生态恢复模式对若尔盖高寒沙化草地土壤微生物群落功能多样性的影响
2022-02-20朱粟锋刘煜杰张强江聪简小枚税伟
朱粟锋,刘煜杰,张强,江聪,简小枚,税伟
1.中国环境科学研究院
2.北京大学城市与环境学院
3.中国科学院城市环境研究所
4.福州大学环境与资源学院
若尔盖高原地处青藏高原东北边缘,不仅是长江和黄河上游重要的水源涵养地,也是全国重要生态功能区和西北干旱区沙尘暴的有效隔离区以及地球上的天然碳库,对维持地区生物多样性和水源涵养、调节生态系统的碳源-碳汇平衡等起到重要的作用[1-2]。长期的过度放牧、不适当的开垦等人类活动以及气候变化等因素导致湿地退化、草地沙化,这成为该区域面临的两大生态环境问题[3-4]。若尔盖县土地沙化监测结果显示,自20 世纪90 年代以来,该县沙化土地面积持续增加,区域生态环境呈逐步恶化趋势[5-6]。对此,当地政府逐渐启动了系列沙地综合治理的研究与示范点工程,在有效控制沙化土地年递增率的同时,其生态恢复效益也受到广泛关注[7],这一举措为高寒草原地区生态保护与修复提供了典范[8]。当前,对若尔盖草地的恢复治理研究主要集中在植物群落特征、土壤理化性质等方面。如税伟等[9]以空间结构代替时间序列分析了若尔盖地区碳含量、碳储量和碳密度的变化;顾城天等[10]以若尔盖高原湿地植被多样性为基础数据探究了若尔盖高原湿地水质演变特征及氮、磷累积效应;张晓丽等[11]研究了生物质改良剂对高寒草地沙化土壤有机碳特征的影响;刘学敏等[12]则基于不同退化程度的若尔盖草地研究了植物种群的生态位特征。但在深入研究若尔盖高寒植被人工治理程度及其治理机制间的关系方面尚显不足,且对恢复过程中的土壤微生物群落功能多样性研究也相对较少。
土壤微生物作为土壤环境的重要组分,是表征土壤质量变化的敏感指标,也是衡量土壤微生物群落功能多样性的重要指标之一,能够反映群落总体的动态变化,使得地表生态系统和地下生态系统相互关联,在维持土壤生态系统稳定性中起着重要作用[13]。土壤微生物群落功能多样性是衡量土壤微生物群落特征的重要指标,土壤微生物功能多样性的相关研究一直备受关注。若尔盖高原由于其生态功能的特殊地位,正在成为研究的热点区域。经过10 多年的保护修复,若尔盖高寒草地现已基本呈现植物多样性初期的情况,这种情况的出现有利于开展不同恢复重建模式下土壤微生物功能多样性差异及其在重建退化草地生态系统中作用的相关研究。笔者以自然恢复模式与人工草地修复治理模式为研究对象,采用Biolog 法通过设置对比试验对4 种生态恢复模式下高寒草地的土壤微生物进行研究,横向比较不同生态恢复模式下土壤微生物功能多样性的变化差异,纵向比较生态治理措施对沙化草地生态系统的恢复是否有促进作用。通过研究若尔盖草地不同恢复模式下的土壤微生物群落特征,进一步说明人工治理的修复模式对草地生态系统的重要性,以期为高寒草地的退牧还草、退耕还林还草等生态环境重建工作的开展提供参考和依据。
1 研究区域概况
若尔盖高原位于102°08′E~103°39′E,32°56′N~34°19′N,平均海拔为3 500 m,是中国最大的高原,总面积约5.3×106hm2,其中天然草原面积约8.1×105hm2,是我国三大湿地之一,主要草地类型为湿地和草甸,属于高山大陆季风气候区,为高寒温带湿润气候,冬季严寒,夏季凉爽,干雨季节分明,雨热同季,日照充足,昼夜温差大。若尔盖高原行政区域主要包括四川省的若尔盖县、红原县、松潘县和阿坝县,甘肃省的碌曲县、玛曲县以及青海省的久治县(图1)。气象数据显示,1981—2010 年,若尔盖县年平均相对湿度为60%,年平均气温为1.5 ℃,极端最高气温为25.6 ℃,极端最低气温为-30.6 ℃,多年平均降水量为643 mm,年均日照2 417 h,年平均风速为2.3 m/s,最大风速为36 m/s[14]。区域内拥有多种植被类型并存的景观生态格局[15],如未治理区由于过度放牧,植被群落单一,主要是以沙生苔草(Carex praeclara Nelmes)等为主的草本植物;而恢复治理区在采取治理措施下,出现了不同恢复程度的草地治理区,逐渐形成以红柳〔多枝柽柳(Tamarix ramosissima Lcdcb)〕为主的灌木、以豆科(Leguminosae)等牧草和莎草科(Cyperaceae)等杂类草组成的相对复杂的植被状况。
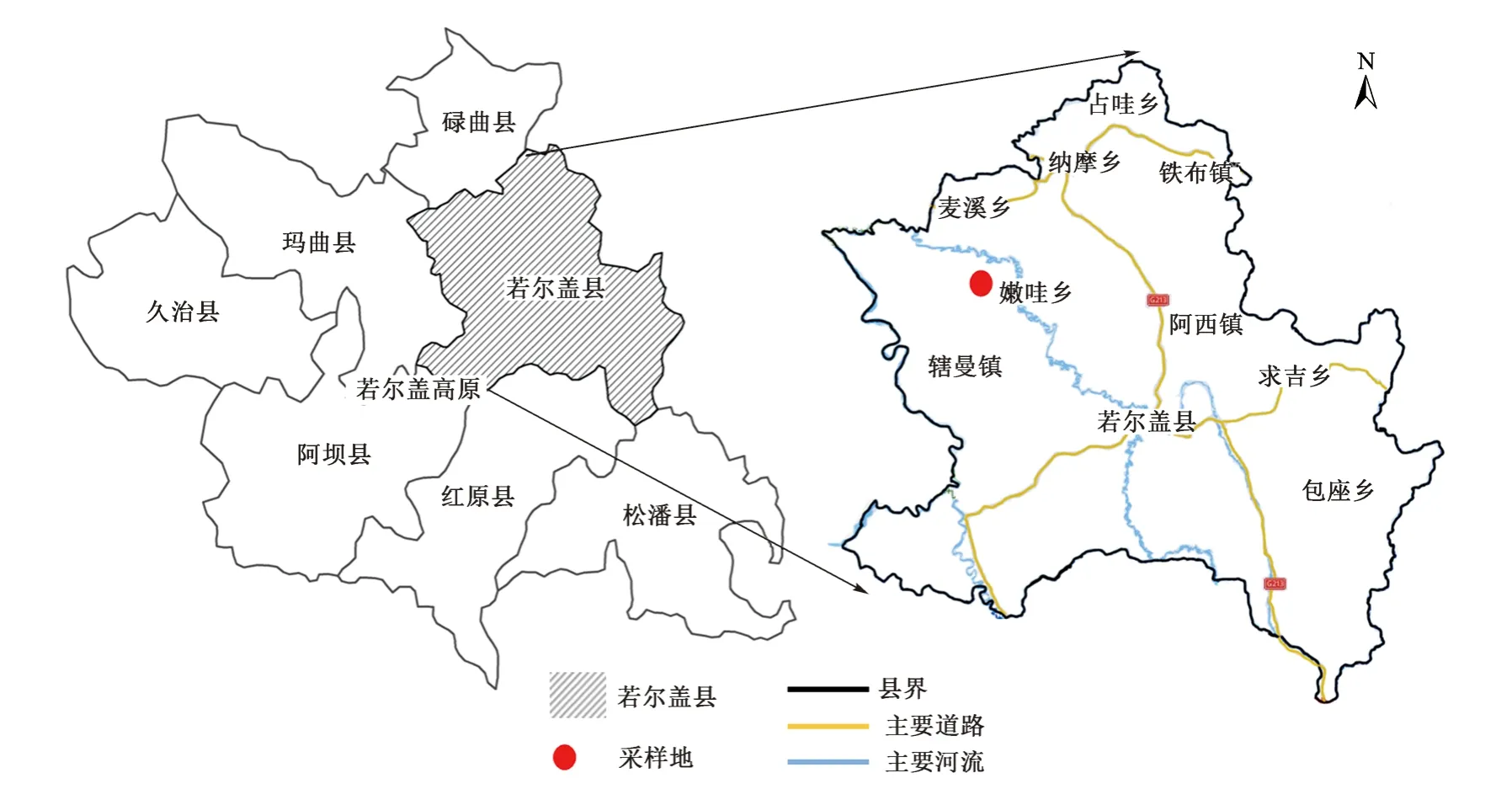
图1 若尔盖县区位Fig.1 Location of Zoige County
2 研究方法
2.1 样地设置
取样地位于若尔盖湿地保护区一片相对平缓的高寒草地内,设置4 种生态恢复模式:条带状高原红柳种植的灌草间作模式和与之相隔5 m 处于全自然状态下的灌草间作模式对照;人工草地建植模式和与之相隔5 m 处于全自然状态下的草地建植模式对照(表1)。
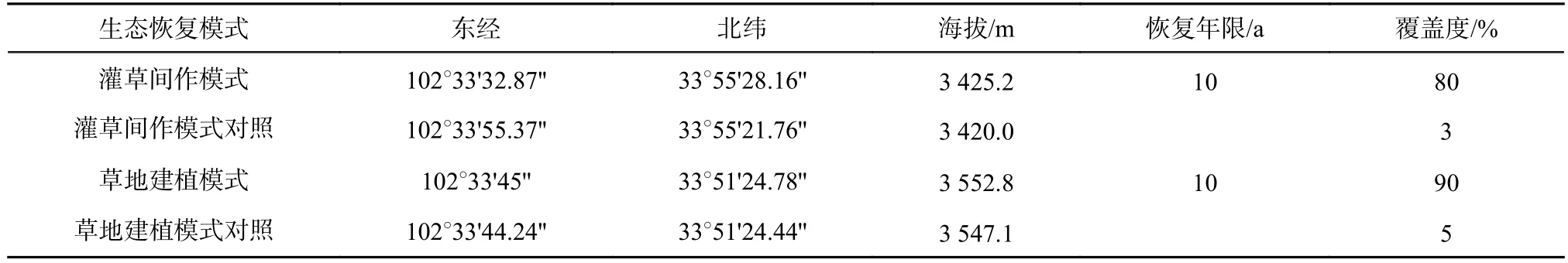
表1 样地基本信息及群落组成Table 1 Basic information and community composition of plots
通过野外考察,在每种典型生态恢复模式样地中随机设置3 个0.5 m×0.5 m 的样方,进行土壤样品采集。在每个样方内,按S 型钻取5 个0~20 cm 土壤样品,均匀混合为1 个土壤样品,放入有冰袋的采集箱中运回实验室,样品过2 mm 筛后,于4 ℃保存,用于土壤微生物功能多样性的测定。样方布设如图2 所示。

图2 不同生态恢复模式的治理样地Fig.2 Test samples of different ecological restoration patterns
2.2 数据处理
采用国际上微生物生态功能多样性研究最典型和最权威方法之一的Biolog 微平板技术分析土壤微生物群落特征,具体操作参照文献[16],计算方法参照文献[17]。
土壤微生物整体活性指标采用微平板每孔颜色平均变化率(average well color development,AWCD)来描述,其可以评判微生物群落对单一碳源利用率的能力。具体计算公式如下:
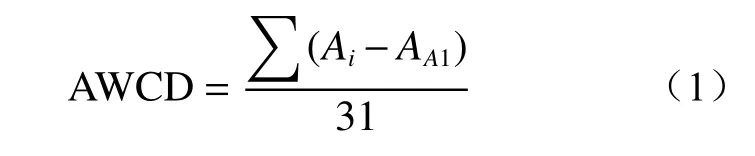
式中:Ai为第i孔的吸光值;AA1为对照孔吸光值。Ai-AA1为负值时归0。
选择AWCD 培养曲线趋于稳定时的数据进行群落多样性指数和主成分分析。其中,采用Shannon指数、Simpson 指数及McIntosh 指数计算和评估土壤微生物群落多样性。计算公式如下:
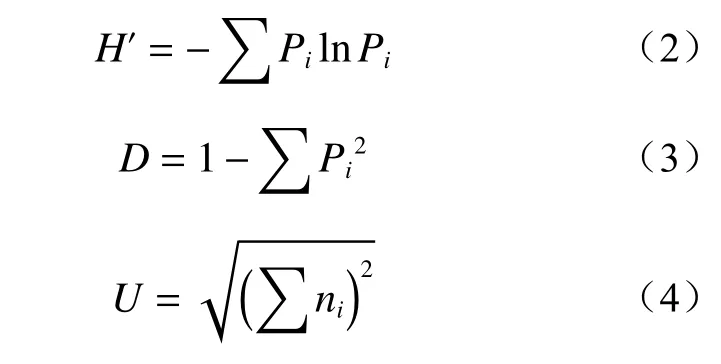
式中:H′为Shannon 指数,用于评估群落丰富度;D为Simpson 指数,用于评估群落常见种优势度;U为McIntosh 指数,用于评估群落均一度;Pi为第i孔的相对吸光值(Ai-AA1)与整个平板相对吸光值总和的比率;ni为第i孔的相对吸光值,即Ai-AA1。
利用SPSS 22.0 及Excel 软件对数据进行处理分析,其中采用单因素方差分析不同生态恢复模式对土壤微生物功能多样性、6 类碳源利用能力的影响差异,采用LSD 最小显著差法和Duncan 多重比较法在0.05 水平上进行显著性差异检验。此外,使用CANOCO for Windows 5.0 软件对培养144 h 的土壤微生物进行主成分分析(principal component analysis,PCA),并采用冗余分析(redundancy analysis,RDA)阐释土壤理化性质与土壤微生物功能多样性的关系。
3 结果与分析
3.1 不同生态恢复模式下草地土壤微生物群落平均颜色变化率
反映土壤微生物在群落水平上利用单一碳源能力的AWCD 变化如图3 所示。由图3 可知,从接种到培养24 h,各土壤样品的AWCD 相对较低,微生物代谢碳源的速率较慢,灌草间作模式变化率在24~48 h 反超草地建植模式,随着培养时间的变化,各恢复模式的AWCD 呈不断上升趋势,微生物进入指数生长期,碳源利用率较高,48 h 后AWCD始终保持灌草间作模式>草地建植模式>灌草间作模式对照>草地建植模式对照。144 h 后各模式AWCD渐趋稳定,但不同样品的AWCD 差异较大,说明不同生态恢复模式对土壤微生物整体活性影响较显著。此外,灌草间作模式下土壤微生物群落对碳源的利用能力显著高于草地建植模式,稳定时二者AWCD 差值为0.563;灌草间作模式下的恢复样地与其对照组的土壤微生物群落的碳源代谢能力相差较大,稳定时二者AWCD 差值为0.761,远大于草地建植模式及其对照组之间的差异。与对照组相比,不同生态恢复模式在恢复过程中均提高了沙化草地的土壤微生物活性,但灌草间作模式的土壤微生物利用碳源的能力远高于草地建植模式。
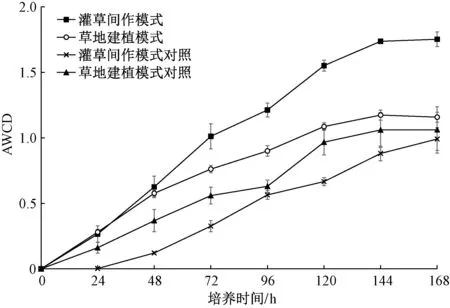
图3 土壤微生物群落平均颜色变化率Fig.3 Average well color development (AWCD) of soil microbial community
3.2 不同生态恢复模式下草地土壤微生物群落功能多样性
选取不同生态恢复模式下平均颜色变化基本趋于稳定状态的144 h 的AWCD 用于土壤微生物群落多样性指数计算,以反映土壤微生物碳源代谢功能多样性差异,结果如表2 所示。由表2 可知,除优势度指数外,2 种生态恢复模式下土壤微生物群落的丰富度指数和均一度指数存在显著性差异,且与相应对照组比较发现,各指数间存在显著性差异。土壤微生物多样性指数差异表明,灌草间作模式及其对照组的草地土壤微生物群落多样性指数相对较高,而草地建植模式及其对照组的多样性指数相对较低。综合比较,灌草间作模式下草地土壤微生物群落的丰富度指数、优势度指数、均一度指数均比草地建植模式及其他对照的土壤微生物群落高,其相应指数分别为3.290、0.960 和10.408,这与AWCD随培养时间的变化曲线表现结果基本一致,表明灌草间作模式下草地的微生物种类最为多样,微生物群落功能多样性最为丰富。
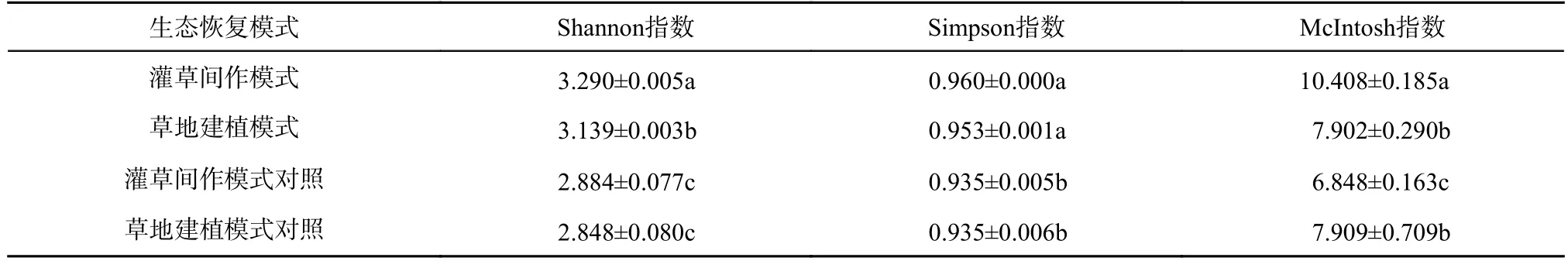
表2 不同生态恢复模式下草地微生物群落功能多样性Table 2 Function diversity index of grassland microbial community of different ecological restoration patterns
3.3 不同生态恢复模式下草地土壤微生物群落碳源利用特征
结合有机化合物化学官能团、微生物代谢途径和生态功能,将Biolog Eco 板的31 种碳源底物分为糖类、氨基酸类、羧酸类、胺类、酚酸类和聚合物类6 类。通过对不同生态恢复模式下草地土壤微生物群落对不同碳源的利用程度分析可知(图4),不同生态恢复模式的草地土壤微生物群落间没有显著差异,氨基酸类是各土壤微生物群落利用最多的碳源,初步反映氨基酸类是若尔盖草地土壤微生物利用的主要碳源。且相较于其他恢复模式和相应对照样地,除对胺类的利用能力不是最高外,灌草间作模式下草地土壤微生物群落的各类碳源能力都是最高的,对糖类、氨基酸类、羧酸类、酚酸类及聚合物类的利用能力值分别达到1.406 2、2.214 6、1.704 4、2.070 3 和2.021 2,草地建植模式对胺类的利用能力最高,达到1.286 5,而对其他几类碳源的利用也相对较高,表明2 种人工修复模式下草地土壤微生物群落有着旺盛的营养代谢活动。因此,若尔盖高寒草地植被在人工种植作用的改善下,对各类碳源的利用能力起到了积极的促进作用,反映出人工治理沙化草地的必要性。
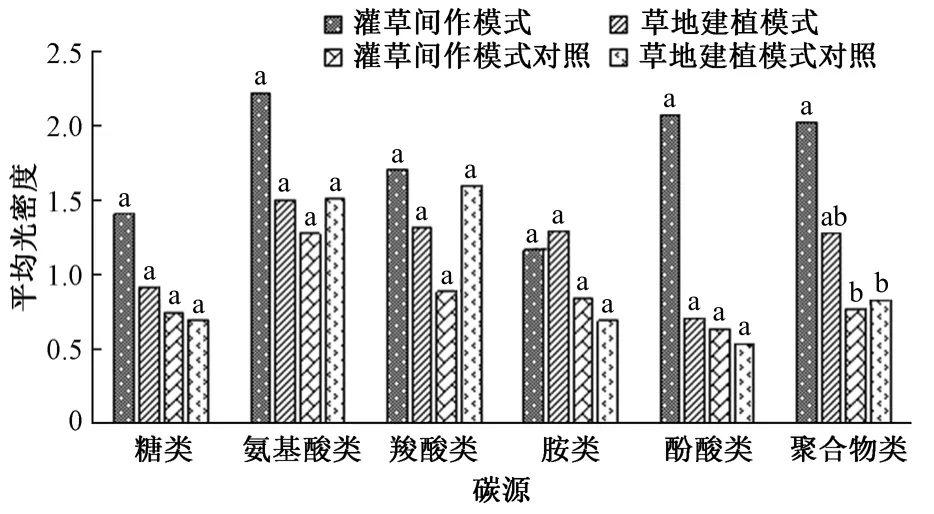
图4 土壤微生物群落对不同碳源的利用能力Fig.4 Utilization capacity of soil microbial community to different carbon sources
3.4 不同生态恢复模式下草地土壤微生物群落碳源利用的主成分分析
微生物群落对碳源的利用情况反映了微生物群落组成的变化,对培养144 h 土壤微生物31 种碳源利用情况进行主成分分析,将31 个因子降维得到4 个主成分(PC),其累积方差贡献率达83.6%,因此认为这4 个主成分可以表示不同生态恢复模式的草地土壤微生物对单一碳源的代谢能力。其中,主成分1 对土壤微生物功能多样性的贡献率为53.3%,主成分2 的贡献率为13.2%,说明前2 个主成分是解释微生物碳源利用变异的主要贡献者(图5)。由图5可知,不同生态恢复模式发生明显分离聚集,表明不同生态恢复模式对土壤微生物群落的碳源利用有较大的影响,且灌草间作模式与其对照组相异明显。通过前2 个主成分的因子载荷发现,对PC1 贡献较大(因子载荷>0.5)的碳源达到20 种,占所有碳源类型的64.5%以上,对PC2 贡献较大(因子载荷>0.5)的碳源则有7 种。此外在主成分分析中,碳水化合物、氨基酸类、多聚物、羧酸类和胺类是灌草间作模式样地上土壤微生物群落主要利用的碳源,草地建植模式样地上土壤微生物群落主要利用的碳源是碳水化合物和胺类。
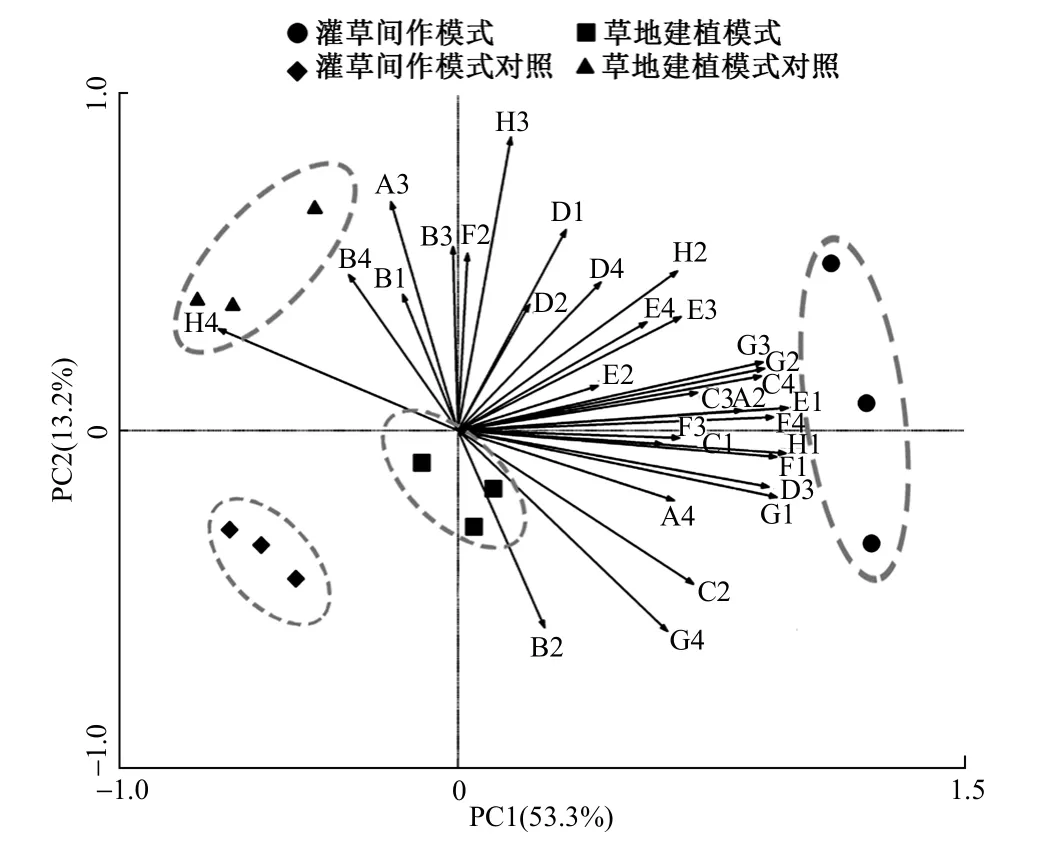
图5 土壤微生物群落主成分分析Fig.5 Principal component analysis of soil microbial community
4 讨论与建议
4.1 人工治理对沙化草地植被恢复的正向促进
人工植被恢复通常是因地制宜地选择适生于当地的植物物种并进行人工培育和种植,以此改善植物在土壤内的根际环境条件,促使沙化草地的土壤功能恢复和改善,能够达到较为长久的防风固沙和植被恢复效果[18]。研究表明,在沙化地采取植被恢复措施,建植适生植被物种,可以有效提高地表植被覆盖度,改善局部气候,降低近地面风速,起到防风固沙的作用,还能够加大地表物质的胶结性,有效改善沙化土地土壤状况,为草原生物的生长繁殖营造良好的生态环境,进而促进植被正向演替,提高区域生物多样性[19-20]。根据“中度干扰理论”[21],适度的人类干扰下,植物根系发育良好且分泌物较为丰富,营造出利于微生物的生存和繁殖的土壤条件,可在一定程度上提高土壤微生物功能多样性[22]。张文娟等[23]也发现砾石方格沙障+播草种的人工干预措施有利于植被的定殖和生长,是一种高寒地区严重沙化土地治理的有效措施。本研究中,人工修复治理本身就是一种中度的干扰形式,灌草间作模式下草地土壤微生物群落的Shannon 指数、Simpson 指数、McIntosh 指数均比草地建植模式及全自然状态下的其他对照样地的土壤微生物群落高,这可能是因为人工培植的高位灌木丛在有效阻挡风沙的同时,也产生了更多的凋落物和根系分泌物来供给土壤微生物,从而提高土壤微生物群落多样性。可见,中度的人工干扰形式有效促进了植被恢复,该结果与前人的研究结果有一定相似性。此外,2 种完全处于自然状态下的对照组之间也表现出差异性,草灌建植对照组McIntosh 指数和AWCD 均更低,这可能是因为草灌建植模式主要采用将灌木类的高原红柳呈条带状种植,并在每株红柳周边自然或人工建植适生于高寒区域的草种植物,红柳和适生草种根系发达,吸收了更多周边草地土壤的营养物质,造成相邻近的草灌建植对照组草地土壤微生物种类分布不均,在长期的生长演替过程中,使草灌建植模式下的人工培植物种在高寒草地生态系统中逐渐处于主导地位,对区域生态植被的改善起到了积极的促进作用。因此,相较于完全处于自然生长状况下的高寒植被,适当的人工修复治理能有效改善高寒草地脆弱的生态环境,应在若尔盖高寒草地的生态修复过程中大面积推广灌草间种的人工治理模式。
4.2 土壤微生物碳源代谢对高寒生境的响应
植被与其生长环境之间是相互影响的,二者处于一种反馈调节的动态平衡之中,在一定程度上共同维持着区域生态系统的稳定[22]。而作为决定土壤微生物功能多样性的主导因素之一,植物群落可通过自身的生长发育来调节土壤中的各种营养物质,进而影响特定生境下土壤微生物功能多样性[24]。地上植被的多样性与土壤微生物群落多样性呈正相关,地表植物多样性能够促进土壤微生物群落代谢及多样性形成[25]。微生物对单一碳源利用程度能够表征土壤微生物群落的代谢活性,微生物也可以通过群落代谢功能的差异来影响环境。本研究中,灌草间作模式草地的AWCD 最高,微生物种类最为多样,微生物群落功能多样性最为丰富,草地土壤微生物群落代谢活性更加活跃,但实地调研发现其植被覆盖度并不是最高的,这可能是因为各植被类型的覆盖度、土壤沙化程度及土壤微生物自身生理特征存在差异。土壤微生物的碳源代谢水平由各立地环境因子共同决定,处于一种动态平衡之中,而土壤理化性状的不同,又直接影响到生长在其上的植物和草地系统生态功能的发挥[26]。若尔盖地区由于高寒的气候环境以及草地的沙化,造成草地土壤的碳、氮、磷、钾等营养元素流失,土壤理化性质以及酶活性改变,降低土壤微生物群落功能多样性,导致植被群落趋向退化状态[27]。对于高寒地区沙化草地而言,采取工程措施和生物措施相结合的方法是其最佳治理方式,而提高酶的代谢活性就是一种重要的生物措施[23]。研究发现,土壤微生物对碳源利用具有选择性,碳水化合物、氨基酸类是当地土壤微生物群落利用的主要碳源,且灌草间作模式的土壤微生物利用碳源的能力远高于其他模式,说明高寒生境下不同人工治理模式的土壤微生物对碳源的利用模式具有差异。这与蒲玉林等[7]通过对比研究若尔盖典型缓丘区不同坡位土壤微生物群落多样性发现氨基酸类是中度坡位上土壤微生物利用的主要碳源的研究结果较为相似。总之,在若尔盖高寒草地大面积推广灌草间种的人工治理模式的同时,应该结合适当增加草地土壤中碳水化合物、氨基酸类等碳源含量的生物措施,以达到高效治理的目的。
5 结语
通过对若尔盖高寒草地不同恢复模式下的土壤微生物群落的研究发现,人工生态恢复模式均能提高沙化草地的土壤微生物的活性,其中灌草间作模式下恢复样地的土壤微生物的活性愈发活跃;土壤微生物群落多样性指数与其群落代谢活性呈正相关,在长期的自然演替过程中灌草间作模式逐渐处于高寒草地生态系统的主导地位;灌草间作恢复模式下土壤微生物群落的各类碳源能力最高,若尔盖草地土壤微生物群落利用最多的碳源是碳水化合物和氨基酸类;采用灌草间作模式的人工治理方式,同时结合适当增加土壤中碳水化合物、氨基酸类碳源的含量的生物措施,可以更高效地对高寒治理生境产生积极响应。
本研究采用的Biolog 微平板法具有一定的局限性,也没有充分考虑物种组成、气候、地形、海拔等外在因子的差异可能对土壤环境带来的影响,今后需进一步采用高通量测序等技术,全面监测地下、地表影响因子数据,多角度对比分析不同生态修复模式下土壤微生物群落组成、结构和多样性等差异,深入探究土壤微生物群落功能多样性,更好地将研究成果服务于若尔盖高寒地区生态植被的保护与修复。
