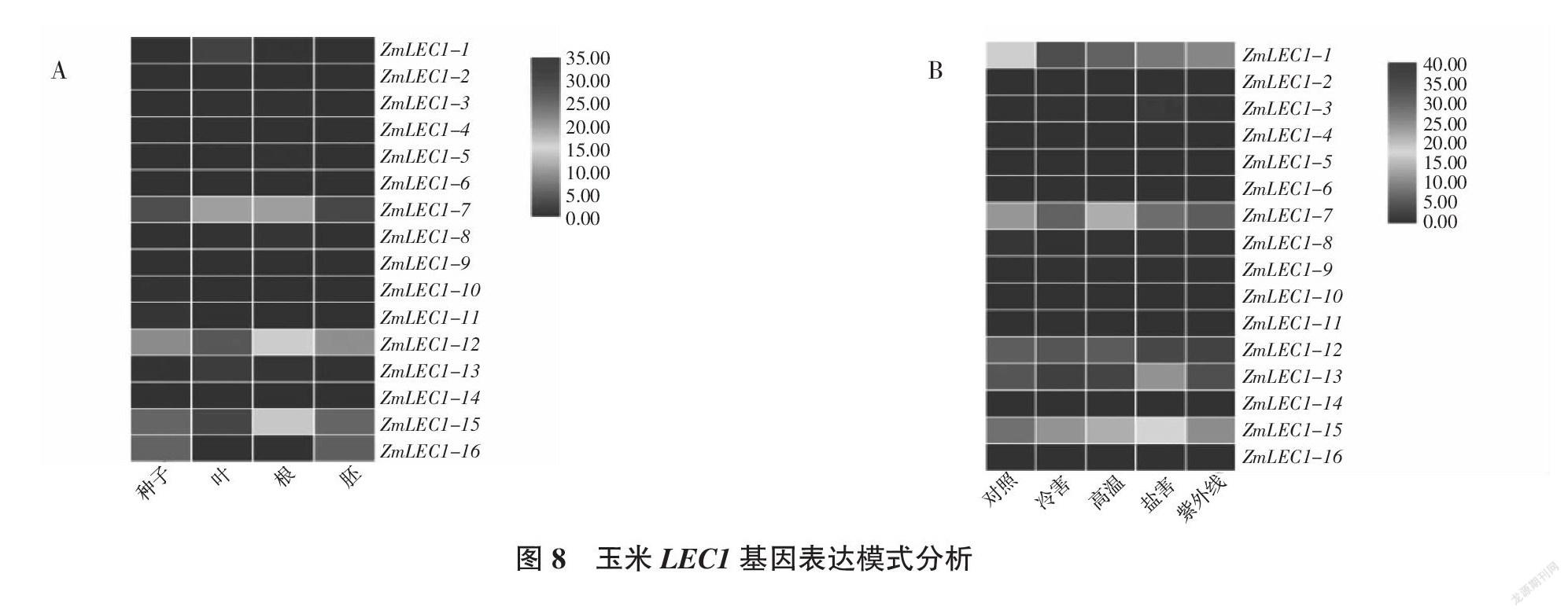
为研究玉米LEC1基因在不同组织和不同胁迫下的特异性表达情况,对玉米转录组数据库的数据进行分析。结果发现(图8),基因ZmLEC1-12在根中的表达量均高于其他组织,ZmLEC1-15在叶中表达量最高,ZmLEC1-7在种子和胚乳中表达量最高。环境对植物的生长发育有着很大的影响,其中盐害,高温,低温等都可能会导致玉米生长发育受到损伤。由分析结果可看出,玉米ZmLEC1-1、ZmLEC1-7、ZmLEC1-12、ZmLEC1-15这4个基因比较容易受到胁迫影响,其中ZmLEC1-12在盐害胁迫下转录水平增高了9倍。
3 结论与讨论
通过生物信息学分析共得到玉米ZmLEC1-1~16,其不均匀分布在9条染色体上。玉米LEC家族具有保守的蛋白结构域,除ZmLEC1~9以外,ZmLEC1蛋白均为酸性的不稳定亲水蛋白,含有1个保守结构域CBFD_NFYB_HMF及10个保守基序。通过与拟南芥LEC1基因和水稻LEC1基因的聚类分析,将玉米LEC1基因家族分为4类:ClassⅠ、ClassⅡ、ClassⅢ、ClassⅣ。RNA-seq等分析发现,玉米LEC1家族基因具有组织特异性,响应非生物胁迫。启动分析发现,玉米LEC1基因启动因子上含有响应激素和非生物胁迫的顺式作用元件。玉米LEC1基因在种子、叶片、根、胚乳中均有不同的表达水平,且基因ZmLEC1~12在盐害胁迫下表达量有明显的提高。本研究为玉米基因LEC1的功能研究提供依据,也为ZmLEC1基因的后续研究奠定基础。
LEC1基因参与调控非生物胁迫。刘豪等[12 ]通过荧光定量PCR对小麦LEC1基因在ABA、干旱、高温胁迫处理后的表达量进行分析,发现ABA处理后的小麦TaLEC1基因表达量上升,干旱和高温胁迫下小麦TaLEC1基因在胁迫初期急剧上调表达,原因很有可能是在小麦受到高温和干旱影响时,ABA发挥作用使小麦脱水率降低。同样通过荧光定量PCR实验发现花生LEC1基因对干旱、盐害、低温胁迫有明显的响应[13 ]。通过生物信息学分析启动子作用元件发现,LEC1基因还可能参与茉莉酸甲酯、生长素、水杨酸、赤霉素等依赖的胁迫响应。人们通过不同耕作方式和研究响应机理来丰富玉米品种资源[14 ]。
生长环境对作物的生长发育有很大的影响,盐害是影响我国玉米产量的主要因素之一。盐害是指土壤含盐过多,土壤溶液渗透压过高引起植物生长发育不良的现象。玉米对盐的承受量最高在0.0017 mol/L左右,土壤盐害高度影响玉米的生长、发育及产量[15 ]。玉米遭受到盐害影响时会使胞内渗透压失衡,导致细胞质壁分离[16 ]、离子失衡、植株失水[17 ]、细胞器的功能丧失[18 ],导致玉米植株死亡。本研究发现,基因ZmLEC1-7在受到盐害胁迫时高度表达,表明ZmLEC1-7基因可能在盐害胁迫方面起着重要作用。
参考文献:
[1] LOTAN T,OHTO M A,YEE K M,et al. Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells[J]. Cell,1998,93(7):1195-1205.
[2] WEST,MAL. LEAFY COTYLEDON1 is an essential regulator of late embryogenesis and cotyledon Identity in Arabidopsis[J]. The Plant Cell,1994,6(12):1731-1745.
[3] ZHANG S,WONG L,LING M,et al. Similarity of expression patterns of knotted1 and ZmLEC1 during somatic and zygotic embryogenesis in maize(Zea mays L.)[J]. Planta,2002,215(2):191-194.
[4] YAZAWA K, TAKAHATA K, KAMADA H. Isolation of the gene encoding Carrot leafy cotyledon1 and expression analysis during somatic and zygotic embryogenesis[J]. Plant Physiology and Biochemistry,2004,42(3):215-223.
[5] 李愛芹,夏 晗,王兴军,等. 花生LEC1基因的克隆及表达研究[J]. 西北植物学报,2009,29(9):1730-1735.
[6] 樊可心,曹爱萍,张文婷,等. 棉花G. LEC1A基因的克隆和鉴定[J]. 石河子大学学报(自然科学版),2015,33(5):535-540.
[7] MU J,TAN H,ZHENG Q,et al. LEAFY COTYLEDON1 is a key regulator of fatty acid biosynthesis in Arabidopsis[J]. Plant physiology,2008,148(2):1042-1054.
[8] BO SHEN,WILLIAM B ALLEN,PEIZHONG ZHENG,et al. Expression of ZmLEC1 and ZmWRI1 Increases Seed Oil Production in Maize[J]. Plant Physiology, 2010,153(3):980-987.
[9] ELAHI N,DUNCAN R W,STASOLLA C. Modification of oil and glucosinolate content in canola seeds with altered expression of Brassica napus LEAFY COTYLEDON1[J]. Plant Physiology and Biochemistry,2016,
100:52-63.
[10] 南彩霞,张南冰,邓喆元. 玉米新品种强硕168选育报告[J]. 甘肃农业科技,2019(4):1-4.
[11] 连彩云,马忠明. 玉米种子活力研究综述[J]. 甘肃农业科技,2019(5):64-68.
[12] 刘 豪,王艳丽,孟晓丹,等. 小麦TaLEC1基因的克隆及其表达特性分析[J]. 西北植物学报,2019,
39(5):904-910.
[13] 席旭东,常 宏,车 卓,等. 6个玉米新品种在定西旱作农业区的引种初报[J]. 甘肃农业科技,2019(7):61-65.
[14] 潘丽娟,王 冕,苏茂文,等. 花生转录因子LEC1的表达特性分析[J]. 花生学报,2021,50(2):15-20.
[15] 李 明,冷冰莹,张晗菡,等. 盐胁迫下调控玉米胞内Na+/K+比稳定的主要机制与措施[J]. 山东农业科学,2021,53(6):133-138.
[16] RANA,MUNNS,MARK,et al. Mechanisms of salinity tolerance[J]. Annual Review of Plant Biology,2008,
59:651-681.
[17] ZHAO Y,LIU M,HE L,et al. A cytosolic NAD+-dependent GPDH from maize (ZmGPDH1) is involved in conferring salt and osmotic stress tolerance[J]. BMC Plant Biology,2019,19(1):16. https://doi.org/10.118 6/s12870-018-1597-6.
[18] 武 燕,解丹丹,马小亭,等. 外源Ca2+对盐胁迫下玉米幼苗生理指标的影响[J]. 安徽农业科学,2014,
42(12):3471-2472.
