云顶山亚高山草甸土壤可培养细菌数量特征
2022-02-24张宝贵朱茜刘晓娇吴佳灵常丽娟
张宝贵 朱茜 刘晓娇 吴佳灵 常丽娟
摘 要:作为土壤生物的主要组成部分,土壤微生物在整个陆地生态系统物质循环和能量转换过程中发挥着非常重要的作用,且对环境变化极其敏感。以吕梁山中麓云顶山亚高山草甸为研究区域,以不同海拔(2 450 m、2 500 m和2 550 m)、不同深度(0~10 cm和20~30 cm)土壤为研究对象,采用可培养方法測定了土壤细菌的数量,同时测定了土壤理化性质并进行了相关分析。结果表明,研究区不同海拔、不同深度可培养细菌数量介于3.13×109 ~10.4×109 CFU/g,且同一海拔表层土壤细菌数量均显著高于30 cm深处(P<0.05),不同海拔0~10 cm与20~30 cm深度土壤细菌数量均随着海拔高度的升高而显著下降(P<0.05)。细菌数量与土壤全碳(STC)和土壤温度(ST)呈极显著正相关(P<0.01),与土壤酸碱度(pH值)呈显著正相关(P<0.05),表明土壤全碳、土壤温度及土壤pH值是影响该区域土壤细菌数量的主要环境因子。研究结果可为区域应对气候变暖提供理论依据与数据支撑。
关键词:云顶山;亚高山草甸;可培养细菌;数量特征
文章编号:1005-2690(2022)01-0017-05 中国图书分类号:S812.2 文献标志码:B
土壤微生物是地球上多样性最高、物种最丰富的生物类群之一,参与了土壤中几乎所有的物质转化过程,对森林、草原、湿地及农田等生态系统具有重要影响,是驱动地球生物化学过程的关键因素[1]。作为土壤圈层能量交换的中介,土壤微生物可以直接分解土壤中的有机质,促进腐殖质形成[2],提供植被生长所需的营养[3],还能间接反映土壤的肥力状况[4]。虽然土壤微生物生物量只占土壤有机质的3%左右,却是植物养分转化和循环的驱动力,在土壤肥力和生态系统评价中起着非常重要的作用[5]。
在当前气候变化具有区域差异性的背景下,人类活动对土壤生态系统的干扰不断加强,使得我国土壤微生物群落结构更为复杂[6]。作为响应气候变暖的产物,亚高山草甸目前已经引起诸多国内外学者的重视。学者主要针对亚高山草甸植物物种的高氮保留能力[7]、物种扩张和土壤养分的影响[8]开展了相关研究,而对地下微生物研究鲜有报道。王萍(2011)[9]研究发现,亚高山草甸土壤微生物多样性在弃耕地恢复中能够得到较好恢复;宋三多(2011)[10]研究发现,在不同雪被厚度覆盖下,亚高山草甸土壤细菌种群结构无明显变化,但对细菌的分布比例影响较大;谢勇等(2017)[11]研究发现,封育后亚高山草甸土壤细菌数量和土壤微生物总数显著降低;朱雪梅(2009)[12]研究发现若尔盖亚高山草甸不同土壤层次的细菌数量有明显差异,随着剖面层次增加,细菌数量逐渐变小。研究亚高山草甸土壤微生物有助于理清高海拔地区土壤微生物对环境变化的响应特征。
云顶山位于山西省吕梁山中部,生态环境复杂,具有大面积的亚高山草甸,属于典型的中纬度、低海拔亚高山草甸[13]。作为全球气候变化的敏感区域,山西省亚高山草甸成为诸多学者关注研究的热点。近几年对山西境内土壤微生物研究多是结合典型植被类型进行宏观方面的研究[14]。相关学者主要对山西境内亚高山草甸植被群落、物种多样性等地上生物进行了时空变化格局的研究[15-17]。上官铁梁、张峰、刘明光、刘莹、李素清、张金屯、杨斌胜等则针对云顶山这一地区的特定植物群落进行了集中研究[18-22]。
以上研究多集中在环境类型和人为干扰对植被群落多样性产生的影响上,没有对该区域土壤微生物进行研究。基于此,本研究以山西省境内云顶山不同海拔、不同深度亚高山草甸土壤为研究对象,通过可培养技术研究云顶山亚高山草甸土壤细菌数量特征,同时分析细菌数量与环境因子之间的相关性,找出限制该区域细菌数量的关键环境因子。研究结果可为系统认识区域内亚高山草甸土壤微生物响应气候变化提供理论依据与数据支撑。
1 研究地区与研究方法
1.1 研究区概况
云顶山地处吕梁山中麓,位于山西省吕梁市交城县、方山县和太原市娄烦县的交界处(111°30'~111°47'E,37°50'~38°02'N),最高海拔2 720 m,年平均气温为3~4 ℃,最热月为7月,平均气温16.1 ℃,最冷月为1月,平均气温-10.6 ℃,年均降水量600~800 mm,降水集中于6—9月,相对湿度70.9%,无霜期100 d,属于温带大陆性季风气候;土壤类型从低海拔到高海拔依次为褐土、山地褐土、山地淋溶褐土、山地棕壤、亚高山草甸土;植被类型从山麓到山顶依次为1 200~1 750 m落叶阔叶林带、1 750~2 200 m针阔叶混交林带、2 200~2 600 m寒温性针叶林带、2 600~2 720 m亚高山灌丛草甸带。
1.2 样地设计与样品采集
样品采集于2019年9月,根据云顶山亚高山草甸分布的实际范围,从底部到顶部按50 m高差确定3个采样点,分别为海拔2 450 m(YDL)、2 500 m(YDM)、2 550 m(YDT)。在每个海拔高度处,选取地势平坦的区域,设置3个间隔5 m的平行样地,在每个样地的中心挖取土壤剖面。
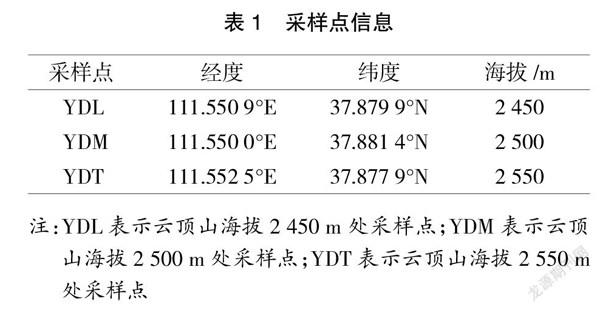
由于山地土层较薄,根据实际情况分别采集表层土(0~10 cm)和剖面底部(20~30 cm)土壤样品,盛于微生物采样袋中,随后放置于储有冰袋的低温保存箱中并运回实验室,保存于-20 ℃冰箱备用。同时采集用于土壤理化性质分析的样品。为保证采样的准确性,采样过程严格按照无菌要求进行。采样过程中使用一次性无菌PE手套,以免对土壤造成污染。采样点信息见表1。
1.3 细菌的分离培养
细菌的分离培养采用稀释涂布平板法培养,使用牛肉膏蛋白胨培养基,具体操作步骤如下。在超净工作台中称取2 g新鲜土壤放入无菌三角瓶中,加入高温灭菌后的18 mL无菌生理盐水(0.85%),以室温250 rmp摇振30 min,静置数分钟后取土壤悬液上清液200 μL加入800 μL无菌生理盐水中并混合均匀,从此溶液中吸取200 μL土壤悬液加入到800 μL无菌生理盐水中。
重复以上步骤将溶液稀释至10-7数量级。为了确保微生物培养计数的准确性,可培养细菌进行预试验,用各种稀释浓度的土壤悬液进行涂布并培养,最终确定最适合细菌生长的浓度为10-6数量级。用移液枪吸取对应浓度的土壤悬液涂布到灭菌的固体培养基平板上,同一样品同一类微生物进行3个重复处理,放置于25 ℃恒温培养箱培养7 d后计数。
1.4 土壤理化性质的测定
土壤含水量(SWC)采用烘干法(105 ℃,24 h)测定;土壤pH值采用METTLER TOLEDO FE28-Standard酸度计测定;土壤电导率(EC)、土壤总溶解性固体(TDS)和土壤盐度(SSC)采用电导率仪FE38 Standard测定;土壤全碳(STC)采用C/N分析仪2100(Elementar Vario-EL,Germany)测定;土壤温度ST采用土壤温度传感器(S-
TMB-M006)测定。
1.5 数据处理与分析
采用Excel 2010处理所有数据;用SPSS 17.0进行相关统计分析,包括单因素方差分析和相关性分析;用Origin 8.0绘图。
2 结果与分析
2.1 不同海拔、不同深度土壤理化性质变化特征
云顶山亚高山草甸不同海拔、不同深度下的土壤理化性质特征,详见表2。从表2可知,STC介于19.57~47.51 g/kg之间。从横向来看,同一海拔下随着土壤深度增加,STC呈现显著下降趋势(P<0.05);从纵向来看,同一深度STC随着海拔高度的增加而呈现先下降后上升的显著趋势(P<0.05)。
EC介于48.24~81.26 μS/cm之间。
SWC介于12.55%~51.01%之间,且变化趋势与STC一致。
ST介于9.41~17.00 ℃之间,从横向来看,同一海拔下ST变化范围与STC一致;从纵向来看,同一深度下ST随着海拔的升高呈现下降趋势。
TDS介于25.16~49.58 mg/L之间,从横向来看,随着土壤深度的增加,不同海拔样品变化趋势不同,高海拔与低海拔地区呈现显著下降趋势(P<0.05),中海拔则呈现相反趋势;从纵向来看,同一深度TDS变化趋势与STC相同,但未达到显著水平。
土壤pH值和SSC变化范围较小。其中pH值介于7.39~8.79之间,呈弱碱性;SSC介于0.03~0.04 psu之间。两者的变化趋势均与STC一致,但未达到显著水平。
2.2 不同海拔、不同深度土壤可培养细菌数量变化特征
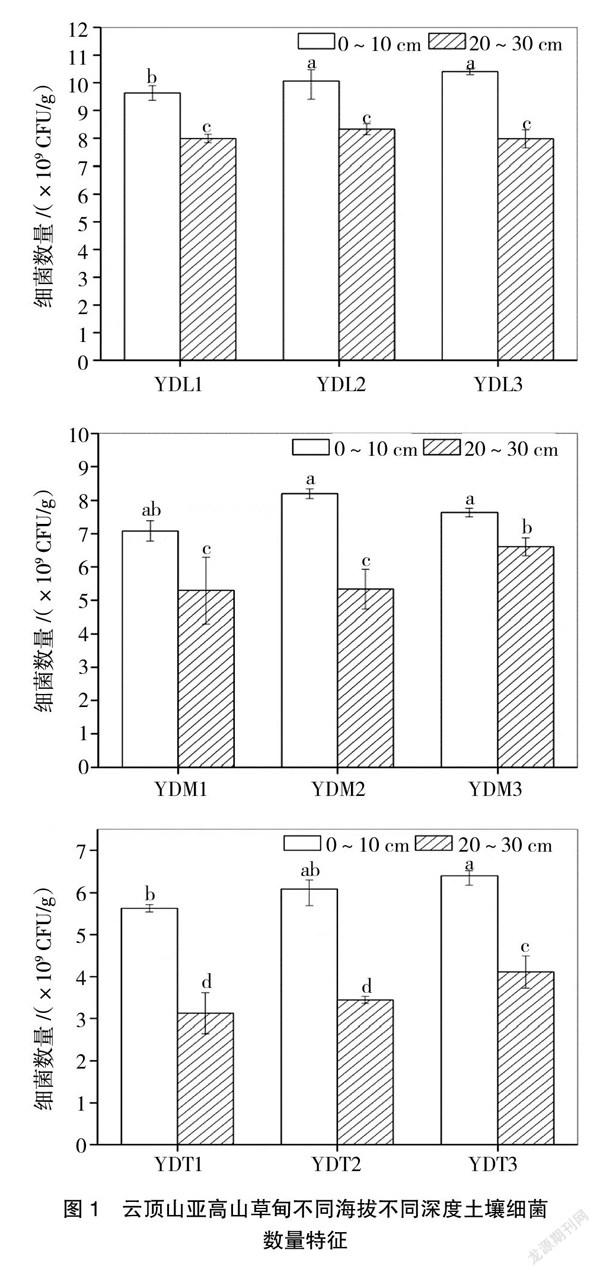
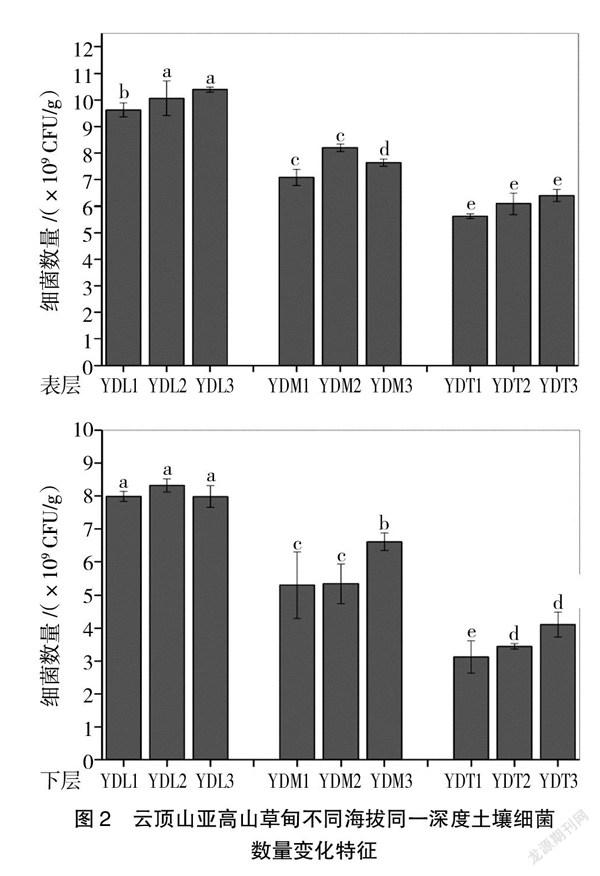
从图1可以看出,研究区域可培养细菌数量介于3.13×109~10.4×109 CFU/g之间,YDL3样点0~10 cm可培养细菌数量最高,为10.4×109 CFU/g,而YDT1样点20~30 cm数量最少,为3.13×109 CFU/g,低了1个数量级。此外,从图1可以看出,同一海拔所有3个重复均表现为0~10 cm细菌数量显著高于20~30 cm细菌数量(P<0.05),说明土壤深度对细菌数量存在显著影响。
纵向来看,从图2可以看出,海拔高度对细菌数量影响显著。不同海拔高度0~10 cm与20~30 cm深度土壤细菌数量均表现为随着海拔高度的升高而显著下降(P<0.05),具体表现为YDL样点土壤细菌数量显著高于YDM与YDT样点(P<0.05),且YDM与YDT样点之间土壤细菌数量也存在显著性差异。
2.3 环境因子与土壤微生物数量的相关分析
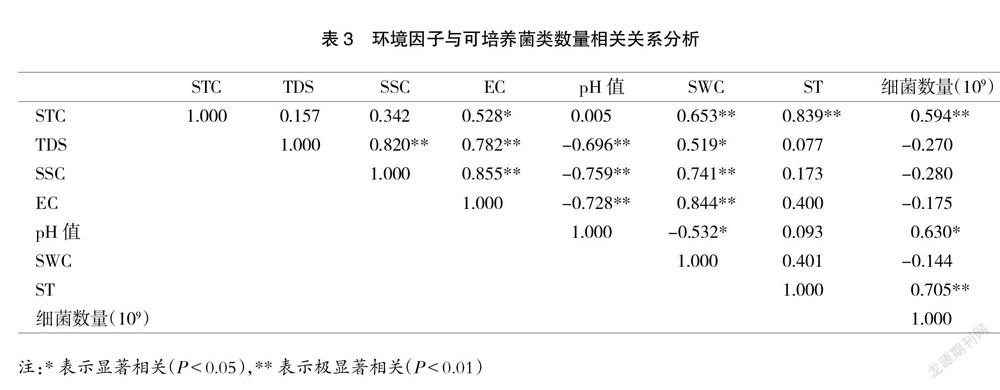
采用SPSS 17.0进行环境因子与可培养细菌数量的相关性进行分析。从表3可知,可培养细菌数量与STC和ST均呈极显著正相关关系(P<0.01),与pH值均呈显著正相关关系(P<0.05)。
3 讨论
细菌是土壤微生物中数量最多的一个类群,占土壤微生物总量的70%~90%,在土壤肥力形成过程中起着重要的作用[23]。研究结果表明,云顶山亚高山草甸土壤细菌数量介于3.13×109 ~10.4×109 CFU/g之间。而相关学者研究发现若尔盖亚高山草甸土壤细菌的数量介于0.5×105~28.5×105 CFU/g之间,这可能是由不同地区环境异质性间接通过冻土温度、植被覆盖度、土壤理化性质等因素对土壤微生物生长繁殖产生影响。
土壤深度和海拔高度也对可培养细菌数量影响明显。本研究表明,同一海拔0~10 cm和20~30 cm深度土壤可培養细菌数量随着土壤深度的增加逐渐变小。这与李定瑶(2014)[24]和李越(2012)[25]等研究者的成果一致,其研究均发现可培养微生物的数量随着土壤深度的增加呈现下降趋势。表层土壤物理环境变化较为剧烈,底物输入波动较大、植物根系与土壤微生物对养分的竞争较为激烈[26],适宜的水分、高含量的有机质和养分状况,使得微生物所受胁迫小,有利于微生物群落的发展[27],而深层土壤水分长期处于饱和状态且土壤温度更低[28],制约了微生物群落的发展,造成不同深度土壤微生物数量变化的因素。
本研究还发现,不同海拔0~10 cm和20~30 cm深度可培养细菌数量均随着海拔的升高呈现下降趋势。这与毛文梁等(2013)[29]研究结果一致,其研究发现不同海拔下祁连山土壤中可培养细菌数量随着海拔的升高而呈现降低的趋势,且两者之间呈现负相关。显然,海拔高度能间接导致温度、水分、光照、土壤微环境及植物群落等环境因子的变化,最终影响到土壤微生物的生长和活性[30]。
相关分析表明,限制研究区内亚高山草甸细菌数量的关键环境因子包括STC、ST与土壤pH值。类似的,相关学者发现北麓河北高寒草原区土壤全碳、含水率与细菌群落结构极显著相关[31];甘南亚高寒草甸不同坡向土壤微生物群落分布的显著差异是受土壤pH值、全磷和含水量等多种环境因子共同作用的结果[32];亚甲贡山土壤温度和总氮是影响细菌生物量的主要因素[33]。也有学者发现,土壤温度是对土壤微生物多样性变化贡献最大的因素[34],这可能是因为细菌适合生存在温度适宜、营养丰富、有机物易于分解的土壤中,而随着海拔不断升高,温度成为环境的主导性因素,因而使得细菌数量呈现随海拔升高而递减的趋势。综上所述,不同区域限制微生物数量的关键环境因子不尽相同,需要区别对待。
4 结论
云顶山亚高山草甸不同海拔、不同土壤深度细菌数量具有明显差异,且与环境因子具有密切关系。本研究结论如下:研究区不同海拔、不同深度可培养细菌数量介于3.13×109~10.4×109 CFU/g之间;不同海拔、不同土壤深度下可培养细菌数量明显不同,具体表现为同一海拔0~10 cm深度土壤细菌数量显著高于20~30 cm深度(P<0.05),不同海拔0~10 cm与20~30 cm深度土壤细菌数量均随着海拔高度的升高而显著下降(P<0.05)。此外,本研究还发现,在环境因素对土壤细菌数量的影响方面,细菌数量受STC和ST极显著影响,受土壤pH值显著影响。
参考文献:
[1]VERESOGLOU S D, HALLEY J M, RILLIG M C. Extinction risk of soil biota[J].Nature Communications,2015(6):8862-8871.
[2]许光辉,郑洪元,张德生,等.长白山北坡自然保护区森林土壤微生物生态分布及其生化特性的研究[J].生态学报,1984,4(3):207-223.
[3]ROY S, SINGH J. Consequences of habitat heterogeneity foravailability of nuterients in a dry tropical forest[J].Journal of Ecology,1994,82(3):503-509.
[4]DILLY O, MUNCH J C. Ratios between estimates of microbialbiomass content and microbial activity in soils[J].Biology and Fertility of Soils,1998,27(4):374-379.
[5]宋长青,吴金水,陆雅海,等.中国土壤微生物学研究10年回顾[J].地球科学进展,2013,28(10):1087-1105.
[6]金凯.中国植被覆盖时空变化及其与气候和人类活动的关系[D].咸阳:西北农林科技大学,2019.
[7]André Pornon, Marion Boutin, Thierry Lamaze. Contribution of plant species to the high N retention capacity of a subalpine meadow undergoing elevated N deposition and warming[J].Environmental Pollution,2018,245(2):235-242.
[8]歐阳青,任健,尹俊,等.短期增温对亚高山草甸土壤养分和脲酶的影响[J].草业科学,2018,35(12):2794-2800.
[9]王萍.亚高山草甸弃耕地恢复演替过程中土壤微生物多样性研究[D].兰州:兰州大学,2011.
[10]宋三多.不同雪被厚度下亚高山草甸土细菌群落结构变化研究[D].雅安:四川农业大学,2011.
[11]谢勇,尹海燕,初晓辉,等.放牧和封育对滇西北亚高山草甸土壤养分及微生物特性的影响[J].云南农业大学学报(自然科学),2017,32(6):1121-1128.
[12]朱雪梅.若尔盖高原亚高山草甸土壤细菌遗传多样性分析[D].雅安:四川农业大学,2009.
[13]李晓丽,徐满厚,孟万忠,等.模拟增温对云顶山亚高山草甸水热因子及群落结构的影响[J].生态学报,2020,40(19):6885-6896.
[14]吴然,康峰峰,韩海荣,等.山西太岳山不同林龄华北落叶松林土壤微生物特性[J].生态学杂志,2016,35(12):3183-3190.
[15]张世雄,杨晓艳,温静,徐满厚.山西吕梁山亚高山草甸物种多样性的时空变化格局[J].生态学报,2018,38(18):685-6693.
[16]马丽,徐满厚,周华坤,等.山西亚高山草甸植被生物量的地理空间分布[J].生态学杂志,2018,37(8):2244-2253.
[17]王誉陶,毕玉婷,王倩,等.山西亚高山草甸植物群落物种多样性的空间分异[J].中国农学通报,2018,34(18):77-83.
[18]上官铁梁,张峰.云顶山虎榛子灌丛群落学特性及生物量[J].山西大学学报(自然科学版),1989,12(3):347-352.
[19]上官铁梁,张峰.云顶山植被及其垂直分布研究[J].山地研究,1991,9(1):26-29.
[20]刘明光,刘莹,张峰,等.云顶山自然保护区植物群落的分类与排序[J].林业资源管理,2011(4):82-88.
[21]李素清,张金屯.山西云顶山亚高山草甸群落生态分析[J].地理研究,2007,26(1):83-90.
[22]李素清,杨斌胜,张金屯.山西云顶山亚高山草甸优势种群和群落的格局分析[J].应用与环境生物学报,2007,13(1):9-13.
[23]Wang K Y, YangW Q, Hu T X. Processes of subalpine forest ecosystem s in the West of Sichuan[M].Chengdu: Sichuan Science& Technologv Press,2004.
[24]李定瑶.青藏高原多年冻土区可培养微生物多样性及生长特性研究[D].兰州:兰州大学,2014.
[25]李越.土壤可培养微生物在黑土碳氮转化中的作用研究[D].长春:吉林农业大学,2012.
[26]Zhu S Y, Zhou B, Mao Z J,et al. Space-time dynamics of fine root biomass of six forests in Maoershan forest region[J].Scientia Silvae Sinicae,2006,42(6):13-19.
[27]李香真,曲秋皓.蒙古高原韵原土境微生物量硕氧特征[J].土壤学报,2002,39(1):97-104.
[28]王长庭,龙瑞军,王启基.不同类型高寒草地群落物种特征和均匀度的重要性[J].草地学报,2005,13(4):320-323.
[29]毛文梁,台喜生,伍修锟,等.黑河上游祁连山区土壤可培养细菌群落生境的垂直分异特征[J].冰川冻土,2013,35(2):447-456.
[30]都耀庭.高寒草甸土壤微生物数量沿海拔梯度的变异[J].黑龙江畜牧兽医,2015(9):130-133.
[31]张晓兰.热融滑塌对青藏高原中部多年冻土区土壤细菌群落的影响[D].兰州:兰州交通大学,2019.
[32]李瑞,刘旻霞,张灿,等.甘南亚高寒草甸不同坡向土壤微生物群落分布特征[J].生态环境学报,2017,26(11):1884-1891.
[33]CUI H J, WANG G X, YANG Y, et al. Soil microbial community composition and its driving factors in alpine grasslands along a mountain elevational gradient[J].Journal of Mountain Science,2016,13(6):1013-1023.
[34]金裕华.武夷山不同海拔土壤微生物多样性的变化特征[D].南京:南京林业大学,2012.